
Optimizing the Effect of tDCS on Motor Sequence Learning in the Elderly

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Transcranial Direct Current Stimulation (tDCS)
2.3. Serial Reaction Time Task (SRTT) as a Measure of Implicit Motor Sequence Learning
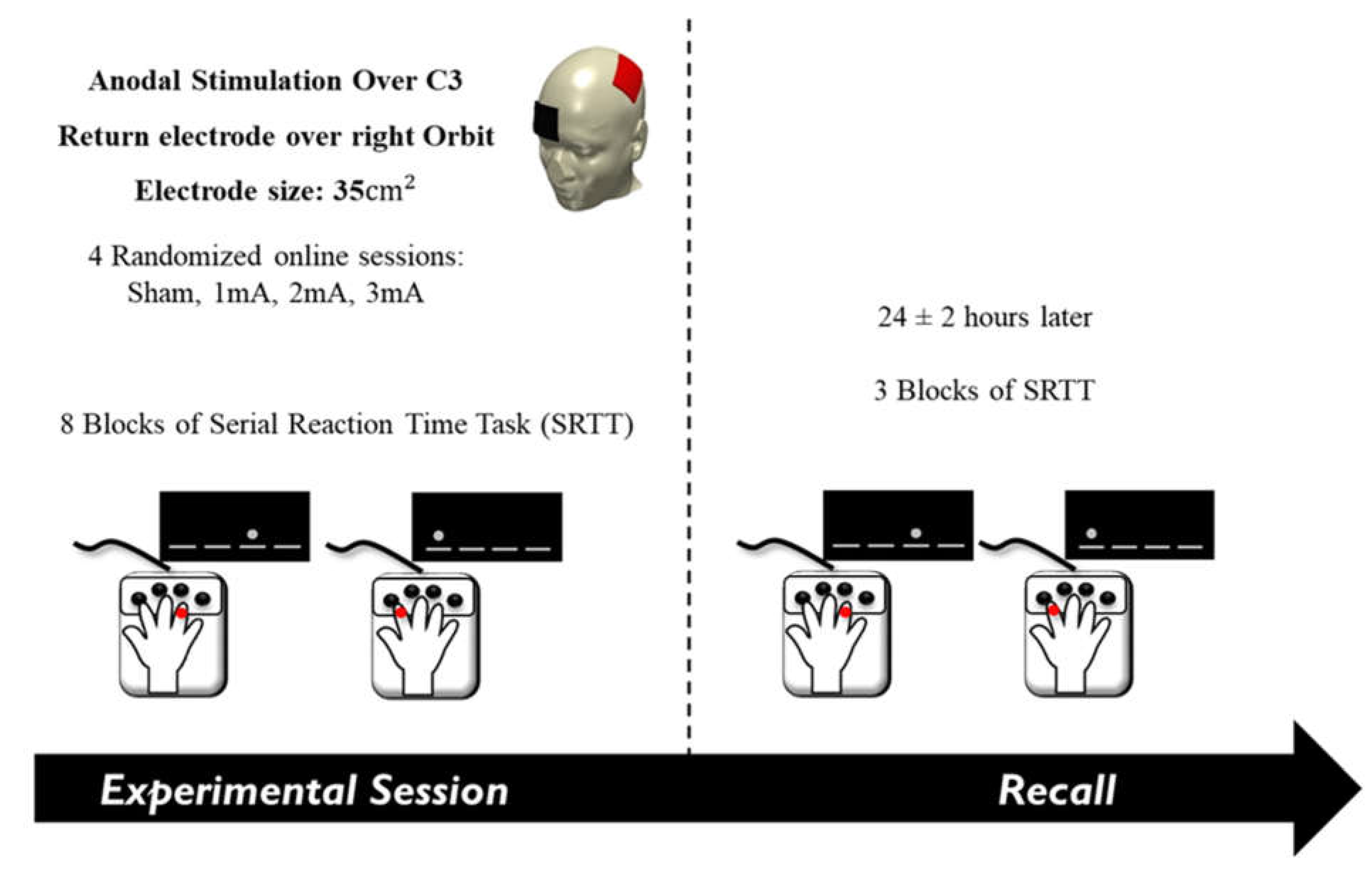
2.4. Experimental Procedure
2.5. Data Analysis and Statistics
2.5.1. Effect of tDCS on M-SRTT
2.5.2. Effect of tDCS on R-SRTT
2.5.3. Assessment of tDCS Side Effects and Blinding
3. Results
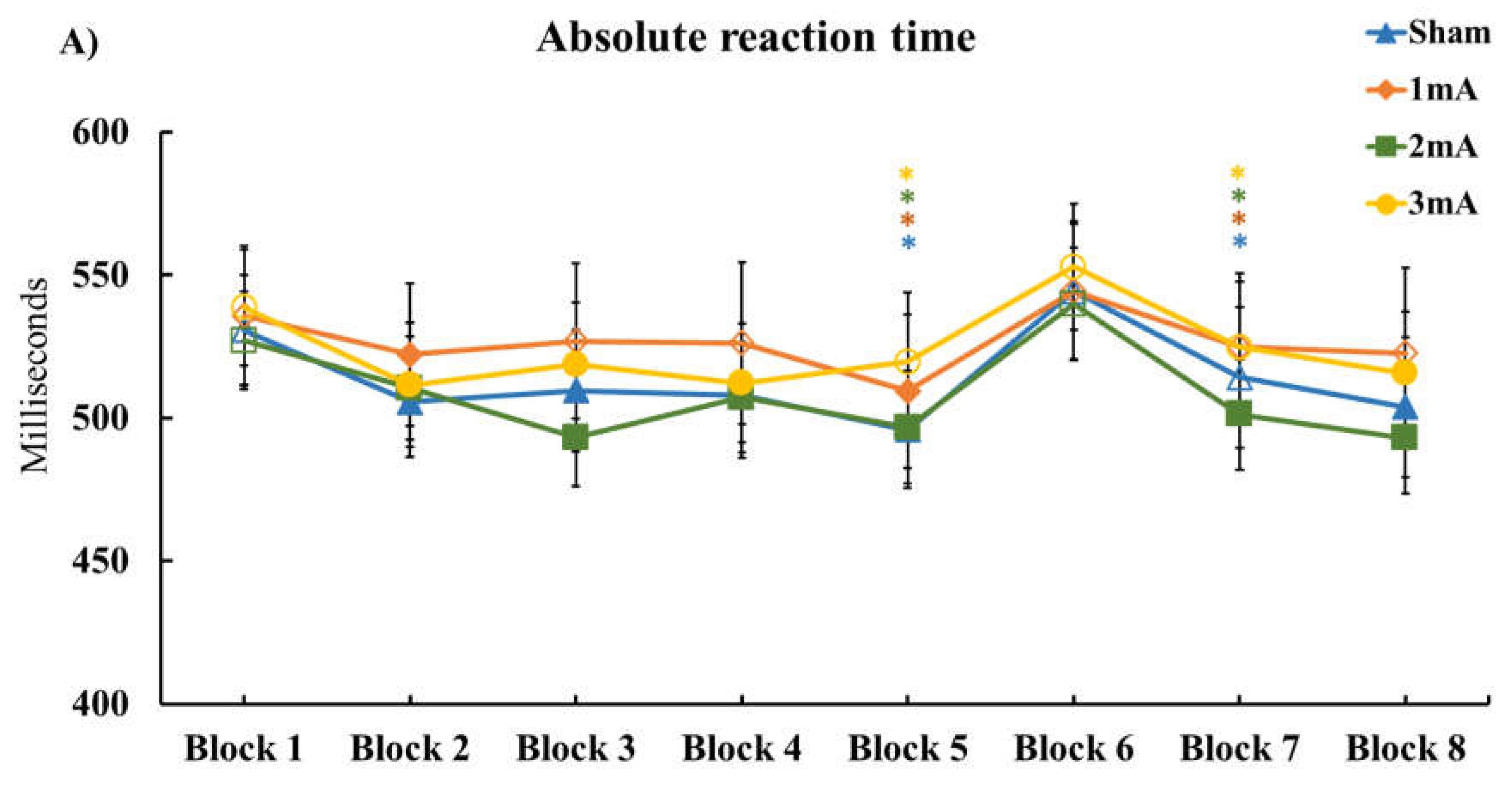
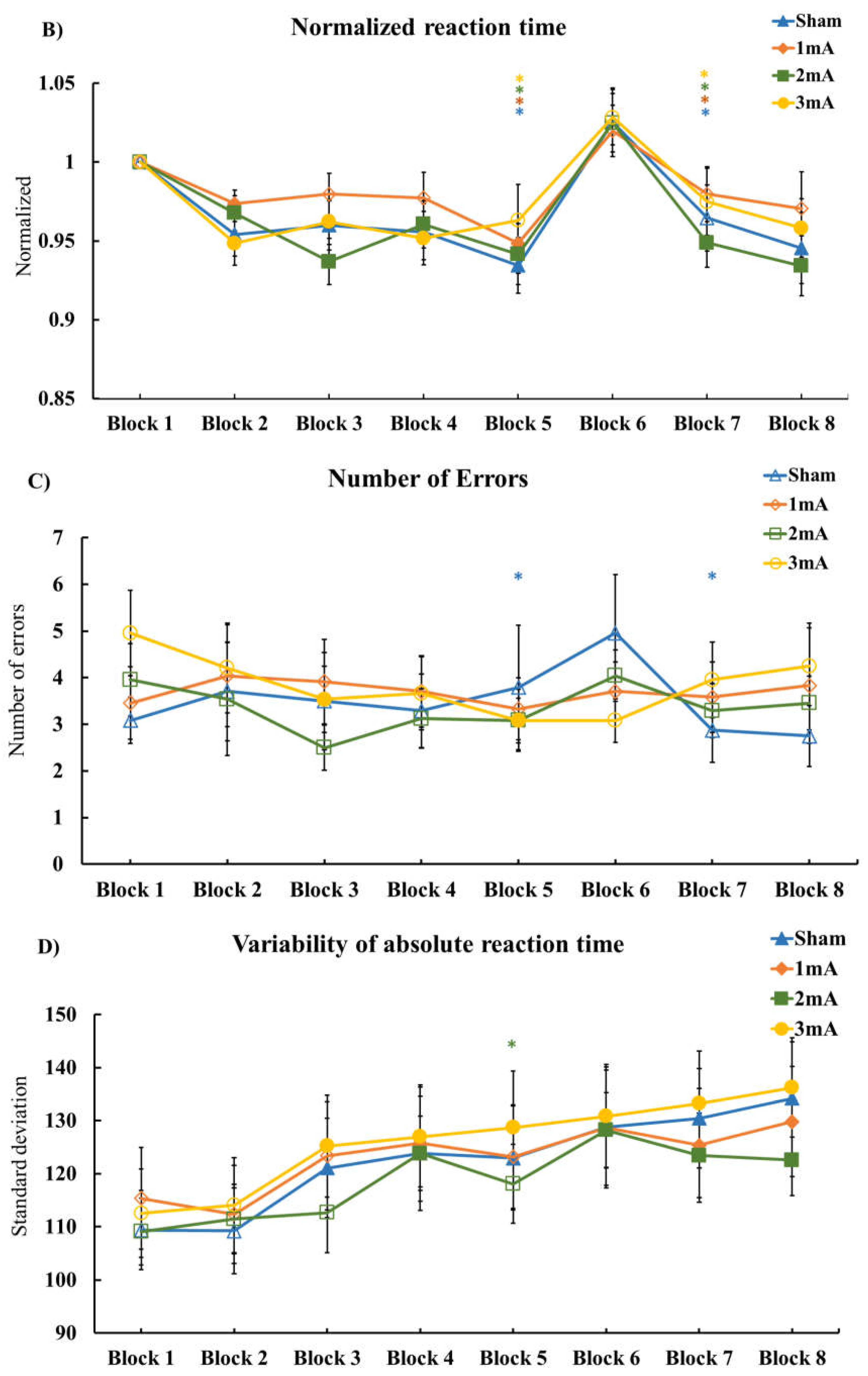
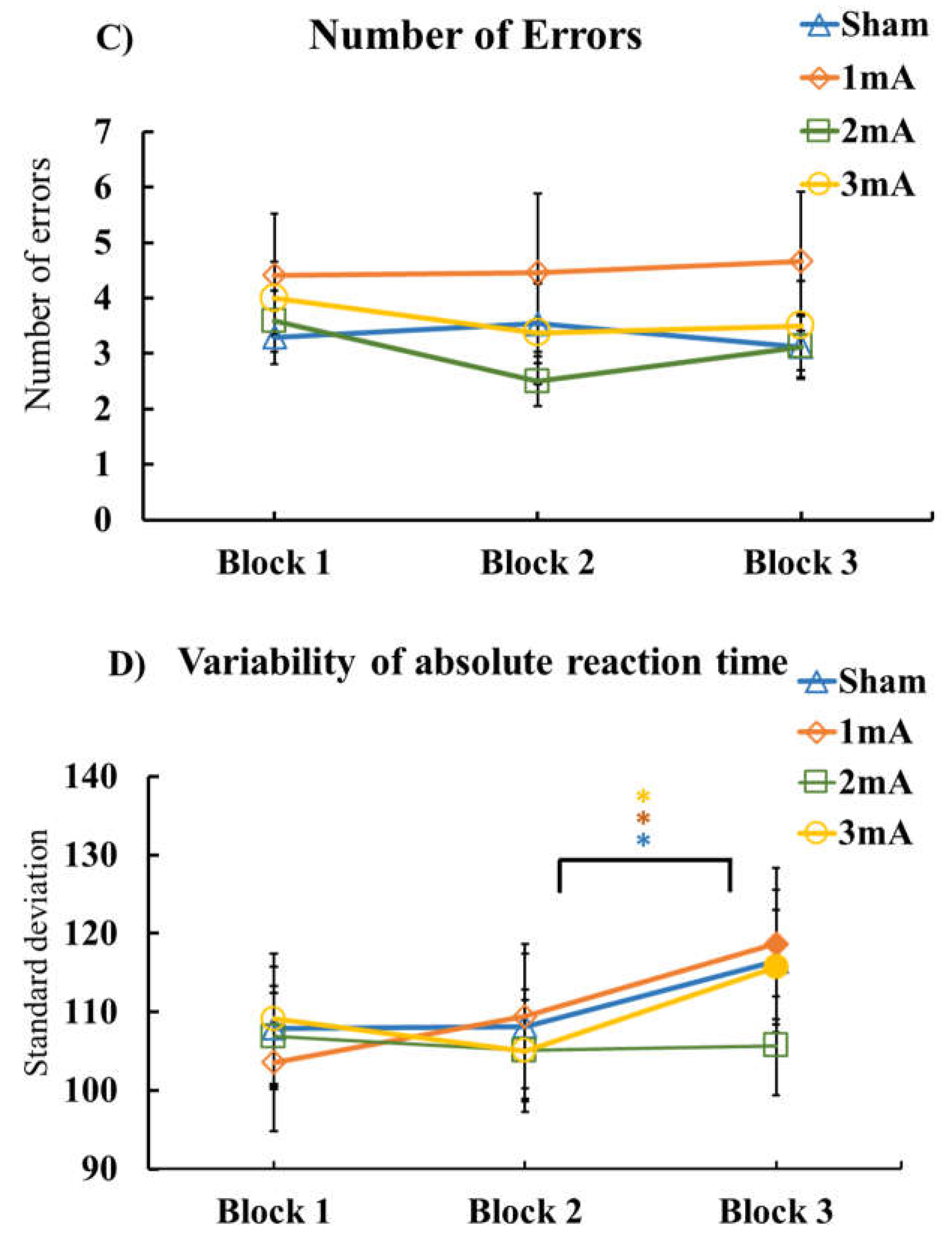
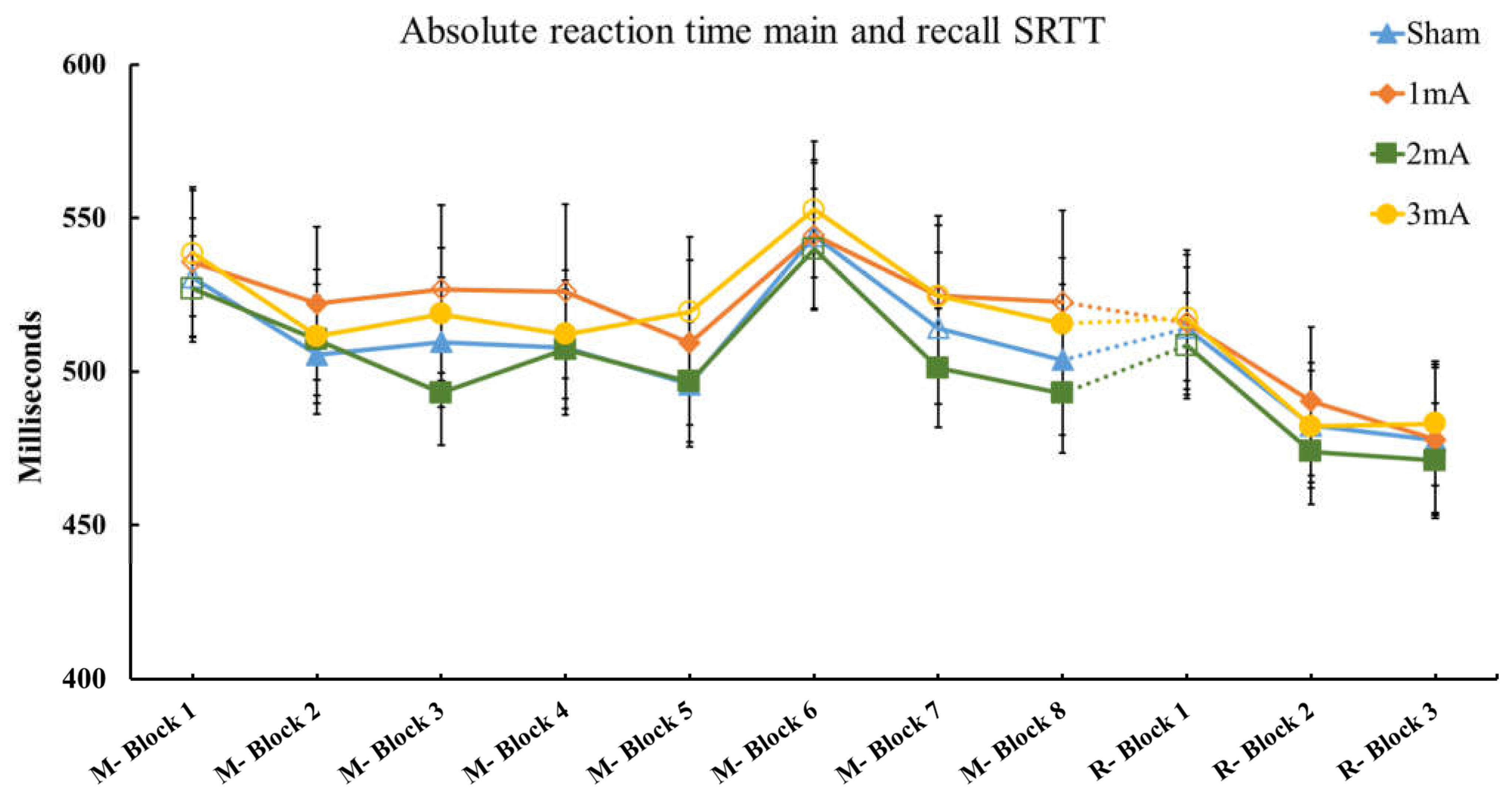
3.1. Effect of tDCS on M-SRTT
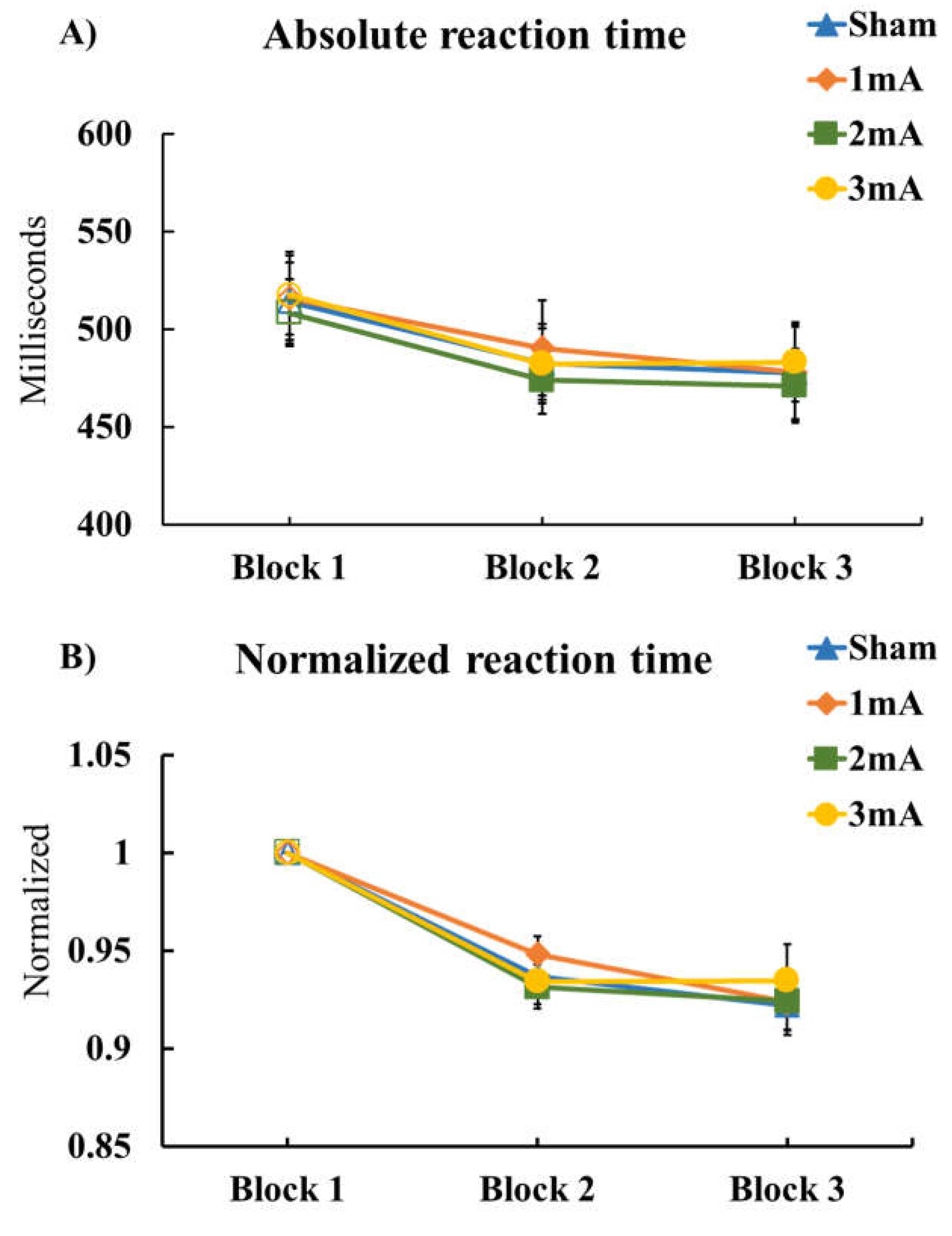
3.2. Effects of tDCS on R-SRTT
3.3. Assessment of tDCS Side Effects and Blinding Efficacy
4. Discussion
Limitations and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polanía, R.; Nitsche, M.A.; Ruff, C.C. Studying and modifying brain function with non-invasive brain stimulation. Nat. Neurosci. 2018, 21, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, P.B.; Brown, T.L.; Daskalakis, Z.J. The application of transcranial magnetic stimulation in psychiatry and neurosciences research. Acta Psychiatr. Scand. 2002, 105, 324–340. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-Z.; Edwards, M.J.; Rounis, E.; Bhatia, K.P.; Rothwell, J.C. Theta Burst Stimulation of the Human Motor Cortex. Neuron 2005, 45, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Rossini, P.M.; Rossini, L.; Ferreri, F. Brain-behavior relations: Transcranial magnetic stimulation: A review. IEEE Eng. Med. Biol. Mag. 2010, 29, 84–95. [Google Scholar] [CrossRef]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.G.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.B.; George, M.S.; et al. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application. An updated report from an I.F.C.N. Committee. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Cohen, L.G.; Wassermann, E.M.; Priori, A.; Lang, N.; Antal, A.; Paulus, W.; Hummel, F.; Boggio, P.S.; Fregni, F. Transcranial direct current stimulation: State of the art 2008. Brain Stimul. 2008, 1, 206–223. [Google Scholar] [CrossRef]
- Dissanayaka, T.; Zoghi, M.; Farrell, M.; Egan, G.F.; Jaberzadeh, S. Does transcranial electrical stimulation enhance corticospinal excitability of the motor cortex in healthy individuals? A systematic review and meta-analysis. Eur. J. Neurosci. 2017, 46, 1968–1990. [Google Scholar] [CrossRef]
- Antal, A.; Alekseichuk, I.; Bikson, M.; Brockmöller, J.; Brunoni, A.R.; Chen, R.; Cohen, L.G.; Dowthwaite, G.; Ellrich, J.; Flöel, A.; et al. Low intensity transcranial electric stimulation: Safety, ethical, legal regulatory and application guidelines. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2017, 128, 1774–1809. [Google Scholar] [CrossRef]
- Lefaucheur, J.-P.; Antal, A.; Ayache, S.S.; Benninger, D.H.; Brunelin, J.; Cogiamanian, F.; Cotelli, M.; De Ridder, D.; Ferrucci, R.; Langguth, B.; et al. Evidence-based guidelines on the therapeutic use of transcranial direct current stimulation (tDCS). Clin. Neurophysiol. 2017, 128, 56–92. [Google Scholar] [CrossRef]
- Hsu, W.Y.; Ku, Y.; Zanto, T.P.; Gazzaley, A. Effects of noninvasive brain stimulation on cognitive function in healthy aging and Alzheimer's disease: A systematic review and meta-analysis. Neurobiol. Aging 2015, 36, 2348–2359. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J. Physiol. 2000, 527, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Nitsche, M.S.; Klein, C.C.; Tergau, F.; Rothwell, J.C.; Paulus, W. Level of action of cathodal DC polarisation induced inhibition of the human motor cortex. Clin. Neurophysiol. 2003, 114, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Paulus, W. Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans. Neurology 2001, 57, 1899–1901. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Fricke, K.; Henschke, U.; Schlitterlau, A.; Liebetanz, D.; Lang, N.; Henning, S.; Tergau, F.; Paulus, W. Pharmacological modulation of cortical excitability shifts induced by transcranial direct current stimulation in humans. J. Physiol. 2003, 553, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Liebetanz, D.; Schlitterlau, A.; Henschke, U.; Fricke, K.; Frommann, K.; Lang, N.; Henning, S.; Paulus, W.; Tergau, F. GABAergic modulation of DC stimulation-induced motor cortex excitability shifts in humans. Eur. J. Neurosci. 2004, 19, 2720–2726. [Google Scholar] [CrossRef] [PubMed]
- Stagg, C.J. Magnetic Resonance Spectroscopy as a tool to study the role of GABA in motor-cortical plasticity. NeuroImage 2014, 86, 19–27. [Google Scholar] [CrossRef]
- Stagg, C.J.; Antal, A.; Nitsche, M.A. Physiology of Transcranial Direct Current Stimulation. J. ECT 2018, 34, 144–152. [Google Scholar] [CrossRef]
- Stagg, C.J.; Nitsche, M.A. Physiological basis of transcranial direct current stimulation. Neurosci. Rev. J. Bringing Neurobiol. Neurol. Psychiatry 2011, 17, 37–53. [Google Scholar] [CrossRef]
- Malenka, R.C.; Bear, M.F. LTP and LTD: An embarrassment of riches. Neuron 2004, 44, 5–21. [Google Scholar] [CrossRef]
- Burke, S.N.; Barnes, C.A. Neural plasticity in the ageing brain. Nat. Rev. Neurosci. 2006, 7, 30–40. [Google Scholar] [CrossRef]
- Rioult-Pedotti, M.-S.; Friedman, D.; Donoghue, J.P. Learning-induced LTP in neocortex. Science 2000, 290, 533–536. [Google Scholar] [CrossRef]
- Levasseur-Moreau, J.; Brunelin, J.; Fecteau, S. Non-invasive brain stimulation can induce paradoxical facilitation. Are these neuroenhancements transferable and meaningful to security services? Front. Hum. Neurosci. 2013, 7, 449. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Shanmugalingam, A.; McIntyre, A.; Burhan, A.M. The effect of non-invasive brain stimulation (nibs) on attention and memory function in stroke rehabilitation patients: A systematic review and meta-analysis. Diagnostics 2021, 11, 227. [Google Scholar] [CrossRef] [PubMed]
- Morishita, T.; Hummel, F.C. Non-invasive Brain Stimulation (NIBS) in Motor Recovery After Stroke: Concepts to Increase Efficacy. Curr. Behav. Neurosci. Rep. 2017, 4, 280–289. [Google Scholar] [CrossRef]
- Šimko, P.; Kent, J.A.; Rektorova, I. Is non-invasive brain stimulation effective for cognitive enhancement in Alzheimer’s disease? An updated meta-analysis. Clin. Neurophysiol. 2022, 144, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Flöel, A. tDCS-enhanced motor and cognitive function in neurological diseases. Neuroimage 2014, 85, 934–947. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Battaglia, S.; Garofalo, S.; Tortora, F.; Avenanti, A.; di Pellegrino, G. State-Dependent TMS over Prefrontal Cortex Disrupts Fear-Memory Reconsolidation and Prevents the Return of Fear. Curr. Biol. 2020, 30, 3672–3679.e3674. [Google Scholar] [CrossRef]
- Garofalo, S.; Battaglia, S.; Starita, F.; di Pellegrino, G. Modulation of cue-guided choices by transcranial direct current stimulation. Cortex 2021, 137, 124–137. [Google Scholar] [CrossRef]
- Zec, R.F. The neuropsychology of aging. Exp. Gerontol. 1995, 30, 431–442. [Google Scholar] [CrossRef]
- Alexander, G.; Ryan, L.; Bowers, D.; Foster, T.; Bizon, J.; Geldmacher, D.; Glisky, E. Characterizing cognitive aging in humans with links to animal models. Front. Aging Neurosci. 2012, 4, 21. [Google Scholar] [CrossRef]
- Summers, J.J.; Kang, N.; Cauraugh, J.H. Does transcranial direct current stimulation enhance cognitive and motor functions in the ageing brain? A systematic review and meta- analysis. Ageing Res. Rev. 2016, 25, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, A.; Radhu, N.; Farzan, F.; Mulsant, B.H.; Rajji, T.K.; Daskalakis, Z.J.; Blumberger, D.M. A meta-analysis of the effects of aging on motor cortex neurophysiology assessed by transcranial magnetic stimulation. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2016, 127, 2834–2845. [Google Scholar] [CrossRef] [PubMed]
- Calautti, C.; Serrati, C.; Baron, J.-C. Effects of Age on Brain Activation During Auditory-Cued Thumb-to-Index Opposition. Stroke 2001, 32, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Bedard, A.C.; Nichols, S.; Barbosa, J.A.; Schachar, R.; Logan, G.D.; Tannock, R. The development of selective inhibitory control across the life span. Dev. Neuropsychol. 2002, 21, 93–111. [Google Scholar] [CrossRef]
- Welford, A.T. Reaction Time, Speed of Performance, and Age. Ann. N. Y. Acad. Sci. 1988, 515, 1–17. [Google Scholar] [CrossRef]
- Swinnen, S.; Verschueren, S.; Bogaerts, H.; Dounskaia, N.; Lee, T.; Stelmach, G.; Serrien, D. Age-related deficits in motor learning and differences in feedback processing during the production of a bimanual coordination pattern. Cogn. Neuropsychol. 1998, 15, 439–466. [Google Scholar] [CrossRef]
- Serrien, D.J.; Swinnen, S.P.; Stelmach, G.E. Age-related deterioration of coordinated interlimb behavior. J. Gerontol. B Psychol. Sci. Soc. Sci. 2000, 55, P295–P303. [Google Scholar] [CrossRef]
- Heuninckx, S.; Debaere, F.; Wenderoth, N.; Verschueren, S.; Swinnen, S.P. Ipsilateral coordination deficits and central processing requirements associated with coordination as a function of aging. J. Gerontol. B Psychol. Sci. Soc. Sci. 2004, 59, P225–P232. [Google Scholar] [CrossRef]
- Ruch, F.L. The Differentiative Effects of Age upon Human Learning. J. Gen. Psychol. 1934, 11, 261–286. [Google Scholar] [CrossRef]
- McNay, E.C.; Willingham, D.B. Deficit in learning of a motor skill requiring strategy, but not of perceptuomotor recalibration, with aging. Learn. Mem. 1998, 4, 411–420. [Google Scholar] [CrossRef]
- Clark, B.C.; Manini, T.M. Sarcopenia =/= dynapenia. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Seidler, R.D.; Bernard, J.A.; Burutolu, T.B.; Fling, B.W.; Gordon, M.T.; Gwin, J.T.; Kwak, Y.; Lipps, D.B. Motor control and aging: Links to age-related brain structural, functional, and biochemical effects. Neurosci. Biobehav. Rev. 2010, 34, 721–733. [Google Scholar] [CrossRef]
- Buckles, V.D. Age-Related Slowing. In Sensorimotor Impairment in the Elderly; Stelmach, G.E., Hömberg, V., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 73–87. [Google Scholar]
- Darbin, O. The aging striatal dopamine function. Park. Relat. Disord. 2012, 18, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Heise, K.F.; Zimerman, M.; Hoppe, J.; Gerloff, C.; Wegscheider, K.; Hummel, F.C. The aging motor system as a model for plastic changes of GABA-mediated intracortical inhibition and their behavioral relevance. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 9039–9049. [Google Scholar] [CrossRef]
- Zhang, Z.; Andersen, A.; Smith, C.; Grondin, R.; Gerhardt, G.; Gash, D. Motor Slowing and Parkinsonian Signs in Aging Rhesus Monkeys Mirror Human Aging. J. Gerontol. Ser. A 2000, 55, B473–B480. [Google Scholar] [CrossRef] [PubMed]
- Raw, R.K.; Allen, R.J.; Mon-Williams, M.; Wilkie, R.M. Motor Sequence Learning in Healthy Older Adults Is Not Necessarily Facilitated by Transcranial Direct Current Stimulation (tDCS). Geriatrics 2016, 1, 32. [Google Scholar] [CrossRef]
- Prehn, K.; Flöel, A. Potentials and limits to enhance cognitive functions in healthy and pathological aging by tDCS. Front. Cell. Neurosci. 2015, 9, 355. [Google Scholar] [CrossRef]
- Bennabi, D.; Pedron, S.; Haffen, E.; Monnin, J.; Peterschmitt, Y.; Van Waes, V. Transcranial direct current stimulation for memory enhancement: From clinical research to animal models. Front. Syst. Neurosci. 2014, 8, 159. [Google Scholar] [CrossRef]
- Halsband, U.; Lange, R.K. Motor learning in man: A review of functional and clinical studies. J. Physiol. Paris 2006, 99, 414–424. [Google Scholar] [CrossRef]
- Doyon, J.; Bellec, P.; Amsel, R.; Penhune, V.; Monchi, O.; Carrier, J.; Lehéricy, S.; Benali, H. Contributions of the basal ganglia and functionally related brain structures to motor learning. Behav. Brain Res. 2009, 199, 61–75. [Google Scholar] [CrossRef]
- Ammann, C.; Spampinato, D.; Márquez-Ruiz, J. Modulating Motor Learning through Transcranial Direct-Current Stimulation: An Integrative View. Front. Psychol. 2016, 7, 1981. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.; Ivry, R.B. Multiple systems for motor skill learning. WIREs Cogn. Sci. 2010, 1, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Kelley, A.E.; Andrzejewski, M.E.; Baldwin, A.E.; Hernandez, P.J.; Pratt, W.E. Glutamate-Mediated Plasticity in Corticostriatal Networks. Ann. N. Y. Acad. Sci. 2003, 1003, 159–168. [Google Scholar] [CrossRef]
- Kim, S.; Stephenson, M.C.; Morris, P.G.; Jackson, S.R. tDCS-induced alterations in GABA concentration within primary motor cortex predict motor learning and motor memory: A 7 T magnetic resonance spectroscopy study. NeuroImage 2014, 99, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Castro-Alamancos, M.A.; Connors, B.W. Short-term synaptic enhancement and long-term potentiation in neocortex. Proc. Natl. Acad. Sci. USA 1996, 93, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Pleger, B.; Schwenkreis, P.; Dinse, H.; Ragert, P.; Höffken, O.; Malin, J.-P.; Tegenthoff, M. Pharmacological suppression of plastic changes in human primary somatosensory cortex after motor learning. Exp. Brain Res. Exp. Hirnforschung. Expérimentation Cérébrale 2003, 148, 525–532. [Google Scholar] [CrossRef]
- Amadi, U.; Allman, C.; Johansen-Berg, H.; Stagg, C.J. The Homeostatic Interaction Between Anodal Transcranial Direct Current Stimulation and Motor Learning in Humans is Related to GABAA Activity. Brain Stimul. 2015, 8, 898–905. [Google Scholar] [CrossRef]
- Roalf, D.R.; Sydnor, V.J.; Woods, M.; Wolk, D.A.; Scott, J.C.; Reddy, R.; Moberg, P.J. A quantitative meta-analysis of brain glutamate metabolites in aging. Neurobiol. Aging 2020, 95, 240–249. [Google Scholar] [CrossRef]
- Segovia, G.; Del Arco, A.; Mora, F. Effects of Aging on the Interaction Between Glutamate, Dopamine, and GABA in Striatum and Nucleus Accumbens of the Awake Rat. J. Neurochem. 1999, 73, 2063–2072. [Google Scholar] [CrossRef]
- Antonenko, D.; Thielscher, A.; Saturnino, G.B.; Aydin, S.; Ittermann, B.; Grittner, U.; Flöel, A. Towards precise brain stimulation: Is electric field simulation related to neuromodulation? Brain Stimul. 2019, 12, 1159–1168. [Google Scholar] [CrossRef]
- Wenk, G.L.; Walker, L.C.; Price, D.L.; Cork, L.C. Loss of NMDA, but not GABA-A, binding in the brains of aged rats and monkeys. Neurobiol. Aging 1991, 12, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Levin, O.; Fujiyama, H.; Boisgontier, M.P.; Swinnen, S.P.; Summers, J.J. Aging and motor inhibition: A converging perspective provided by brain stimulation and imaging approaches. Neurosci. Biobehav. Rev. 2014, 43, 100–117. [Google Scholar] [CrossRef]
- Gleichmann, M.; Chow, V.W.; Mattson, M.P. Homeostatic disinhibition in the aging brain and Alzheimer's disease. J. Alzheimer's Dis. 2011, 24, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Dumel, G.; Bourassa, M.-E.; Charlebois-Plante, C.; Desjardins, M.; Doyon, J.; Saint-Amour, D.; De Beaumont, L. Motor Learning Improvement Remains 3 Months After a Multisession Anodal tDCS Intervention in an Aging Population. Front. Aging Neurosci. 2018, 10, 335. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Schauenburg, A.; Lang, N.; Liebetanz, D.; Exner, C.; Paulus, W.; Tergau, F. Facilitation of implicit motor learning by weak transcranial direct current stimulation of the primary motor cortex in the human. J. Cogn. Neurosci. 2003, 15, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Galea, J.M.; Celnik, P. Brain polarization enhances the formation and retention of motor memories. J. Neurophysiol. 2009, 102, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Kantak, S.S.; Mummidisetty, C.K.; Stinear, J.W. Primary motor and premotor cortex in implicit sequence learning--evidence for competition between implicit and explicit human motor memory systems. Eur. J. Neurosci. 2012, 36, 2710–2715. [Google Scholar] [CrossRef]
- Kang, E.K.; Paik, N.-J. Effect of a tDCS electrode montage on implicit motor sequence learning in healthy subjects. Exp. Transl. Stroke Med. 2011, 3, 4. [Google Scholar] [CrossRef]
- Cuypers, K.; Leenus, D.J.F.; van den Berg, F.E.; Nitsche, M.A.; Thijs, H.; Wenderoth, N.; Meesen, R.L.J. Is motor learning mediated by tDCS intensity? PLoS ONE 2013, 8, e67344. [Google Scholar] [CrossRef]
- Ehsani, F.; Bakhtiary, A.H.; Jaberzadeh, S.; Talimkhani, A.; Hajihasani, A. Differential effects of primary motor cortex and cerebellar transcranial direct current stimulation on motor learning in healthy individuals: A randomized double-blind sham-controlled study. Neurosci. Res. 2016, 112, 10–19. [Google Scholar] [CrossRef]
- Reis, J.; Schambra, H.M.; Cohen, L.G.; Buch, E.R.; Fritsch, B.; Zarahn, E.; Celnik, P.A.; Krakauer, J.W. Noninvasive cortical stimulation enhances motor skill acquisition over multiple days through an effect on consolidation. Proc. Natl. Acad. Sci. USA 2009, 106, 1590–1595. [Google Scholar] [CrossRef]
- Stagg, C.J.; Jayaram, G.; Pastor, D.; Kincses, Z.T.; Matthews, P.M.; Johansen-Berg, H. Polarity and timing-dependent effects of transcranial direct current stimulation in explicit motor learning. Neuropsychologia 2011, 49, 800–804. [Google Scholar] [CrossRef]
- Kuo, M.-F.; Unger, M.; Liebetanz, D.; Lang, N.; Tergau, F.; Paulus, W.; Nitsche, M.A. Limited impact of homeostatic plasticity on motor learning in humans. Neuropsychologia 2008, 46, 2122–2128. [Google Scholar] [CrossRef]
- Buch, E.R.; Santarnecchi, E.; Antal, A.; Born, J.; Celnik, P.A.; Classen, J.; Gerloff, C.; Hallett, M.; Hummel, F.C.; Nitsche, M.A.; et al. Effects of tDCS on motor learning and memory formation: A consensus and critical position paper. Clin. Neurophysiol. 2017, 128, 589–603. [Google Scholar] [CrossRef]
- Hummel, F.C.; Heise, K.; Celnik, P.; Floel, A.; Gerloff, C.; Cohen, L.G. Facilitating skilled right hand motor function in older subjects by anodal polarization over the left primary motor cortex. Neurobiol. Aging 2010, 31, 2160–2168. [Google Scholar] [CrossRef]
- Zimerman, M.; Hummel, F.C. Non-invasive brain stimulation: Enhancing motor and cognitive functions in healthy old subjects. Front. Aging Neurosci. 2010, 2, 149. [Google Scholar] [CrossRef]
- Zimerman, M.; Nitsch, M.; Giraux, P.; Gerloff, C.; Cohen, L.G.; Hummel, F.C. Neuroenhancement of the aging brain: Restoring skill acquisition in old subjects. Ann. Neurol. 2013, 73, 10–15. [Google Scholar] [CrossRef]
- Rumpf, J.-J.; Wegscheider, M.; Hinselmann, K.; Fricke, C.; King, B.R.; Weise, D.; Klann, J.; Binkofski, F.; Buccino, G.; Karni, A.; et al. Enhancement of motor consolidation by post-training transcranial direct current stimulation in older people. Neurobiol. Aging 2017, 49, 1–8. [Google Scholar] [CrossRef]
- Fregni, F.; Boggio, P.S.; Santos, M.C.; Lima, M.; Vieira, A.L.; Rigonatti, S.P.; Silva, M.T.A.; Barbosa, E.R.; Nitsche, M.A.; Pascual-Leone, A. Noninvasive cortical stimulation with transcranial direct current stimulation in Parkinson's disease. Mov. Disord. 2006, 21, 1693–1702. [Google Scholar] [CrossRef]
- Andrade, S.M.; Ferreira, J.J.d.A.; Rufino, T.S.; Medeiros, G.; Brito, J.D.; da Silva, M.A.; Moreira, R.d.N. Effects of different montages of transcranial direct current stimulation on the risk of falls and lower limb function after stroke. Neurol. Res. 2017, 39, 1037–1043. [Google Scholar] [CrossRef]
- Saeys, W.; Vereeck, L.; Lafosse, C.; Truijen, S.; Wuyts, F.L.; Van De Heyning, P. Transcranial direct current stimulation in the recovery of postural control after stroke: A pilot study. Disabil. Rehabil. 2015, 37, 1857–1863. [Google Scholar] [CrossRef]
- Yosephi, M.H.; Ehsani, F.; Zoghi, M.; Jaberzadeh, S. Multi-session anodal tDCS enhances the effects of postural training on balance and postural stability in older adults with high fall risk: Primary motor cortex versus cerebellar stimulation. Brain Stimul. 2018, 11, 1239–1250. [Google Scholar] [CrossRef]
- Fregni, F.; Boggio, P.S.; Mansur, C.G.; Wagner, T.; Ferreira, M.J.; Lima, M.C.; Rigonatti, S.P.; Marcolin, M.A.; Freedman, S.D.; Nitsche, M.A.; et al. Transcranial direct current stimulation of the unaffected hemisphere in stroke patients. Neuroreport 2005, 16, 1551–1555. [Google Scholar] [CrossRef]
- Rossi, C.; Sallustio, F.; Di Legge, S.; Stanzione, P.; Koch, G. Transcranial direct current stimulation of the affected hemisphere does not accelerate recovery of acute stroke patients. Eur. J. Neurol. 2013, 20, 202–204. [Google Scholar] [CrossRef]
- Koini, M.; Duering, M.; Gesierich, B.G.; Rombouts, S.A.R.B.; Ropele, S.; Wagner, F.; Enzinger, C.; Schmidt, R. Grey-matter network disintegration as predictor of cognitive and motor function with aging. Brain Struct. Funct. 2018, 223, 2475–2487. [Google Scholar] [CrossRef]
- Nandi, T.; Puonti, O.; Clarke, W.T.; Nettekoven, C.; Barron, H.C.; Kolasinski, J.; Hanayik, T.; Hinson, E.L.; Berrington, A.; Bachtiar, V.; et al. tDCS induced GABA change is associated with the simulated electric field in M1, an effect mediated by grey matter volume in the MRS voxel. Brain Stimul. 2022, 15, 1153–1162. [Google Scholar] [CrossRef]
- Fathi, D.; Ueki, Y.; Mima, T.; Koganemaru, S.; Nagamine, T.; Tawfik, A.; Fukuyama, H. Effects of aging on the human motor cortical plasticity studied by paired associative stimulation. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2010, 121, 90–93. [Google Scholar] [CrossRef]
- Foster, T.C. Involvement of hippocampal synaptic plasticity in age-related memory decline. Brain Res. Brain Res. Rev. 1999, 30, 236–249. [Google Scholar] [CrossRef]
- Ghasemian-Shirvan, E.; Farnad, L.; Mosayebi-Samani, M.; Verstraelen, S.; Meesen, R.L.J.; Kuo, M.-F.; Nitsche, M.A. Age-related differences of motor cortex plasticity in adults: A transcranial direct current stimulation study. Brain Stimul. 2020, 13, 1588–1599. [Google Scholar] [CrossRef]
- Freitas, C.; Farzan, F.; Pascual-Leone, A. Assessing brain plasticity across the lifespan with transcranial magnetic stimulation: Why, how, and what is the ultimate goal? Front. Neurosci. 2013, 7, 42. [Google Scholar] [CrossRef]
- Balietti, M.; Tamagnini, F.; Fattoretti, P.; Burattini, C.; Casoli, T.; Platano, D.; Lattanzio, F.; Aicardi, G. Impairments of synaptic plasticity in aged animals and in animal models of Alzheimer's disease. Rejuvenation Res. 2012, 15, 235–238. [Google Scholar] [CrossRef]
- Farnad, L.; Ghasemian-Shirvan, E.; Mosayebi-Samani, M.; Kuo, M.-F.; Nitsche, M.A. Exploring and optimizing the neuroplastic effects of anodal transcranial direct current stimulation over the primary motor cortex of older humans. Brain Stimul. 2021, 14, 622–634. [Google Scholar] [CrossRef]
- Bikson, M.; Grossman, P.; Thomas, C.; Zannou, A.L.; Jiang, J.; Adnan, T.; Mourdoukoutas, A.P.; Kronberg, G.; Truong, D.; Boggio, P.; et al. Safety of Transcranial Direct Current Stimulation: Evidence Based Update 2016. Brain Stimul. 2016, 9, 641–661. [Google Scholar] [CrossRef]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Nasreddine, Z.S.; Phillips, N.A.; Bédirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The Montreal Cognitive Assessment, MoCA: A brief screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef]
- Association, W.M. World Medical Association Declaration of Helsinki: Ethical Principles for Medical Research Involving Human Subjects. JAMA 2013, 310, 2191–2194. [Google Scholar] [CrossRef]
- Guleyupoglu, B.; Febles, N.; Minhas, P.; Hahn, C.; Bikson, M. Reduced discomfort during high-definition transcutaneous stimulation using 6% benzocaine. Front. Neuroeng. 2014, 7, 28. [Google Scholar] [CrossRef]
- Robertson, E.M. The Serial Reaction Time Task: Implicit Motor Skill Learning? J. Neurosci. 2007, 27, 10073–10075. [Google Scholar] [CrossRef] [PubMed]
- Poreisz, C.; Boros, K.; Antal, A.; Paulus, W. Safety aspects of transcranial direct current stimulation concerning healthy subjects and patients. Brain Res. Bull. 2007, 72, 208–214. [Google Scholar] [CrossRef]
- Brunoni, A.R.; Amadera, J.; Berbel, B.; Volz, M.S.; Rizzerio, B.G.; Fregni, F. A systematic review on reporting and assessment of adverse effects associated with transcranial direct current stimulation. Int. J. Neuropsychopharmacol. 2011, 14, 1133–1145. [Google Scholar] [CrossRef]
- Savic, B.; Meier, B. How Transcranial Direct Current Stimulation Can Modulate Implicit Motor Sequence Learning and Consolidation: A Brief Review. Front. Hum. Neurosci. 2016, 10, 26. [Google Scholar] [CrossRef]
- López-Alonso, V.; Cheeran, B.; Fernández-del-Olmo, M. Relationship Between Non-invasive Brain Stimulation-induced Plasticity and Capacity for Motor Learning. Brain Stimul. 2015, 8, 1209–1219. [Google Scholar] [CrossRef]
- Bachtiar, V.; Stagg, C.J. The role of inhibition in human motor cortical plasticity. Neuroscience 2014, 278, 93–104. [Google Scholar] [CrossRef]
- King, B.R.; Rumpf, J.J.; Verbaanderd, E.; Heise, K.F.; Dolfen, N.; Sunaert, S.; Doyon, J.; Classen, J.; Mantini, D.; Puts, N.A.J.; et al. Baseline sensorimotor GABA levels shape neuroplastic processes induced by motor learning in older adults. Hum. Brain Mapp. 2020, 41, 3680–3695. [Google Scholar] [CrossRef]
- Kolasinski, J.; Hinson, E.L.; Divanbeighi Zand, A.P.; Rizov, A.; Emir, U.E.; Stagg, C.J. The dynamics of cortical GABA in human motor learning. J. Physiol. 2019, 597, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Stagg, C.J.; Bachtiar, V.; Johansen-Berg, H. The Role of GABA in Human Motor Learning. Curr. Biol. 2011, 21, 480–484. [Google Scholar] [CrossRef]
- Antonenko, D.; Schubert, F.; Bohm, F.; Ittermann, B.; Aydin, S.; Hayek, D.; Grittner, U.; Floel, A. tDCS-Induced Modulation of GABA Levels and Resting-State Functional Connectivity in Older Adults. J. Neurosci. Off. J. Soc. Neurosci. 2017, 37, 4065–4073. [Google Scholar] [CrossRef]
- Floyer-Lea, A.; Wylezinska, M.; Kincses, T.; Matthews, P.M. Rapid modulation of GABA concentration in human sensorimotor cortex during motor learning. J. Neurophysiol. 2006, 95, 1639–1644. [Google Scholar] [CrossRef] [PubMed]
- Berghuis, K.M.M.; De Rond, V.; Zijdewind, I.; Koch, G.; Veldman, M.P.; Hortobágyi, T. Neuronal mechanisms of motor learning are age dependent. Neurobiol. Aging 2016, 46, 149–159. [Google Scholar] [CrossRef]
- Cuypers, K.; Verstraelen, S.; Maes, C.; Hermans, L.; Hehl, M.; Heise, K.F.; Chalavi, S.; Mikkelsen, M.; Edden, R.; Levin, O.; et al. Task-related measures of short-interval intracortical inhibition and GABA levels in healthy young and older adults: A multimodal TMS-MRS study. NeuroImage 2020, 208, 116470. [Google Scholar] [CrossRef]
- Chang, L.; Jiang, C.S.; Ernst, T. Effects of age and sex on brain glutamate and other metabolites. Magn. Reson. Imaging 2009, 27, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Hadel, S.; Wirth, C.; Rapp, M.; Gallinat, J.; Schubert, F. Effects of age and sex on the concentrations of glutamate and glutamine in the human brain. J. Magn. Reson. Imaging JMRI 2013, 38, 1480–1487. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, L.G.; Schuff, N.; Cashdollar, N.; Weiner, M.W. Age-related glutamate and glutamine concentration changes in normal human brain: 1H MR spectroscopy study at 4 T. Neurobiol. Aging 2005, 26, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Sawaki, L.; Yaseen, Z.; Kopylev, L.; Cohen, L.G. Age-dependent changes in the ability to encode a novel elementary motor memory. Ann. Neurol. 2003, 53, 521–524. [Google Scholar] [CrossRef]
- Fujiyama, H.; Garry, M.I.; Levin, O.; Swinnen, S.P.; Summers, J.J. Age-related differences in inhibitory processes during interlimb coordination. Brain Res. 2009, 1262, 38–47. [Google Scholar] [CrossRef]
- Rogasch, N.C.; Dartnall, T.J.; Cirillo, J.; Nordstrom, M.A.; Semmler, J.G. Corticomotor plasticity and learning of a ballistic thumb training task are diminished in older adults. J. Appl. Physiol. 2009, 107, 1874–1883. [Google Scholar] [CrossRef]
- Antal, A.; Nitsche, M.A.; Kruse, W.; Kincses, T.Z.; Hoffmann, K.-P.; Paulus, W. Direct current stimulation over V5 enhances visuomotor coordination by improving motion perception in humans. J. Cogn. Neurosci. 2004, 16, 521–527. [Google Scholar] [CrossRef]
- Goh, J.O.S. Functional Dedifferentiation and Altered Connectivity in Older Adults: Neural Accounts of Cognitive Aging. Aging Dis. 2011, 2, 30–48. [Google Scholar] [PubMed]
- Goh, J.O.; Park, D.C. Neuroplasticity and cognitive aging: The scaffolding theory of aging and cognition. Restor. Neurol. Neurosci. 2009, 27, 391–403. [Google Scholar] [CrossRef]
- Mosayebi-Samani, M.; Jamil, A.; Salvador, R.; Ruffini, G.; Haueisen, J.; Nitsche, M.A. The impact of individual electrical fields and anatomical factors on the neurophysiological outcomes of tDCS: A TMS-MEP and MRI study. Brain Stimul. 2021, 14, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Polania, R.; Nitsche, M.A.; Paulus, W. Modulating functional connectivity patterns and topological functional organization of the human brain with transcranial direct current stimulation. Hum. Brain Mapp. 2011, 32, 1236–1249. [Google Scholar] [CrossRef]
- Cassady, K.; Gagnon, H.; Lalwani, P.; Simmonite, M.; Foerster, B.; Park, D.; Peltier, S.J.; Petrou, M.; Taylor, S.F.; Weissman, D.H.; et al. Sensorimotor network segregation declines with age and is linked to GABA and to sensorimotor performance. NeuroImage 2019, 186, 234–244. [Google Scholar] [CrossRef]
- Heuninckx, S.; Wenderoth, N.; Swinnen, S.P. Systems neuroplasticity in the aging brain: Recruiting additional neural resources for successful motor performance in elderly persons. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Galea, J.M.; Vazquez, A.; Pasricha, N.; de Xivry, J.J.; Celnik, P. Dissociating the roles of the cerebellum and motor cortex during adaptive learning: The motor cortex retains what the cerebellum learns. Cereb. Cortex 2011, 21, 1761–1770. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | d.f., Error | F Value | P Value | ||
|---|---|---|---|---|---|
| (A) Absolute RT | Condition | 3, 69 | 0.827 | 0.035 | 0.484 |
| Block | 2.521, 57.977 | 13.486 | 0.370 | <0.001 * | |
| Condition × Block | 8.790, 202.169 | 1.290 | 0.053 | 0.245 | |
| (B) Normalized RT | Condition | 3, 69 | 0.572 | 0.024 | 0.635 |
| Block | 2.502, 57.539 | 14.996 | 0.395 | <0.001 * | |
| Condition × Block | 8.982, 206.595 | 1.192 | 0.049 | 0.302 | |
| (C) Errors | Condition | 1.785, 41.044 | 0.408 | 0.017 | 0.645 |
| Block | 2.639, 60.695 | 0.637 | 0.027 | 0.575 | |
| Condition × Block | 3.763, 86.548 | 1.286 | 0.053 | 0.282 | |
| (D) Variability | Condition | 3, 69 | 0.538 | 0.023 | 0.658 |
| Block | 2.742, 63.069 | 11.787 | 0.339 | <0.001 * | |
| Condition × Block | 9.399, 216.171 | 0.743 | 0.031 | 0.788 |
| Factor | d.f., Error | F Value | P Value | ||
|---|---|---|---|---|---|
| (A) Absolute RT | Condition | 3, 69 | 0.351 | 0.015 | 0.789 |
| Block | 1.554, 35.751 | 42.761 | 0.650 | <0.001 * | |
| Condition × Block | 3.773, 86.780 | 0.517 | 0.022 | 0.713 | |
| (B) Normalized RT | Condition | 3, 69 | 0.139 | 0.006 | 0.936 |
| Block | 1.397, 32.128 | 41.488 | 0.643 | <0.001 * | |
| Condition × Block | 3.634, 83.587 | 0.508 | 0.022 | 0.712 | |
| (C) Errors | Condition | 1.779, 40.912 | 1.118 | 0.046 | 0.331 |
| Block | 2, 46 | 0.666 | 0.028 | 0.517 | |
| Condition × Block | 3.584, 82.437 | 0.598 | 0.025 | 0.647 | |
| (D) Variability | Condition | 3, 69 | 0.472 | 0.020 | 0.703 |
| Block | 1.565, 36.001 | 6.024 | 0.208 | 0.009 * | |
| Condition × Block | 6, 138 | 2.643 | 0.103 | 0.019* |
| Data Points | Guessed Stimulation Intensity | Total Data Points | ||||
|---|---|---|---|---|---|---|
| Sham | 1 mA | 2 mA | 3 mA | |||
| Real tDCS Intensity | Sham | 14 | 6 | 1 | 4 | 25 |
| 1 mA | 16 | 4 | 3 | 2 | 25 | |
| 2 mA | 13 | 6 | 2 | 4 | 25 | |
| 3 mA | 13 | 6 | 3 | 3 | 25 | |
| Side Effects | Sham | 1 mA | 2 mA | 3 mA |
|---|---|---|---|---|
| Visual Phenomena | 0.08 ± 0.40 | 0.04 ± 0.20 | 0.37 ± 1.27 | 0.12 ± 0.33 |
| Itching | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.12 ± 0.44 | 0.25 ± 0.67 |
| Tingling | 0.04 ± 0.20 | 0.04 ± 0.20 | 0.04 ± 0.20 | 0.16 ± 0.48 |
| Burning | 0.04 ± 0.20 | 0.12 ± 0.61 | 0.20 ± 0.65 | 0.20 ± 0.65 |
| Pain | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.12 ± 0.44 |
| Redness | 0.00 ± 0.00 | 0.16 ± 0.56 | 0.08 ± 0.40 | 0.08 ± 0.40 |
| Headache | 0.00 ± 0.00 | 0.12 ± 0.61 | 0.08 ± 0.40 | 0.04 ± 0.20 |
| Fatigue | 0.20 ± 0.50 | 0.12 ± 0.44 | 0.16 ± 0.48 | 0.08 ± 0.28 |
| Concentration | 0.08 ± 0.28 | 0.08 ± 0.28 | 0.20 ± 0.41 | 0.16 ± 0.38 |
| Nervousness | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.04 ± 0.20 |
| Sleep Problems | 0.00 ± 0.00 | 0.04 ± 0.20 | 0.08 ± 0.40 | 0.04 ± 0.20 |
| Side Effects | d.f., Error | F Value | P Value | ||
|---|---|---|---|---|---|
| During Stimulation | Visual phenomena | 1.216, 27.967 | 1.174 | 0.049 | 0.300 |
| Itching | 1.692, 38.922 | 2.213 | 0.088 | 0.094 | |
| Tingling | 1.000, 23.000 | 1.865 | 0.075 | 0.185 | |
| Burning | 1.690, 38.866 | 1.105 | 0.046 | 0.333 | |
| Pain | 1.000, 23.000 | 1.865 | 0.075 | 0.185 | |
| After Stimulation | Redness | 1.428, 32.840 | 0.793 | 0.033 | 0.422 |
| Headache | 1.000, 23.000 | 1.000 | 0.042 | 0.328 | |
| Fatigue | 2.284, 52.542 | 1.264 | 0.052 | 0.294 | |
| Concentration | 3, 69 | 1.084 | 0.045 | 0.362 | |
| Nervousness | 1.000, 23.000 | 1.000 | 0.042 | 0.328 | |
| Sleep Problems | 1.308, 30.088 | 0.561 | 0.024 | 0.505 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghasemian-Shirvan, E.; Ungureanu, R.; Melo, L.; van Dun, K.; Kuo, M.-F.; Nitsche, M.A.; Meesen, R.L.J. Optimizing the Effect of tDCS on Motor Sequence Learning in the Elderly. Brain Sci. 2023, 13, 137. https://doi.org/10.3390/brainsci13010137
Ghasemian-Shirvan E, Ungureanu R, Melo L, van Dun K, Kuo M-F, Nitsche MA, Meesen RLJ. Optimizing the Effect of tDCS on Motor Sequence Learning in the Elderly. Brain Sciences. 2023; 13(1):137. https://doi.org/10.3390/brainsci13010137
Chicago/Turabian StyleGhasemian-Shirvan, Ensiyeh, Ruxandra Ungureanu, Lorena Melo, Kim van Dun, Min-Fang Kuo, Michael A. Nitsche, and Raf L. J. Meesen. 2023. "Optimizing the Effect of tDCS on Motor Sequence Learning in the Elderly" Brain Sciences 13, no. 1: 137. https://doi.org/10.3390/brainsci13010137
APA StyleGhasemian-Shirvan, E., Ungureanu, R., Melo, L., van Dun, K., Kuo, M.-F., Nitsche, M. A., & Meesen, R. L. J. (2023). Optimizing the Effect of tDCS on Motor Sequence Learning in the Elderly. Brain Sciences, 13(1), 137. https://doi.org/10.3390/brainsci13010137