
Sexually Dimorphic Effects of Neuromodulatory Drugs on Normal and Stress-Induced Social Interaction in Rats


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
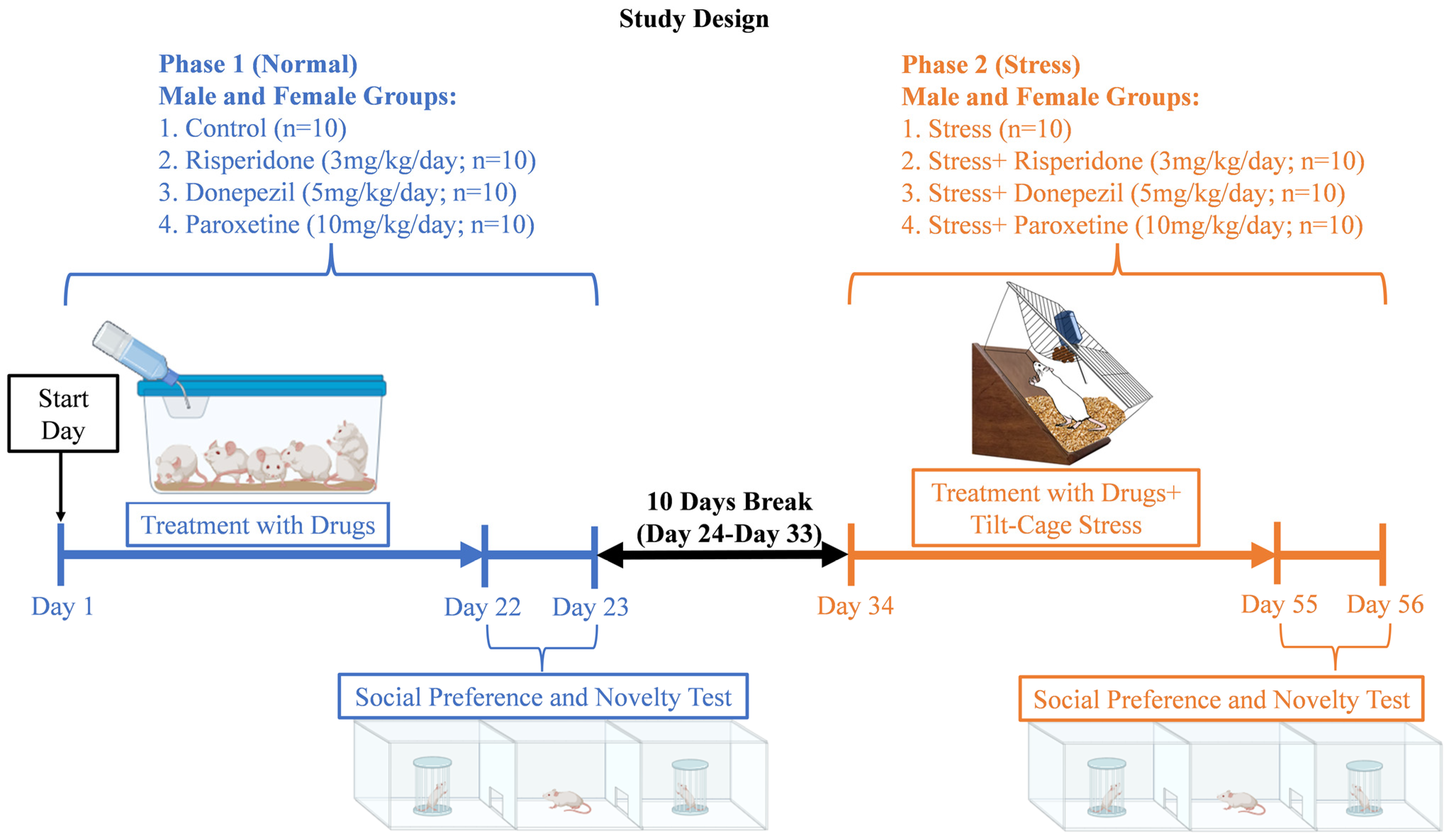
2.1. Animal Groups
2.2. Drugs and Doses
2.3. Scheme of Study
2.4. Social Preference and Novelty Test
2.5. Statistical Analyses
3. Results
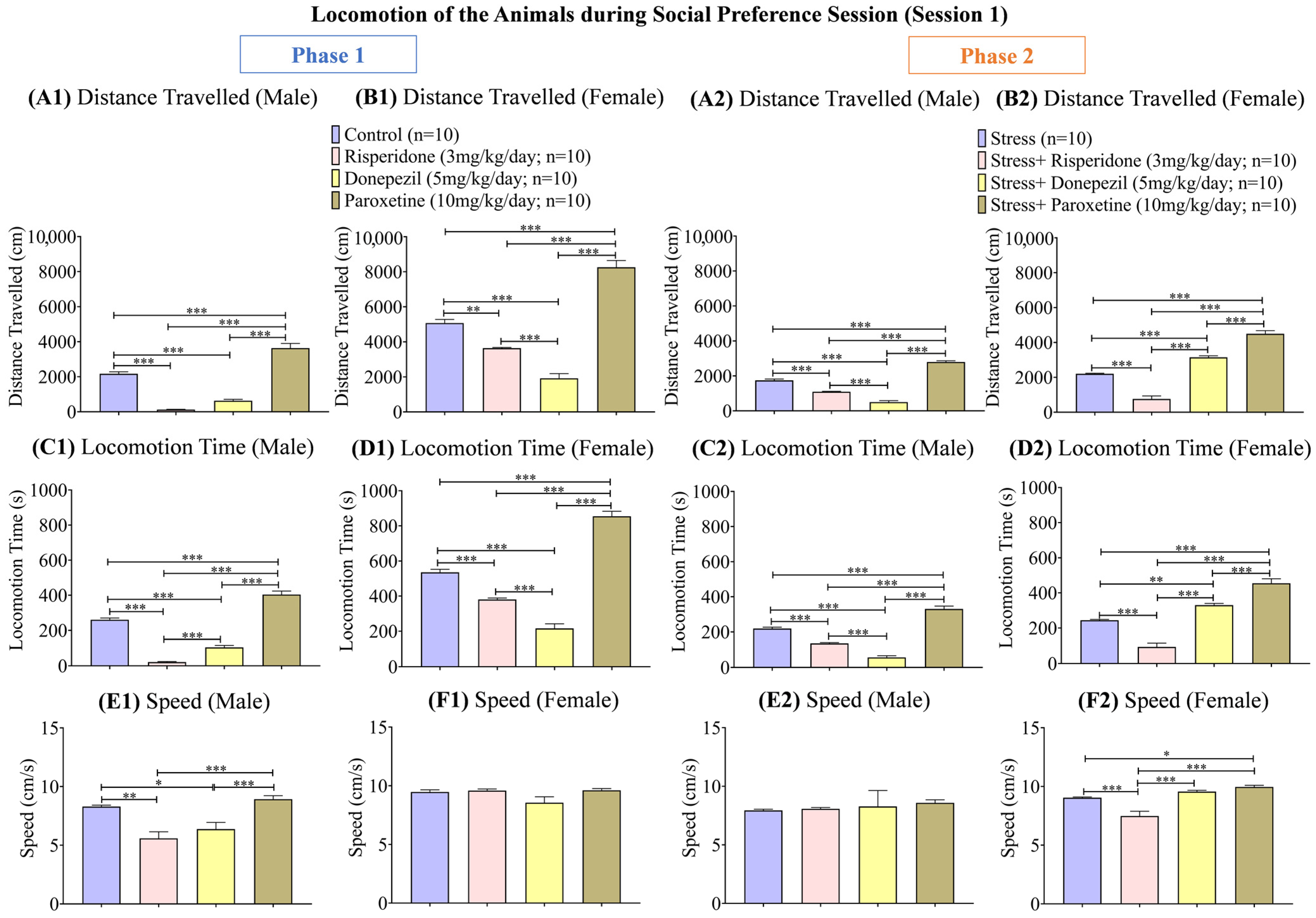
3.1. Sex-Dependent Effects of Neuromodulatory Drugs on Locomotion in Session 1 (Social Preference Session)
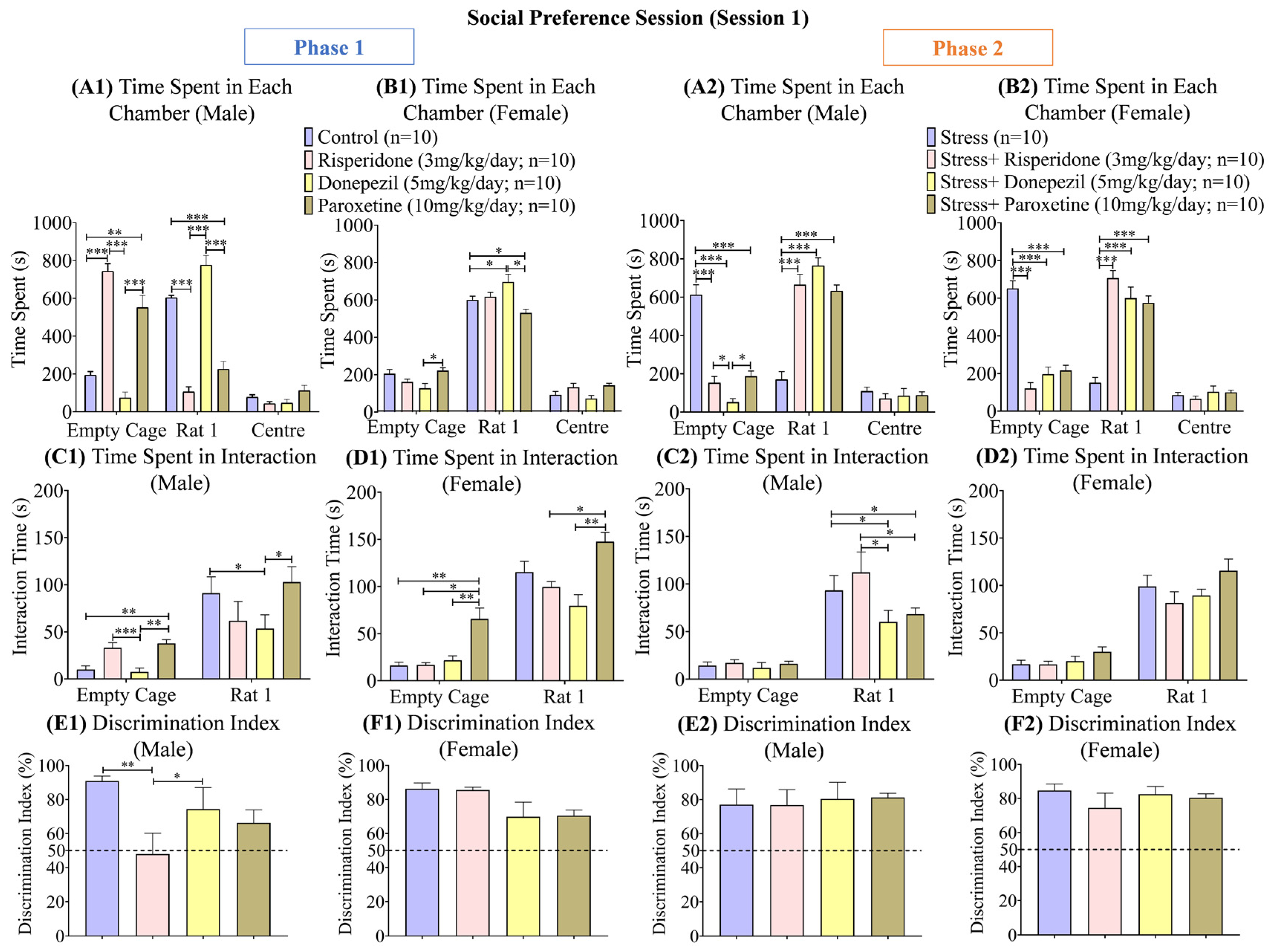
3.2. Sex-Dependent Effects of Neuromodulatory Drugs on Social Preference (Session 1)
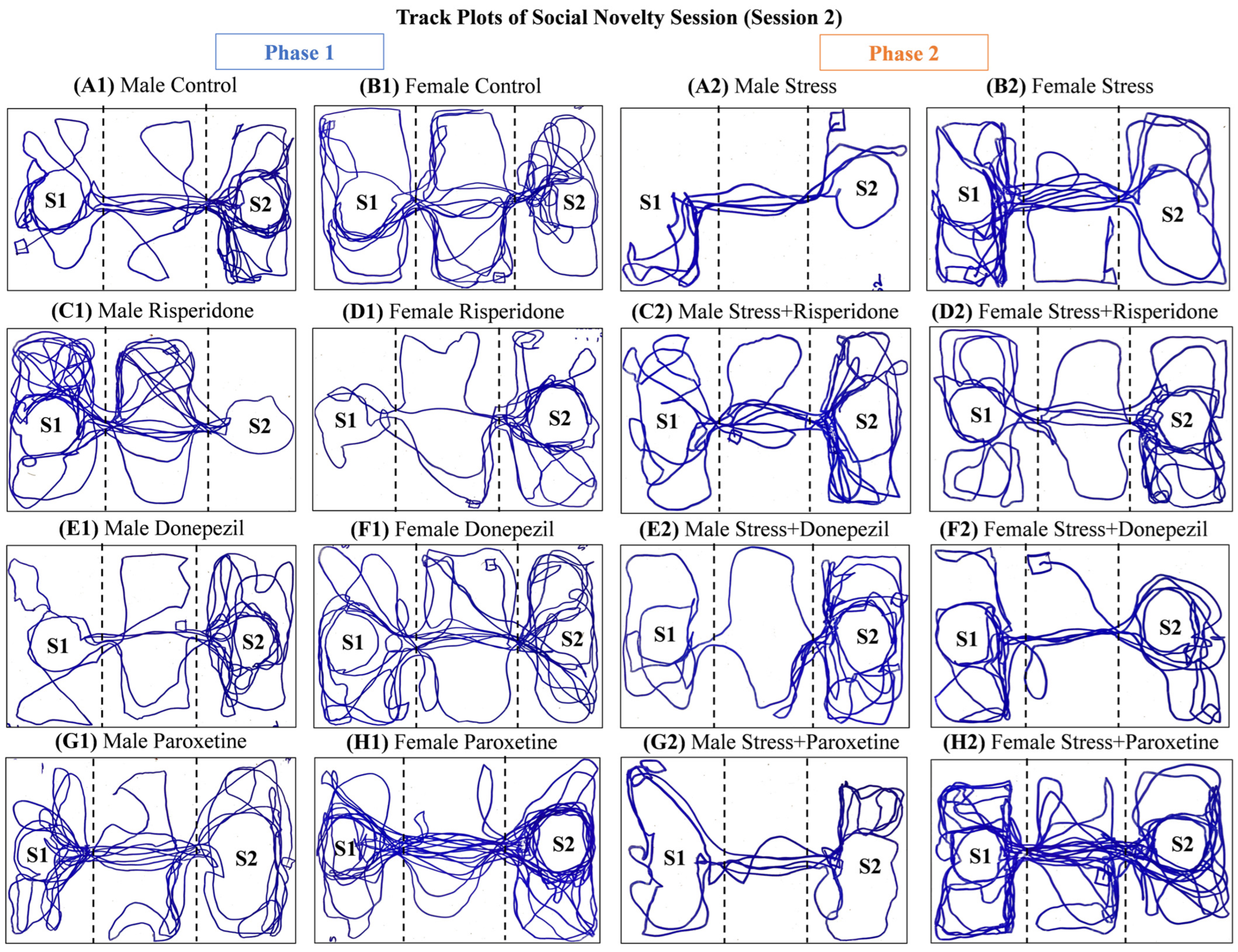
3.3. Sex-Dependent Effects of Neuromodulatory Drugs on Locomotion in Session 2 (Social Novelty Session)
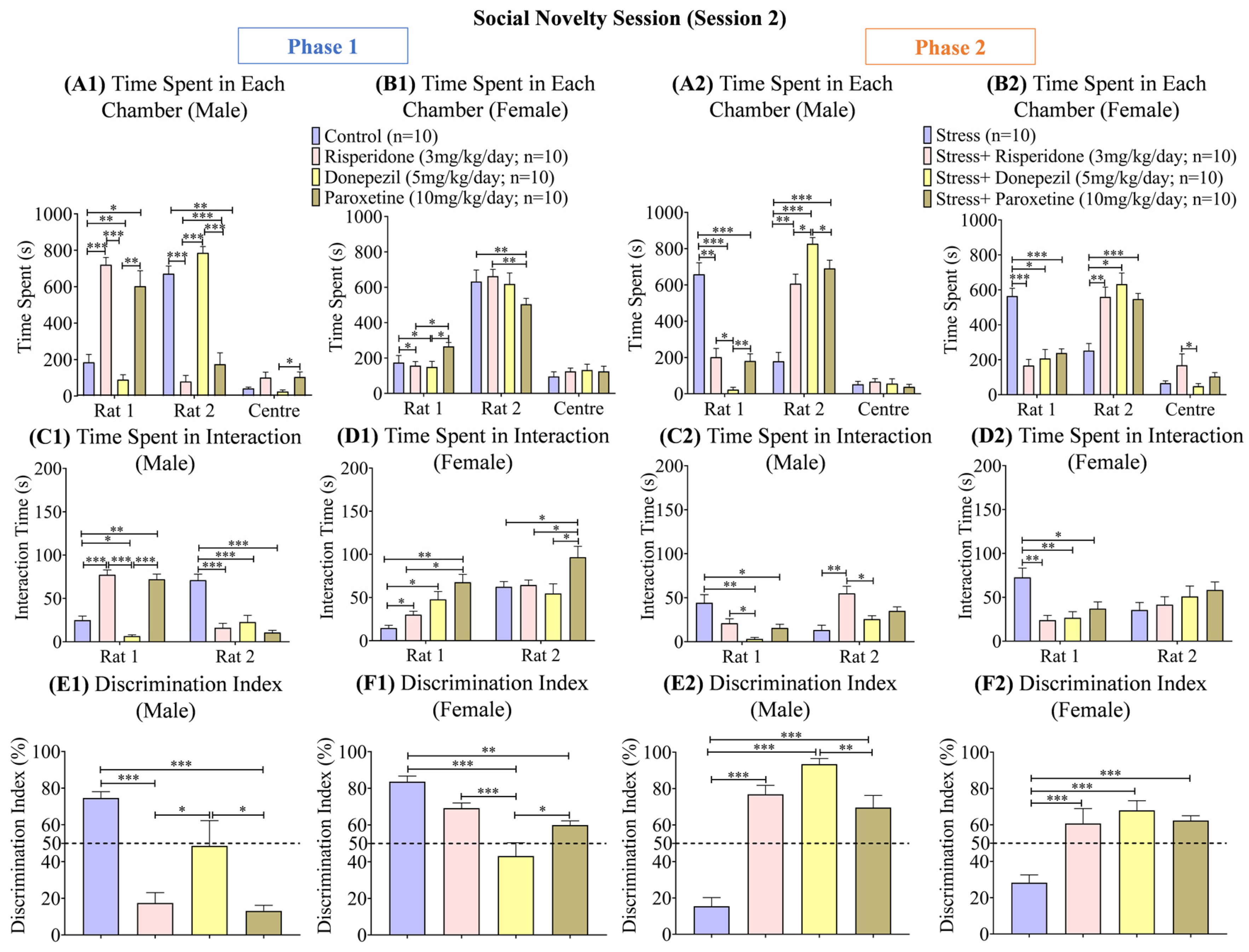
3.4. Sex-Dependent Effects of Neuromodulatory Drugs on Social Novelty (Session 2)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AChE | Acetylcholinesterase |
| ADHD | Attention Deficit Hyperactivity Disorder |
| AD | Alzheimer’s Disease |
| ALS | Amyotrophic Lateral Sclerosis |
| ANOVA | Analysis of Variance |
| ASD | Autism Spectrum Disorder |
| D2 | Dopaminergic Receptor 2 |
| FDA | USA Food and Drug Administration |
| HPA | Hypothalamus-Pituitary-Adrenal |
| HPG | Hypothalamus-Pituitary-Gonadal |
| IRB | Internal Review Board |
| LC | Locus Coeruleus |
| MS | Multiple Sclerosis |
| NAc | Nucleus Accumbens |
| NAOH | Sodium hydroxide |
| NIH | National Institute of Health |
| PD | Parkinson’s Disease |
| S1 | Stranger Animal 1 |
| S2 | Stranger Animal 2 |
| SEM | Standard Error Mean |
| SERT | Serotonin Transporter |
| 5HT1A | 5-Hydroxy tryptamine 1 Receptor A |
| 5HT2A | 5-Hydroxy tryptamine 2 Receptor A |
References
- Chen, P.; Hong, W. Neural circuit mechanisms of social behavior. Neuron 2018, 98, 16–30. [Google Scholar] [CrossRef]
- Alexander, R.D. The evolution of social behavior. Ann. Rev. Ecol. Syst. 1974, 5, 325–383. [Google Scholar] [CrossRef]
- Spencer, S.J.; Buller, K.M.; Day, T.A. Medial prefrontal cortex control of the paraventricular hypothalamic nucleus response to psychological stress: Possible role of the bed nucleus of the stria terminalis. J. Comp Neurol. 2005, 481, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Croxson, P.L.; Kyriazis, D.A.; Baxter, M.G. Cholinergic modulation of a specific memory function of prefrontal cortex. Nat. Neurosci. 2011, 14, 1510–1512. [Google Scholar] [CrossRef]
- Bossert, J.M.; Stern, A.L.; Theberge, F.R.; Marchant, N.J.; Wang, H.-L.; Morales, M.; Shaham, Y. Role of projections from ventral medial prefrontal cortex to nucleus accumbens shell in context-induced reinstatement of heroin seeking. J. Neurosci. 2012, 32, 4982–4991. [Google Scholar] [CrossRef]
- Grossmann, T. The role of medial prefrontal cortex in early social cognition. Front. Human Neurosci. 2013, 7, 340. [Google Scholar] [CrossRef]
- Adolphs, R. The social brain: Neural basis of social knowledge. Ann. Rev. Psych. 2009, 60, 693–716. [Google Scholar] [CrossRef]
- Izquierdo, I.; Bevilaqua, L.R.; Rossato, J.I.; Bonini, J.S.; Medina, J.H.; Cammarota, M. Different molecular cascades in different sites of the brain control memory consolidation. Trends Neurosci. 2006, 29, 496–505. [Google Scholar] [CrossRef]
- Izquierdo, I.; da Cunha, C.; Rosat, R.; Jerusalinsky, D.; Ferreira, M.B.C.; Medina, J.H. Neurotransmitter receptors involved in post-training memory processing by the amygdala, medial septum, and hippocampus of the rat. Behav. Neural Bio. 1992, 58, 16–26. [Google Scholar] [CrossRef]
- McKinsey, G.; Ahmed, O.M.; Shah, N.M. Neural control of sexually dimorphic social behaviors. Curr. Opin. Physiol. 2018, 6, 89–95. [Google Scholar] [CrossRef]
- Luine, V. Sex Differences in Chronic Stress Effects on Memory in Rats. Stress 2002, 5, 205–216. [Google Scholar] [CrossRef]
- Rincón-Cortés, M.; Grace, A.A. Sex-Dependent Effects of Stress on Immobility Behavior and VTA Dopamine Neuron Activity: Modulation by Ketamine. Intern. J. Neuropsychopharmacol. 2017, 20, 823–832. [Google Scholar] [CrossRef]
- Jacobson, L. Glucocorticoid receptor deletion from locus coeruleus norepinephrine neurons promotes depression-like social withdrawal in female but not male mice. Brain Res. 2019, 1710, 82–91. [Google Scholar] [CrossRef]
- Patchev, V.K.; Almeida, O.F. Gonadal steroids exert facilitating and “buffering” effects on glucocorticoid-mediated transcriptional regulation of corticotropin-releasing hormone and corticosteroid receptor genes in rat brain. J. Neurosci. 1996, 16, 7077–7084. [Google Scholar] [CrossRef]
- Duchesne, A.; Dufresne, M.M.; Sullivan, R.M. Sex differences in corticolimbic dopamine and serotonin systems in the rat and the effect of postnatal handling. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, W.; Barker, J.L.; Rubinow, D.R. Sex differences in expression of serotonin receptors (subtypes 1A and 2A) in rat brain: A possible role of testosterone. Neuroscience 1999, 94, 251–259. [Google Scholar] [CrossRef]
- Rashid, H.; Ahmed, T. Gender dimorphic effect of dopamine D2 and muscarinic cholinergic receptors on memory retrieval. Psychopharmacology 2021, 238, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Zagni, E.; Simoni, L.; Colombo, D. Sex and gender differences in central nervous system-related disorders. Neurosci. J. 2016, 2016, 2827090. [Google Scholar] [CrossRef] [PubMed]
- Porcelli, S.; Van Der Wee, N.; van der Werff, S.; Aghajani, M.; Glennon, J.C.; van Heukelum, S.; Mogavero, F.; Lobo, A.; Olivera, F.J.; Lobo, E. Social brain, social dysfunction and social withdrawal. Neurosci. Biobehav. Rev. 2019, 97, 10–33. [Google Scholar]
- Murray, C.J.; Vos, T.; Lozano, R.; Naghavi, M.; Flaxman, A.D.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2197–2223. [Google Scholar] [CrossRef]
- Roehrig, C. Mental disorders top the list of the most costly conditions in the United States: $201 billion. Health Aff. 2016, 35, 1130–1135. [Google Scholar] [CrossRef]
- Jeste, D.V.; Blazer, D.; Casey, D.; Meeks, T.; Salzman, C.; Schneider, L.; Tariot, P.; Yaffe, K. ACNP White Paper: Update on use of antipsychotic drugs in elderly persons with dementia. Neuropsychopharmacology 2008, 33, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.; Lai, T.J.; Hemrungrojn, S.; Mohandas, E.; Yun Kim, S.; Nair, G.; Dash, A. Role of donepezil in the management of neuropsychiatric symptoms in Alzheimer’s disease and dementia with Lewy bodies. CNS Neurosci. Ther. 2016, 22, 159–166. [Google Scholar] [CrossRef]
- Pepeu, G.; Giovannini, M.G. Cholinesterase inhibitors and beyond. Curr. Alzh. Res. 2009, 6, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Cuijpers, P.; Noma, H.; Karyotaki, E.; Vinkers, C.H.; Cipriani, A.; Furukawa, T.A. A network meta-analysis of the effects of psychotherapies, pharmacotherapies and their combination in the treatment of adult depression. World Psych. 2020, 19, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Moran-Gates, T.; Gardner, M.P.; Tarazi, F.I. Effects of repeated risperidone exposure on serotonin receptor subtypes in developing rats. Eur. Neuropsychopharmacol. 2010, 20, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Nevels, R.M.; Gontkovsky, S.T.; Williams, B.E. Paroxetine-The Antidepressant from Hell? Probably Not, But Caution Required. Psychopharmacol. Bull. 2016, 46, 77–104. [Google Scholar]
- Hughes, R.N. Sex still matters: Has the prevalence of male-only studies of drug effects on rodent behaviour changed during the past decade? Behav. Pharmacol. 2019, 30, 95–99. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.M.; Woolley, C.S.; Arnold, A.P. Incorporating sex as a biological variable in neuroscience: What do we gain? Nat. Rev. Neurosci. 2017, 18, 707–708. [Google Scholar] [CrossRef]
- Scharf, S.H.; Sterlemann, V.; Liebl, C.; Müller, M.B.; Schmidt, M.V. Chronic social stress during adolescence: Interplay of paroxetine treatment and ageing. Neuropharmacology 2013, 72, 38–46. [Google Scholar] [CrossRef]
- Sousa, N.; Almeida, O.F. Disconnection and reconnection: The morphological basis of (mal) adaptation to stress. Trends Neurosci. 2012, 35, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Sze, Y.; Brunton, P.J. Sex, stress and steroids. Eur. J. Neurosci. 2020, 52, 2487–2515. [Google Scholar] [CrossRef]
- Rashid, H.; Mahboob, A.; Ahmed, T. Role of cholinergic receptors in memory retrieval depends on gender and age of memory. Behav. Brain Res. 2017, 331, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Rashid, H.; Ahmed, T. Influence of Sex and Muscarinic Activity on Memory Retrieval in Mouse Model of Traumatic Brain Injury. Brain Sci. 2023, 13, 108. [Google Scholar] [CrossRef] [PubMed]
- Rashid, H.; Ahmed, T. Gender dependent contribution of muscarinic receptors in memory retrieval under sub-chronic stress. Neurosci. Lett. 2018, 681, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Kaidanovich-Beilin, O.; Lipina, T.; Vukobradovic, I.; Roder, J.; Woodgett, J.R. Assessment of social interaction behaviors. J. Vis. Exp. 2011, e2473. [Google Scholar] [CrossRef]
- Rizwan, S.; Idrees, A.; Ashraf, M.; Ahmed, T. Memory-enhancing effect of aspirin is mediated through opioid system modulation in an AlCl3-induced neurotoxicity mouse model. Exp. Ther. Med. 2016, 11, 1961–1970. [Google Scholar] [CrossRef]
- Eagly, A.H.; Wood, W. Social role theory. Handb. Theor. Soc. Psychol. 2012, 2, 458–476. [Google Scholar]
- Becker, J.B.; McClellan, M.L.; Reed, B.G. Sex differences, gender and addiction. J. Neurosci. Res. 2017, 95, 136–147. [Google Scholar] [CrossRef]
- Murlanova, K.; Michaelevski, I.; Kreinin, A.; Terrillion, C.; Pletnikov, M.; Pinhasov, A. Link between temperament traits, brain neurochemistry and response to SSRI: Insights from animal model of social behavior. J. Affect. Dis. 2021, 282, 1055–1066. [Google Scholar] [CrossRef]
- Yardimci, A.; Ertugrul, N.U.; Ozgen, A.; Ozbeg, G.; Ozdede, M.R.; Ercan, E.C.; Canpolat, S. Effects of chronic irisin treatment on brain monoamine levels in the hypothalamic and subcortical nuclei of adult male and female rats: An HPLC-ECD study. Neurosci. Lett. 2023, 806, 137245. [Google Scholar] [CrossRef]
- Mei, L.; Zhou, Y.; Sun, Y.; Liu, H.; Zhang, D.; Liu, P.; Shu, H. Acetylcholine Muscarinic Receptors in Ventral Hippocampus Modulate Stress-Induced Anxiety-Like Behaviors in Mice. Front. Mol. Neurosci. 2020, 13, 598811. [Google Scholar] [CrossRef]
- Hudd, T.R.; Blake, C.S.; Rimola-Dejesus, Y.; Nguyen, T.-T.; Zaiken, K. A Case Report of Serotonin Syndrome in a Patient on Selective Serotonin Reuptake Inhibitor (SSRI) Monotherapy. J. Pharm. Prac. 2020, 33, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Morishita, S.; Arita, S. Differential effects of milnacipran, fluvoxamine and paroxetine for depression, especially in gender. Eur. Psych. 2003, 18, 418–420. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, P.J.; Hale, P.J.; Ghimire, A.; Watson, B.O. The cholinesterase inhibitor donepezil has antidepressant-like properties in the mouse forced swim test. Trans. Psychiatry 2020, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, V.; Valentim-Lima, E.; Mencalha, R.; Carbalan, Q.S.R.; Dos-Santos, R.C.; Felintro, V.; Girardi, C.E.N.; Rorato, R.; Lustrino, D.; Reis, L.C.; et al. Neonatal Serotonin Depletion Induces Hyperactivity and Anxiolytic-like Sex-Dependent Effects in Adult Rats. Mol. Neurobiol. 2021, 58, 1036–1051. [Google Scholar] [CrossRef]
- Orendain-Jaime, E.N.; Ortega-Ibarra, J.M.; López-Pérez, S.J. Evidence of sexual dimorphism in D1 and D2 dopaminergic receptors expression in frontal cortex and striatum of young rats. Neurochem. Intern. 2016, 100, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Etaee, F.; Komaki, A.; Faraji, N.; Rezvani-Kamran, A.; Komaki, S.; Hasanein, P.; Taheri, M.; Omidi, G. The effects of cinnamaldehyde on acute or chronic stress-induced anxiety-related behavior and locomotion in male mice. Stress 2019, 22, 358–365. [Google Scholar] [CrossRef]
- Pistovcakova, J.; Makatsori, A.; Sulcova, A.; Jezova, D. Felbamate reduces hormone release and locomotor hypoactivity induced by repeated stress of social defeat in mice. Eur. Neuropsychopharmacol. 2005, 15, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Barth, C.; Villringer, A.; Sacher, J. Sex hormones affect neurotransmitters and shape the adult female brain during hormonal transition periods. Front. Neurosci. 2015, 9, 37. [Google Scholar] [CrossRef]
- Karelia, B.N.; Pillai, A.; Vegada, B.N. The levels of difficulty and discrimination indices and relationship between them in four-response type multiple choice questions of pharmacology summative tests of year II MBBS students. IeJSME 2013, 7, 41–46. [Google Scholar] [CrossRef]
- Wray, N.H.; Rasenick, M.M. Chapter Two—Lipid rafts in psychiatry. In Advances in Pharmacology; Witkin, J.M., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 86, pp. 21–45. [Google Scholar]
- Biederman, J.; Kim, J.W.; Doyle, A.E.; Mick, E.; Fagerness, J.; Smoller, J.W.; Faraone, S.V. Sexually dimorphic effects of four genes (COMT, SLC6A2, MAOA, SLC6A4) in genetic associations of ADHD: A preliminary study. Am. J. Med. Gene. Part B Neuropsychiatr. Genet. 2008, 147b, 1511–1518. [Google Scholar] [CrossRef]
- Morris, L.S.; McCall, J.G.; Charney, D.S.; Murrough, J.W. The role of the locus coeruleus in the generation of pathological anxiety. Brain Neurosci. Adv. 2020, 4, 2398212820930321. [Google Scholar] [CrossRef]
- Borodovitsyna, O.; Joshi, N.; Chandler, D. Persistent Stress-Induced Neuroplastic Changes in the Locus Coeruleus/Norepinephrine System. Neural. Plast. 2018, 2018, 1892570. [Google Scholar] [CrossRef]
- Greenberg, G.D.; Steinman, M.Q.; Doig, I.E.; Hao, R.; Trainor, B.C. Effects of social defeat on dopamine neurons in the ventral tegmental area in male and female California mice. Eur. J. Neurosci. 2015, 42, 3081–3094. [Google Scholar] [CrossRef]
- Kikusui, T.; Winslow, J.T.; Mori, Y. Social buffering: Relief from stress and anxiety. Philos. Transac. Royal Soc. B Biol. Sci. 2006, 361, 2215–2228. [Google Scholar] [CrossRef]
- Benmansour, S.; Cecchi, M.; Morilak, D.A.; Gerhardt, G.A.; Javors, M.A.; Gould, G.G.; Frazer, A. Effects of chronic antidepressant treatments on serotonin transporter function, density, and mRNA level. J. Neurosci. 1999, 19, 10494–10501. [Google Scholar] [CrossRef]
- Silverman, J.L.; Ellegood, J. Behavioral and neuroanatomical approaches in models of neurodevelopmental disorders: Opportunities for translation. Curr. Opin. Neurol. 2018, 31, 126. [Google Scholar] [CrossRef]
- Inagaki, H.; Mori, Y. The emission of stress-induced 22-kHz calls in female rats is independent of testosterone levels. Horm. Behav. 2015, 69, 116–118. [Google Scholar] [CrossRef]
- Council, N.R. Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2010. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishaq, S.; Ahmed, T. Sexually Dimorphic Effects of Neuromodulatory Drugs on Normal and Stress-Induced Social Interaction in Rats. Brain Sci. 2023, 13, 1378. https://doi.org/10.3390/brainsci13101378
Ishaq S, Ahmed T. Sexually Dimorphic Effects of Neuromodulatory Drugs on Normal and Stress-Induced Social Interaction in Rats. Brain Sciences. 2023; 13(10):1378. https://doi.org/10.3390/brainsci13101378
Chicago/Turabian StyleIshaq, Sara, and Touqeer Ahmed. 2023. "Sexually Dimorphic Effects of Neuromodulatory Drugs on Normal and Stress-Induced Social Interaction in Rats" Brain Sciences 13, no. 10: 1378. https://doi.org/10.3390/brainsci13101378
APA StyleIshaq, S., & Ahmed, T. (2023). Sexually Dimorphic Effects of Neuromodulatory Drugs on Normal and Stress-Induced Social Interaction in Rats. Brain Sciences, 13(10), 1378. https://doi.org/10.3390/brainsci13101378