Neurocognitive Endophenotypes for Eating Disorders: A Preliminary High-Risk Family Study

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Procedures
2.3. Measures
2.3.1. Socio Demographic Data
2.3.2. Anthropometry and Pubertal Stage
2.3.3. Child Psychopathology
2.3.4. Global Intelligence
2.3.5. Neurocognitive Function
Attention Switching Task (AST)
Rapid Visual Information Processing (RVP)
Spatial Working Memory (SWM)
2.4. MRI Session
2.5. Data Analysis
2.5.1. Demographic Data and Neurocognitive Function
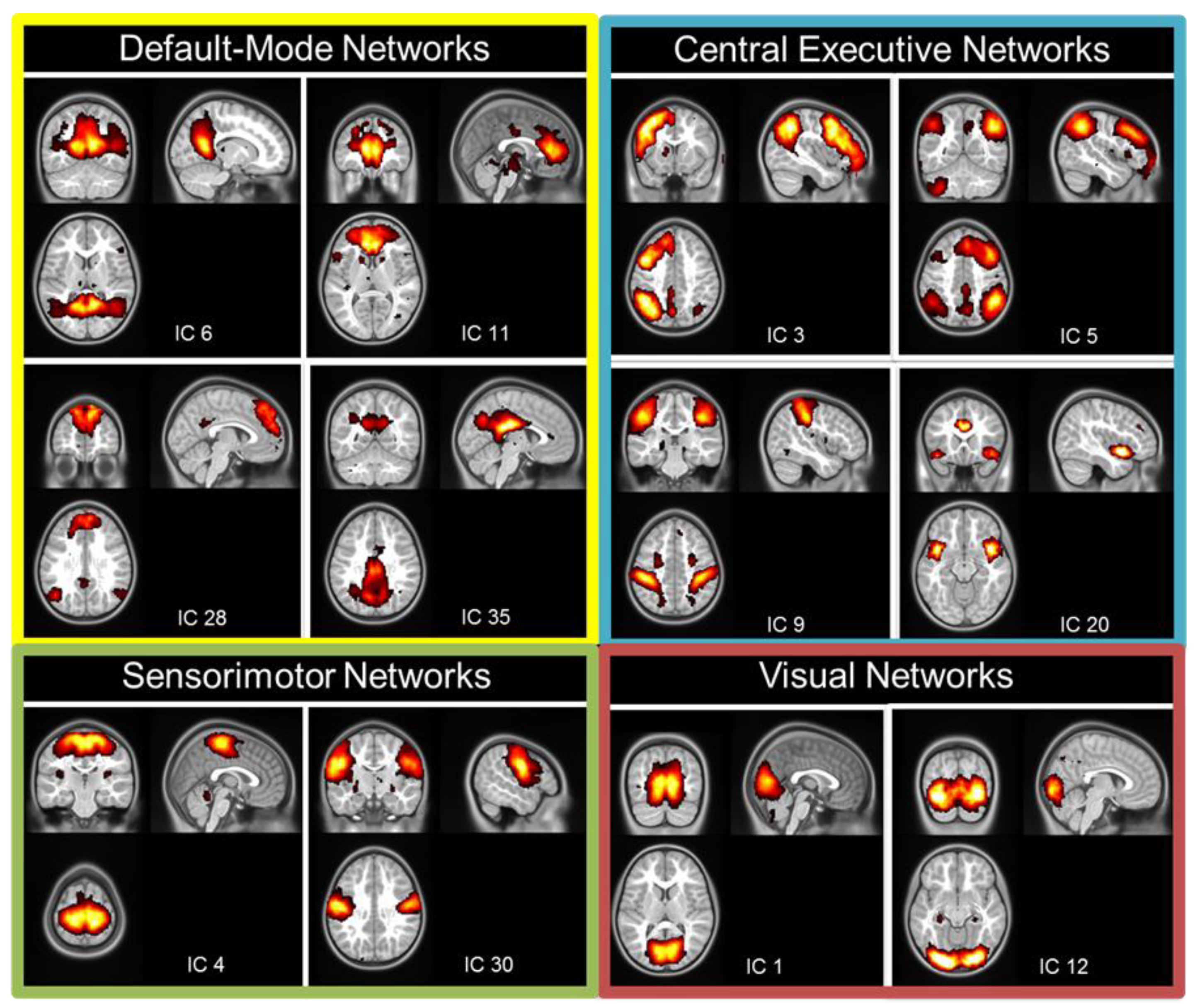
2.5.2. Resting-State fMRI Analysis
3. Results
3.1. Demographic Information
3.2. Neurocognitive Function
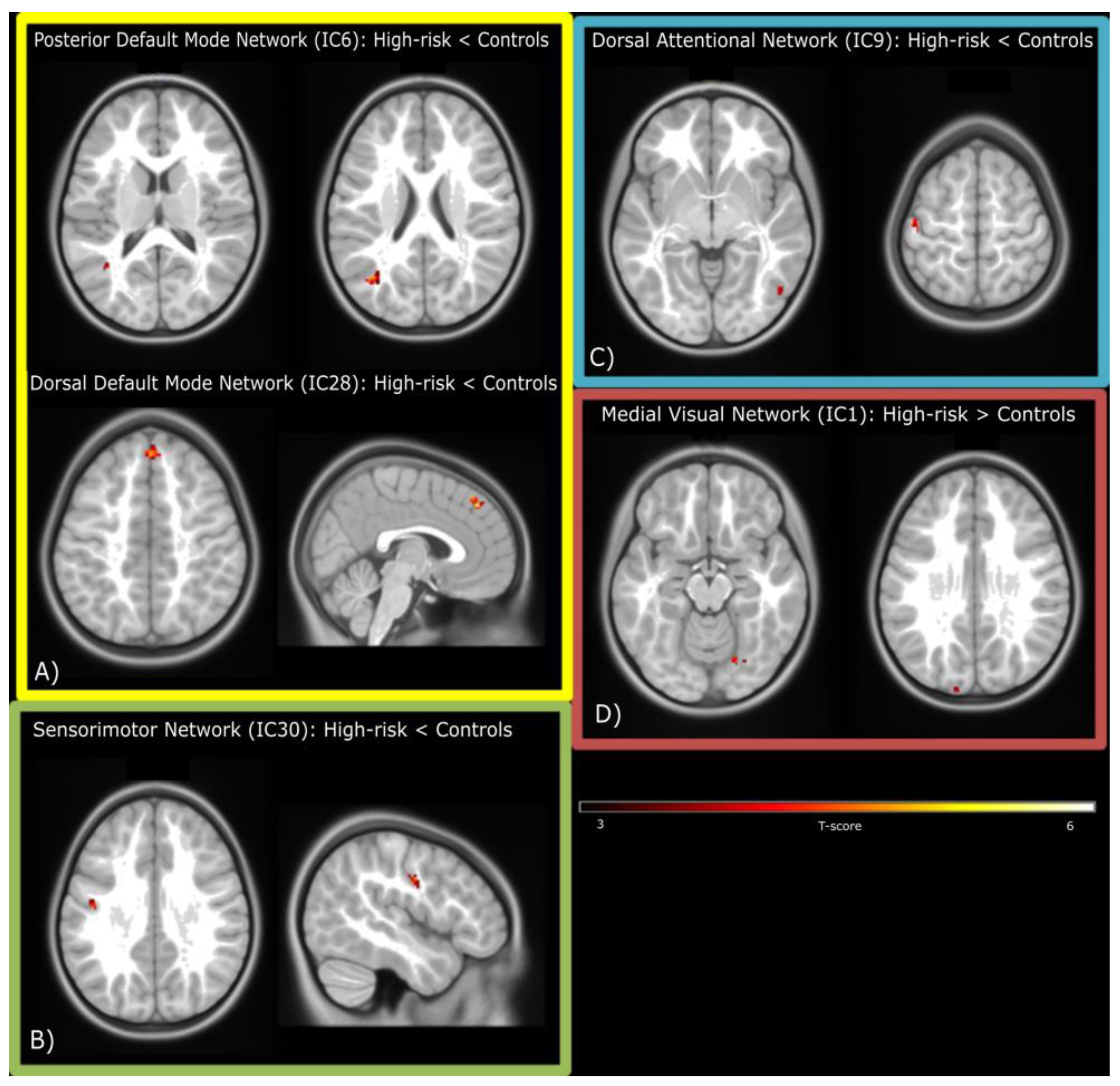
3.3. Resting-State fMRI
4. Discussion
Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Treasure, J.; Duarte, T.A.; Schmidt, U. Eating Disorders. Lancet 2020, 395, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Micali, N.; Solmi, F.; Horton, N.J.; Crosby, R.D.; Eddy, K.T.; Calzo, J.P.; Sonneville, K.R.; Swanson, S.A.; Field, A.E. Adolescent Eating Disorders Predict Psychiatric, High-Risk Behaviors and Weight Outcomes in Young Adulthood. J. Am. Acad. Child Adolesc. Psychiatry 2015, 54, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Micali, N.; Martini, M.G.; Thomas, J.J.; Eddy, K.T.; Kothari, R.; Russell, E.; Bulik, C.M.; Treasure, J. Lifetime and 12-Month Prevalence of Eating Disorders amongst Women in Mid-Life: A Population-Based Study of Diagnoses and Risk Factors. BMC Med. 2017, 15, 12. [Google Scholar] [CrossRef]
- Barona, M.; Brown, M.; Clark, C.; Frangou, S.; White, T.; Micali, N. White Matter Alterations in Anorexia Nervosa: Evidence from a Voxel-Based Meta-Analysis. Neurosci. Biobehav. Rev. 2019, 100, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Kaye, W.H.; Wierenga, C.E.; Bailer, U.F.; Simmons, A.N.; Bischoff-Grethe, A. Nothing Tastes as Good as Skinny Feels: The Neurobiology of Anorexia Nervosa. Trends Neurosci. 2013, 36, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Treasure, J.; Schmidt, U. The Cognitive-Interpersonal Maintenance Model of Anorexia Nervosa Revisited: A Summary of the Evidence for Cognitive, Socio-Emotional and Interpersonal Predisposing and Perpetuating Factors. J. Eat. Disord. 2013, 1, 13. [Google Scholar] [CrossRef] [PubMed]
- Hirst, R.B.; Beard, C.L.; Colby, K.A.; Quittner, Z.; Mills, B.M.; Lavender, J.M. Anorexia Nervosa and Bulimia Nervosa: A Meta-Analysis of Executive Functioning. Neurosci. Biobehav. Rev. 2017, 83, 678–690. [Google Scholar] [CrossRef]
- Lang, K.; Treasure, J.; Tchanturia, K. Is Inefficient Cognitive Processing in Anorexia Nervosa a Familial Trait? A Neuropsychological Pilot Study of Mothers of Offspring with a Diagnosis of Anorexia Nervosa. World J. Biol. Psychiatry 2016, 17, 258–265. [Google Scholar] [CrossRef]
- Naor-Ziv, R.; Glicksohn, J. Investigating Cognitive Deficits as Risk Factors for Developing Eating Disorders During Adolescence. Dev. Neuropsychol. 2016, 41, 107–124. [Google Scholar] [CrossRef]
- Talbot, A.; Hay, P.; Buckett, G.; Touyz, S. Cognitive Deficits as an Endophenotype for Anorexia Nervosa: An Accepted Fact or a Need for Re-Examination? Int. J. Eat. Disord. 2015, 48, 15–25. [Google Scholar] [CrossRef]
- Tchanturia, K.; Harrison, A.; Davies, H.; Roberts, M.; Oldershaw, A.; Nakazato, M.; Stahl, D.; Morris, R.; Schmidt, U.; Treasure, J. Cognitive Flexibility and Clinical Severity in Eating Disorders. PLoS ONE 2011, 6, e20462. [Google Scholar] [CrossRef] [PubMed]
- Tchanturia, K.; Davies, H.; Roberts, M.; Harrison, A.; Nakazato, M.; Schmidt, U.; Treasure, J.; Morris, R. Poor Cognitive Flexibility in Eating Disorders: Examining the Evidence Using the Wisconsin Card Sorting Task. PLoS ONE 2012, 7, e28331. [Google Scholar] [CrossRef] [PubMed]
- Lena, S.M.; Fiocco, A.J.; Leyenaar, J.K. The Role of Cognitive Deficits in the Development of Eating Disorders. Neuropsychol. Rev. 2004, 14, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Greicius, M.D.; Krasnow, B.; Reiss, A.L.; Menon, V. Functional Connectivity in the Resting Brain: A Network Analysis of the Default Mode Hypothesis. Proc. Natl. Acad. Sci. USA 2003, 100, 253–258. [Google Scholar] [CrossRef]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A Default Mode of Brain Function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef]
- Raichle, M.E.; Snyder, A.Z. A Default Mode of Brain Function: A Brief History of an Evolving Idea. NeuroImage 2007, 37, 1083–1090, discussion 1097–1099. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L.; Andrews-Hanna, J.R.; Schacter, D.L. The Brain’s Default Network: Anatomy, Function, and Relevance to Disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Menon, V. Large-Scale Functional Brain Organization. In Brain Mapping; Toga, A.W., Ed.; Academic Press: Waltham, MA, USA, 2015; pp. 449–459. ISBN 978-0-12-397316-0. [Google Scholar]
- Smith, S.M.; Fox, P.T.; Miller, K.L.; Glahn, D.C.; Fox, P.M.; Mackay, C.E.; Filippini, N.; Watkins, K.E.; Toro, R.; Laird, A.R.; et al. Correspondence of the Brain’s Functional Architecture during Activation and Rest. Proc. Natl. Acad. Sci. USA 2009, 106, 13040–13045. [Google Scholar] [CrossRef] [PubMed]
- McFadden, K.L.; Tregellas, J.R.; Shott, M.E.; Frank, G.K.W. Reduced Salience and Default Mode Network Activity in Women with Anorexia Nervosa. J. Psychiatry Neurosci. 2014, 39, 178–188. [Google Scholar] [CrossRef]
- Stopyra, M.A.; Simon, J.J.; Skunde, M.; Walther, S.; Bendszus, M.; Herzog, W.; Friederich, H.-C. Altered Functional Connectivity in Binge Eating Disorder and Bulimia Nervosa: A Resting-State FMRI Study. Brain Behav. 2019, 9, e01207. [Google Scholar] [CrossRef]
- Favaro, A.; Santonastaso, P.; Manara, R.; Bosello, R.; Bommarito, G.; Tenconi, E.; Di Salle, F. Disruption of Visuospatial and Somatosensory Functional Connectivity in Anorexia Nervosa. Biol. Psychiatry 2012, 72, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Phillipou, A.; Abel, L.A.; Castle, D.J.; Hughes, M.E.; Nibbs, R.G.; Gurvich, C.; Rossell, S.L. Resting State Functional Connectivity in Anorexia Nervosa. Psychiatry Res. Neuroimaging 2016, 251, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Scaife, J.C.; Godier, L.R.; Filippini, N.; Harmer, C.J.; Park, R.J. Reduced Resting-State Functional Connectivity in Current and Recovered Restrictive Anorexia Nervosa. Front. Psychiatry 2017, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, B.; Touyz, S.; Hay, P.; Burton, A.; Russell, J.; Caterson, I. Neuroimaging in Bulimia Nervosa and Binge Eating Disorder: A Systematic Review. J. Eat. Disord. 2018, 6, 3. [Google Scholar] [CrossRef]
- Gaudio, S.; Wiemerslage, L.; Brooks, S.J.; Schiöth, H.B. A Systematic Review of Resting-State Functional-MRI Studies in Anorexia Nervosa: Evidence for Functional Connectivity Impairment in Cognitive Control and Visuospatial and Body-Signal Integration. Neurosci. Biobehav. Rev. 2016, 71, 578–589. [Google Scholar] [CrossRef]
- Iacono, W.G. Endophenotypes in Psychiatric Disease: Prospects and Challenges. Genome Med. 2018, 10, 11. [Google Scholar] [CrossRef]
- Bulik, C.M.; Blake, L.; Austin, J. Genetics of Eating Disorders: What the Clinician Needs to Know. Psychiatr. Clin. 2019, 42, 59–73. [Google Scholar] [CrossRef]
- Thornton, L.M.; Mazzeo, S.E.; Bulik, C.M. The Heritability of Eating Disorders: Methods and Current Findings. Curr. Top. Behav. Neurosci. 2011, 6, 141–156. [Google Scholar] [CrossRef]
- Sandstrom, A.; Sahiti, Q.; Pavlova, B.; Uher, R. Offspring of Parents with Schizophrenia, Bipolar Disorder, and Depression: A Review of Familial High-Risk and Molecular Genetics Studies. Psychiatr. Genet. 2019, 29, 160–169. [Google Scholar] [CrossRef]
- Kothari, R.; Solmi, F.; Treasure, J.; Micali, N. The Neuropsychological Profile of Children at High Risk of Developing an Eating Disorder. Psychol. Med. 2013, 43, 1543–1554. [Google Scholar] [CrossRef]
- Kothari, R.; Rosinska, M.; Treasure, J.; Micali, N. The Early Cognitive Development of Children at High Risk of Developing an Eating Disorder. Eur. Eat. Disord. Rev. 2014, 22, 152–156. [Google Scholar] [CrossRef] [PubMed]
- APA. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; APA: Washington, DC, USA, 2013. [Google Scholar]
- First, M.B.; Spitzer, R.L.; Gibbon, M.; Williams, J. Structured Clinical Interview for DSM-IV-TR Axis I Disorders, Research Version; Biometrics Research; New York State Psychiatric Institute: New York, NY, USA, 2002. [Google Scholar]
- Goodman, R. The Strengths and Difficulties Questionnaire: A Research Note. J. Child Psychol. Psychiatry 1997, 38, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R.; Ford, T.; Richards, H.; Gatward, R.; Meltzer, H. The Development and Well-Being Assessment: Description and Initial Validation of an Integrated Assessment of Child and Adolescent Psychopathology. J. Child Psychol. Psychiatry 2000, 41, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Marshall, W.A.; Tanner, J.M. Variations in Pattern of Pubertal Changes in Girls. Arch. Dis. Child. 1969, 44, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Meltzer, H.; Gatward, R.; Goodman, R.; Ford, T. Mental Health of Children and Adolescents in Great Britain. Int. Rev. Psychiatry 2003, 15, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, D. Wechsler Abbreviated Scale of Intelligence—Second Edition; APA PsycNet: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Patel, A.X.; Kundu, P.; Rubinov, M.; Jones, P.S.; Vértes, P.E.; Ersche, K.D.; Suckling, J.; Bullmore, E.T. A Wavelet Method for Modeling and Despiking Motion Artifacts from Resting-State FMRI Time Series. NeuroImage 2014, 95, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Fonov, V.; Evans, A.C.; Botteron, K.; Almli, C.R.; McKinstry, R.C.; Collins, D.L. Unbiased Average Age-Appropriate Atlases for Pediatric Studies. NeuroImage 2011, 54, 313–327. [Google Scholar] [CrossRef]
- Jenkinson, M.; Beckmann, C.F.; Behrens, T.E.J.; Woolrich, M.W.; Smith, S.M. FSL. NeuroImage 2012, 62, 782–790. [Google Scholar] [CrossRef]
- Beckmann, C.F.; Smith, S.M. Probabilistic Independent Component Analysis for Functional Magnetic Resonance Imaging. IEEE Trans. Med. Imaging 2004, 23, 137–152. [Google Scholar] [CrossRef]
- Beckmann, C.; Mackay, C.; Filippini, N.; Smith, S. Group Comparison of Resting-State FMRI Data Using Multi-Subject ICA and Dual Regression. NeuroImage 2009, 47, S148. [Google Scholar] [CrossRef]
- Raichle, M.E. The Brain’s Default Mode Network. Annu. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef]
- Damoiseaux, J.S.; Rombouts, S.A.R.B.; Barkhof, F.; Scheltens, P.; Stam, C.J.; Smith, S.M.; Beckmann, C.F. Consistent Resting-State Networks across Healthy Subjects. Proc. Natl. Acad. Sci. USA 2006, 103, 13848–13853. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Corbetta, M.; Essen, D.C.V.; Raichle, M.E. The Human Brain Is Intrinsically Organized into Dynamic, Anticorrelated Functional Networks. Proc. Natl. Acad. Sci. USA 2005, 102, 9673–9678. [Google Scholar] [CrossRef] [PubMed]
- Dosenbach, N.U.F.; Fair, D.A.; Miezin, F.M.; Cohen, A.L.; Wenger, K.K.; Dosenbach, R.A.T.; Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Raichle, M.E.; et al. Distinct Brain Networks for Adaptive and Stable Task Control in Humans. Proc. Natl. Acad. Sci. USA 2007, 104, 11073–11078. [Google Scholar] [CrossRef]
- Fuglset, T.S. Set-Shifting, Central Coherence and Decision-Making in Individuals Recovered from Anorexia Nervosa: A Systematic Review. J. Eat. Disord. 2019, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.E.; Tchanturia, K.; Stahl, D.; Southgate, L.; Treasure, J. A Systematic Review and Meta-Analysis of Set-Shifting Ability in Eating Disorders. Psychol. Med. 2007, 37, 1075–1084. [Google Scholar] [CrossRef]
- Van den Eynde, F.; Guillaume, S.; Broadbent, H.; Stahl, D.; Campbell, I.C.; Schmidt, U.; Tchanturia, K. Neurocognition in Bulimic Eating Disorders: A Systematic Review. Acta Psychiatr. Scand. 2011, 124, 120–140. [Google Scholar] [CrossRef]
- Holliday, J.; Tchanturia, K.; Landau, S.; Collier, D.; Treasure, J. Is Impaired Set-Shifting an Endophenotype of Anorexia Nervosa? Am. J. Psychiatry 2005, 162, 2269–2275. [Google Scholar] [CrossRef]
- Kanakam, N.; Raoult, C.; Collier, D.; Treasure, J. Set Shifting and Central Coherence as Neurocognitive Endophenotypes in Eating Disorders: A Preliminary Investigation in Twins. World J. Biol. Psychiatry 2013, 14, 464–475. [Google Scholar] [CrossRef]
- Roberts, M.E.; Tchanturia, K.; Treasure, J.L. Exploring the Neurocognitive Signature of Poor Set-Shifting in Anorexia and Bulimia Nervosa. J. Psychiatr. Res. 2010, 44, 964–970. [Google Scholar] [CrossRef]
- Tenconi, E.; Santonastaso, P.; Degortes, D.; Bosello, R.; Titton, F.; Mapelli, D.; Favaro, A. Set-Shifting Abilities, Central Coherence, and Handedness in Anorexia Nervosa Patients, Their Unaffected Siblings and Healthy Controls: Exploring Putative Endophenotypes. World J. Biol. Psychiatry 2010, 11, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Malcolm, A.; Phillipou, A. Current Directions in Biomarkers and Endophenotypes for Anorexia Nervosa: A Scoping Review. J. Psychiatr. Res. 2021, 137, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Weinbach, N.; Bohon, C.; Lock, J. Set-Shifting in Adolescents with Weight-Restored Anorexia Nervosa and Their Unaffected Family Members. J. Psychiatr. Res. 2019, 112, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.J. A Debate on Working Memory and Cognitive Control: Can We Learn about the Treatment of Substance Use Disorders from the Neural Correlates of Anorexia Nervosa? BMC Psychiatry 2016, 16, 10. [Google Scholar] [CrossRef] [PubMed]
- Hatch, A.; Madden, S.; Kohn, M.R.; Clarke, S.; Touyz, S.; Gordon, E.; Williams, L.M. In First Presentation Adolescent Anorexia Nervosa, Do Cognitive Markers of Underweight Status Change with Weight Gain Following a Refeeding Intervention? Int. J. Eat. Disord. 2010, 43, 295–306. [Google Scholar] [CrossRef]
- Seghier, M.L. The Angular Gyrus. Neurosci. 2013, 19, 43–61. [Google Scholar] [CrossRef] [PubMed]
- Vartanian, O.; Goel, V.; Lam, E.; Fisher, M.; Granic, J. Middle Temporal Gyrus Encodes Individual Differences in Perceived Facial Attractiveness. Psychol. Aesthet. Creat. Arts 2013, 7, 38–47. [Google Scholar] [CrossRef]
- Schulte-Rüther, M.; Mainz, V.; Fink, G.R.; Herpertz-Dahlmann, B.; Konrad, K. Theory of Mind and the Brain in Anorexia Nervosa: Relation to Treatment Outcome. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 832–841.e11. [Google Scholar] [CrossRef]
- Goldberg, I.I.; Harel, M.; Malach, R. When the Brain Loses Its Self: Prefrontal Inactivation during Sensorimotor Processing. Neuron 2006, 50, 329–339. [Google Scholar] [CrossRef]
- Mohr, H.M.; Zimmermann, J.; Röder, C.; Lenz, C.; Overbeck, G.; Grabhorn, R. Separating Two Components of Body Image in Anorexia Nervosa Using FMRI. Psychol. Med. 2010, 40, 1519–1529. [Google Scholar] [CrossRef]
- Blatt, G.J. Inhibitory and Excitatory Systems in Autism Spectrum Disorders. In The Neuroscience of Autism Spectrum Disorders; Buxbaum, J.D., Hof, P.R., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 335–346. ISBN 978-0-12-391924-3. [Google Scholar]
- Lavagnino, L.; Amianto, F.; D’Agata, F.; Huang, Z.; Mortara, P.; Abbate-Daga, G.; Marzola, E.; Spalatro, A.; Fassino, S.; Northoff, G. Reduced Resting-State Functional Connectivity of the Somatosensory Cortex Predicts Psychopathological Symptoms in Women with Bulimia Nervosa. Front. Behav. Neurosci. 2014, 8, 270. [Google Scholar] [CrossRef] [PubMed]
- Bartholdy, S.; Rennalls, S.J.; Jacques, C.; Danby, H.; Campbell, I.C.; Schmidt, U.; O’Daly, O.G. Proactive and Reactive Inhibitory Control in Eating Disorders. Psychiatry Res. 2017, 255, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Yano, M.; Kawano, N.; Tanaka, S.; Kohmura, K.; Katayama, H.; Nishioka, K.; Ozaki, N. Dysfunction of Response Inhibition in Eating Disorders. J. Clin. Exp. Neuropsychol. 2016, 38, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Tang, Y.; Kang, Q.; Wang, Q.; Wang, J.; Chen, J. Deficits in Response Inhibition on Varied Levels of Demand Load in Anorexia Nervosa: An Event-Related Potentials Study. Eat. Weight Disord.-Stud. Anorex. Bulim. Obes. 2020, 25, 231–240. [Google Scholar] [CrossRef]
- Fox, M.D.; Corbetta, M.; Snyder, A.Z.; Vincent, J.L.; Raichle, M.E. Spontaneous Neuronal Activity Distinguishes Human Dorsal and Ventral Attention Systems. Proc. Natl. Acad. Sci. USA 2006, 103, 10046–10051. [Google Scholar] [CrossRef] [PubMed]
- Cooper, Z.; Dalle Grave, R. Chapter 14—Eating Disorders: Transdiagnostic Theory and Treatment. In The Science of Cognitive Behavioral Therapy; Hofmann, S.G., Asmundson, G.J.G., Eds.; Academic Press: San Diego, CA, USA, 2017; pp. 337–357. ISBN 978-0-12-803457-6. [Google Scholar]
- Fairburn, C.G.; Cooper, Z.; Shafran, R. Cognitive Behaviour Therapy for Eating Disorders: A “Transdiagnostic” Theory and Treatment. Behav. Res. Ther. 2003, 41, 509–528. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| FHR for ED (N = 16) | HC (N = 20) | Group Comparison 1 | |
|---|---|---|---|
| Means (SD) | Means (SD) | ||
| Age (years) | 12 (2.19) | 12.25 (1.94) | t = −0.36, p = 0.72 |
| Breast Developmental Stage | 3 (1.55) | 3.05 (1.43) | t = −0.10, p = 0.92 |
| Pubic Hair Developmental Stage | 3.25 (1.77) | 3.16 (1.46) | t = 0.17, p = 0.87 |
| WASI-Full-scale IQ (FIQ) | 120.94 (12.04) | 120.55 (14.45) | t = 0.08, p = 0.93 |
| SDQ—Total difficulties | 11 (4.87) | 8.89 (5.85) | t = 1.14, p = 0.26 |
| SQD—Emotional difficulties | 3.8 (2.30) | 3 (2.49) | t = 0.97, p = 0.34 |
| SDQ—Conduct difficulties | 1.60 (1.24) | 1.21 (1.40) | t = 0.86, p = 0.40 |
| SDQ—Hyperactivity difficulties | 3.5 (2.59) | 2.68 (2) | t = 1.13, p = 0.27 |
| SDQ—Peer difficulties | 2.47 (2.26) | 2 (1.76) | t = 0.66, p = 0.52 |
| SDQ—Prosocial difficulties | 7.93 (2.60) | 8.32 (1.86) | t = −0.50, p = 0.62 |
| Maternal age (years) | 43.81 (5.6) | 45.73 (5.41) | t = −1.03, p = 0.31 |
| Frequencies | Frequencies | ||
| Maternal Lifetime Diagnosis | AN (14) BN (2) | ||
| Maternal Education | Higher education (13) Up to A level (3) | Higher education (18) Up to A level (2) | X2(1) = 0.57 |
| Maternal Ethnicity | White/Caucasian (15) Other (1) | White/Caucasian (17) Other (3) | X2(1) = 0.69 |
| Outcome | Controls | High-Risk ED | Crude Group Comparison | Adjusted Group Comparison + |
|---|---|---|---|---|
| AST—Correct | 92.56 (6.48) | 94.65 (4.96) | F(1,34) = 1.13, p = 0.30, η2 = 0.03 | F(1,32) = 1.54, p = 0.22, η2= 0.04 |
| AST—Congruency cost | 43.86 (59.75) | 71.25 (38.49) | F(1,34) = 2.52, p = 0.12, η2 = 0.07 | F(1,32) = 2.29, p = 0.14, η2 = 0.14 |
| AST—Switching cost | 159.95 (94.22) | 240.86 (111.80) | F(1,34) = 5.55, p = 0.02, η2 = 0.14 | F(1,32) = 5.53, p = 0.02, η2 = 0.12 |
| AST—Mean latency | 682.67 (143.88) | 774.52 (135.37) | F(1,34) = 3.81, p = 0.06, η2 =0.10 | F(1,32) = 3.80, p = 0.06, η2 =0.09 |
| RVP—Mean latency | 364.94 (90.76) | 376.57 (88.12) | F(1,32) = 0.06, p = 0.81, η2 =0.001 | F(1,32) = 0.15, p = 0.70, η2 = 0.004 |
| SWM—Errors | 28.20 (23.20) | 26.94 (21.67) | F(1,34) = 0.03, p = 0.87, η2 =0.001 | F(1,32) = 0.19, p = 0.67, η2 =0.003 |
| SWM—Strategy | 28.65 (9.01) | 33.44 (6.69) | F(1,34) = 3.13, p = 0.09, η2 =0.08 | F(1,32) = 2.96, p = 0.09, η2 = 0.08 |
| Regions | Hemisphere | T | n Voxels | p-Value (FDR Corrected) | x | y | z |
|---|---|---|---|---|---|---|---|
| Posterior DMN (IC6): Controls > High-risk ED children | |||||||
| Angular Gyrus | Left | 4.99 | 22 | 0.048 | −38 | −66 | 22 |
| Middle Temporal Gyrus | Left | 4.61 | 31 | 0.016 | −40 | −60 | 12 |
| Dorsal DMN (IC28): Controls > High-risk ED children | |||||||
| Medial Superior Frontal Gyrus | Right | 5.8 | 52 | <0.001 | 2 | 42 | 44 |
| Sensorimotor Network (IC30): Controls > High-risk ED children | |||||||
| Postcentral Gyrus | Left | 4.7 | 25 | 0.012 | −46 | −10 | 34 |
| Attentional Network (IC9): Controls > High-risk ED children | |||||||
| Postcentral Gyrus | Left | 5.78 | 32 | 0.03 | −48 | −18 | 62 |
| Inferior Temporal Gyrus | Right | 5.22 | 24 | 0.03 | 50 | −64 | −2 |
| Visual Network (IC1): High-risk ED children > Controls | |||||||
| Fusiform Gyrus | Right | 4.79 | 21 | 0.028 | 22 | −64 | −12 |
| Superior Occipital Gyrus | Left | 4.6 | 29 | 0.024 | −14 | −100 | 26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pappaianni, E.; Barona, M.; Doucet, G.E.; Clark, C.; Frangou, S.; Micali, N. Neurocognitive Endophenotypes for Eating Disorders: A Preliminary High-Risk Family Study. Brain Sci. 2023, 13, 99. https://doi.org/10.3390/brainsci13010099
Pappaianni E, Barona M, Doucet GE, Clark C, Frangou S, Micali N. Neurocognitive Endophenotypes for Eating Disorders: A Preliminary High-Risk Family Study. Brain Sciences. 2023; 13(1):99. https://doi.org/10.3390/brainsci13010099
Chicago/Turabian StylePappaianni, Edoardo, Manuela Barona, Gaelle E. Doucet, Christopher Clark, Sophia Frangou, and Nadia Micali. 2023. "Neurocognitive Endophenotypes for Eating Disorders: A Preliminary High-Risk Family Study" Brain Sciences 13, no. 1: 99. https://doi.org/10.3390/brainsci13010099
APA StylePappaianni, E., Barona, M., Doucet, G. E., Clark, C., Frangou, S., & Micali, N. (2023). Neurocognitive Endophenotypes for Eating Disorders: A Preliminary High-Risk Family Study. Brain Sciences, 13(1), 99. https://doi.org/10.3390/brainsci13010099