
Frequency-Tagging EEG of Superimposed Social and Non-Social Visual Stimulation Streams Provides No Support for Social Salience Enhancement after Intranasal Oxytocin Administration

 , , and
, , and
Abstract
:1. Introduction
1.1. Oxytocin as a Complex Modulator of Complex Social Behavior
1.2. The Social Salience Hypothesis of OT
1.3. Pinpointing the Neural Salience of Social Versus Non-Social Information via Frequency-Tagging EEG
1.4. The Present Study
2. Materials and Methods
2.1. Participants
2.2. Study Design
2.3. Procedure
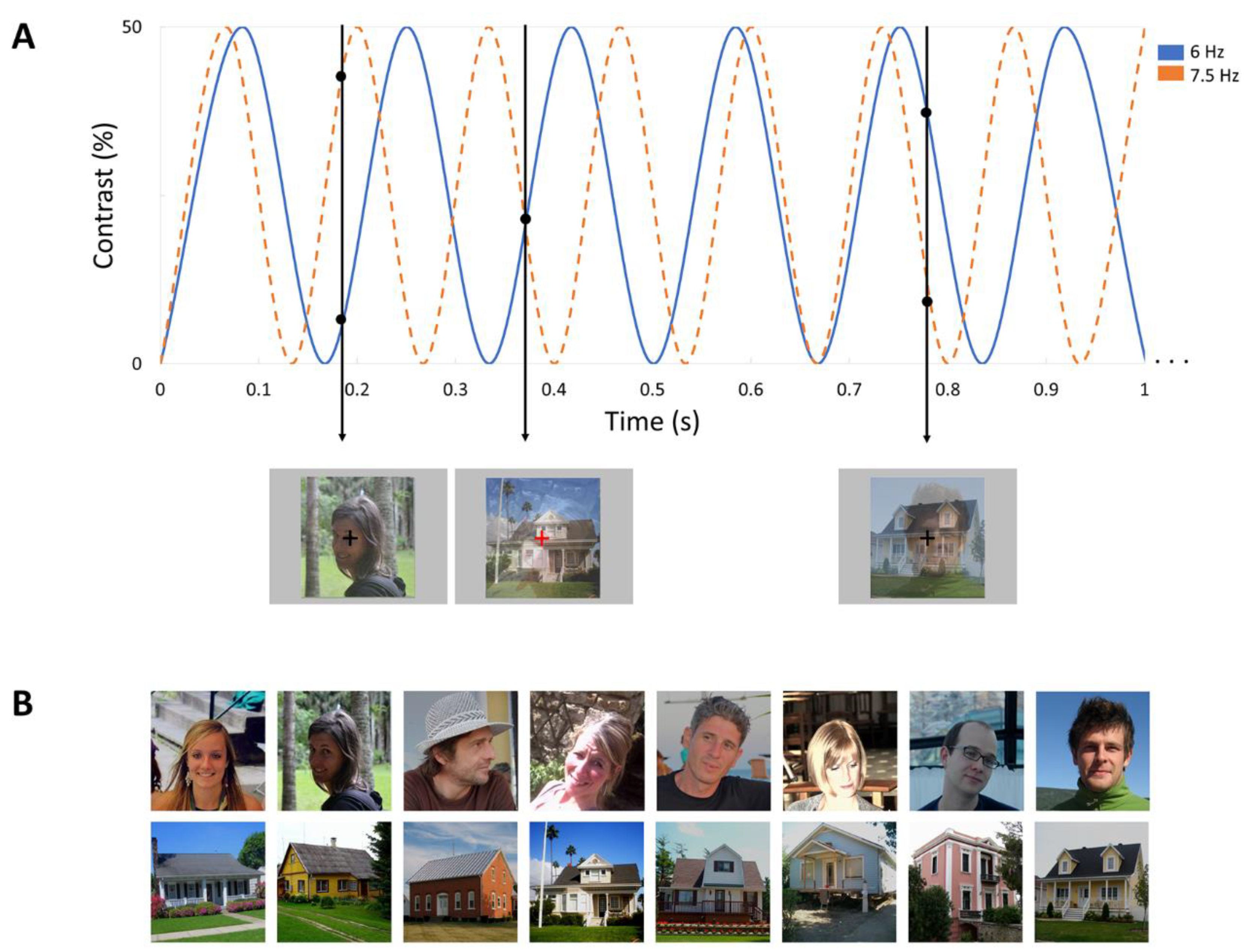
2.3.1. Stimuli
2.3.2. Frequency Tagging Paradigm
2.3.3. EEG Acquisition
2.4. Data Analysis
2.4.1. EEG Analysis
- Preprocessing
- Frequency-Domain Analysis
- Defining ROIs
2.4.2. Orthogonal Data Analysis
2.4.3. Statistical Analysis
3. Results
3.1. Orthogonal Task Performance
3.2. Neural Responses
3.2.1. Test–Retest Reliability and Power Analysis
3.2.2. Oxytocin Modulation Effect on Social and Non-Social Stimuli
4. Discussion
4.1. Quantifying Implicit Attentional Processing via Frequency-Tagging EEG
4.2. Topographical Selectivity for Responses toward Houses Versus Faces
4.3. Strong Biomarker Characteristics but No Treatment Effect
4.4. Lack of Treatment Effects Due to Baseline Ceiling Levels?
4.5. Lack of Treatment Effects Due to Lack of Personal Relevance?
4.6. Lack of Treatment Effect Because of Lack of OT Impact?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kosfeld, M.; Heinrichs, M.L.; Zak, P.J.; Fischbacher, U.; Fehr, E. Oxytocin increases trust in humans. Nature 2005, 435, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Zak, P.J.; Stanton, A.A.; Ahmadi, S. Oxytocin Increases Generosity in Humans. PLoS ONE 2007, 2, e1128. [Google Scholar] [CrossRef] [PubMed]
- De Dreu, C.K.W.; Greer, L.L.; Handgraaf, M.J.J.; Shalvi, S.; Van Kleef, G.A.; Baas, M.; Velden, F.S.T.; Van Dijk, E.; Feith, S.W.W. The Neuropeptide Oxytocin Regulates Parochial Altruism in Intergroup Conflict Among Humans. Science 2010, 328, 1408–1411. [Google Scholar] [CrossRef] [PubMed]
- Declerck, C.H.; Boone, C.; Kiyonari, T. Oxytocin and cooperation under conditions of uncertainty: The modulating role of incentives and social information. Horm. Behav. 2010, 57, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, T.; Heinrichs, M.; Vonlanthen, A.; Fischbacher, U.; Fehr, E. Oxytocin Shapes the Neural Circuitry of Trust and Trust Adaptation in Humans. Neuron 2008, 58, 639–650. [Google Scholar] [CrossRef]
- Marsh, A.A.; Yu, H.; Pine, D.S.; Blair, R.J.R. Oxytocin improves specific recognition of positive facial expressions. Psychopharmacology 2010, 209, 225–232. [Google Scholar] [CrossRef]
- Lischke, A.; Berger, C.; Prehn, K.; Heinrichs, M.; Herpertz, S.C.; Domes, G. Intranasal oxytocin enhances emotion recognition from dynamic facial expressions and leaves eye-gaze unaffected. Psychoneuroendocrinology 2012, 37, 475–481. [Google Scholar] [CrossRef]
- Domes, G.; Heinrichs, M.; Michel, A.; Berger, C.; Herpertz, S.C. Oxytocin Improves “Mind-Reading” in Humans. Biol. Psychiatry 2007, 61, 731–733. [Google Scholar] [CrossRef]
- Domes, G.; Steiner, A.; Porges, S.W.; Heinrichs, M. Oxytocin differentially modulates eye gaze to naturalistic social signals of happiness and anger. Psychoneuroendocrinology 2013, 38, 1198–1202. [Google Scholar] [CrossRef]
- Guastella, A.J.; Mitchell, P.B.; Dadds, M.R. Oxytocin Increases Gaze to the Eye Region of Human Faces. Biol. Psychiatry 2008, 63, 3–5. [Google Scholar] [CrossRef]
- Guastella, A.J.; Einfeld, S.L.; Gray, K.M.; Rinehart, N.J.; Tonge, B.J.; Lambert, T.J.; Hickie, I.B. Intranasal Oxytocin Improves Emotion Recognition for Youth with Autism Spectrum Disorders. Biol. Psychiatry 2010, 67, 692–694. [Google Scholar] [CrossRef] [PubMed]
- Dadds, M.R.; Macdonald, E.; Cauchi, A.; Williams, K.; Levy, F.; Brennan, J. Nasal Oxytocin for Social Deficits in Childhood Autism: A Randomized Controlled Trial. J. Autism Dev. Disord. 2013, 44, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Huang, X.; Ebstein, R.P.; Yu, R. Intranasal oxytocin in the treatment of autism spectrum disorders: A multilevel meta-analysis. Neurosci. Biobehav. Rev. 2021, 122, 18–27. [Google Scholar] [CrossRef]
- Bakermans-Kranenburg, M.J.; Van Ijzendoorn, M.H. Sniffing around oxytocin: Review and meta-analyses of trials in healthy and clinical groups with implications for pharmacotherapy. Transl. Psychiatry 2013, 3, e258. [Google Scholar] [CrossRef] [PubMed]
- Feifel, D.; MacDonald, K.; Cobb, P.; Minassian, A. Adjunctive intranasal oxytocin improves verbal memory in people with schizophrenia. Schizophr. Res. 2012, 139, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Simeon, D.; Bartz, J.; Hamilton, H.; Crystal, S.; Braun, A.; Ketay, S.; Hollander, E. Oxytocin administration attenuates stress reactivity in borderline personality disorder: A pilot study. Psychoneuroendocrinology 2011, 36, 1418–1421. [Google Scholar] [CrossRef] [PubMed]
- Shamay-Tsoory, S.G.; Fischer, M.; Dvash, J.; Harari, H.; Perach-Bloom, N.; Levkovitz, Y. Intranasal Administration of Oxytocin Increases Envy and Schadenfreude (Gloating). Biol. Psychiatry 2009, 66, 864–870. [Google Scholar] [CrossRef]
- Taylor, S.E.; Gonzaga, G.C.; Klein, L.C.; Hu, P.; Greendale, G.A.; Seeman, T.E. Relation of Oxytocin to Psychological Stress Responses and Hypothalamic-Pituitary-Adrenocortical Axis Activity in Older Women. Psychosom. Med. 2006, 68, 238–245. [Google Scholar] [CrossRef]
- Tabak, B.A.; McCullough, M.E.; Szeto, A.; Mendez, A.J.; McCabe, P.M. Oxytocin indexes relational distress following interpersonal harms in women. Psychoneuroendocrinology 2011, 36, 115–122. [Google Scholar] [CrossRef]
- Groppe, S.E.; Gossen, A.; Rademacher, L.; Hahn, A.; Westphal, L.; Gründer, G.; Spreckelmeyer, K.N. Oxytocin Influences Processing of Socially Relevant Cues in the Ventral Tegmental Area of the Human Brain. Biol. Psychiatry 2013, 74, 172–179. [Google Scholar] [CrossRef]
- Shamay-Tsoory, S.G.; Abu-Akel, A. The Social Salience Hypothesis of Oxytocin. Biol. Psychiatry 2016, 79, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Rimmele, U.; Hediger, K.; Heinrichs, M.; Klaver, P. Oxytocin Makes a Face in Memory Familiar. J. Neurosci. 2009, 29, 38–42. [Google Scholar] [CrossRef]
- Unkelbach, C.; Guastella, A.J.; Forgas, J.P. Oxytocin Selectively Facilitates Recognition of Positive Sex and Relationship Words. Psychol. Sci. 2008, 19, 1092–1094. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Qi, S.; Becker, B.; Luo, L.; Gao, S.; Gong, Q.; Hurlemann, R.; Kendrick, K.M. Oxytocin selectively facilitates learning with social feedback and increases activity and functional connectivity in emotional memory and reward processing regions. Hum. Brain Mapp. 2015, 36, 2132–2146. [Google Scholar] [CrossRef]
- Norman, G.J.; Cacioppo, J.T.; Morris, J.S.; Karelina, K.; Malarkey, W.B.; DeVries, A.C.; Berntson, G.G. Selective influences of oxytocin on the evaluative processing of social stimuli. J. Psychopharmacol. 2010, 25, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Tollenaar, M.S.; Chatzimanoli, M.; van der Wee, N.J.; Putman, P. Enhanced orienting of attention in response to emotional gaze cues after oxytocin administration in healthy young men. Psychoneuroendocrinology 2013, 38, 1797–1802. [Google Scholar] [CrossRef] [PubMed]
- Domes, G.; Sibold, M.; Schulze, L.; Lischke, A.; Herpertz, S.C.; Heinrichs, M. Intranasal oxytocin increases covert attention to positive social cues. Psychol. Med. 2012, 43, 1747–1753. [Google Scholar] [CrossRef]
- Baum, A.; Sachidanandam, R.; García-Sastre, A. Different Amygdala Subregions Mediate Valence- Related and Attentional Effects of Oxytocin in Humans. Proc. Natl. Acad. Sci. USA 2011, 108, 3092. [Google Scholar] [CrossRef]
- Leknes, S.; Wessberg, J.; Ellingsen, D.-M.; Chelnokova, O.; Olausson, H.; Laeng, B. Oxytocin enhances pupil dilation and sensitivity to ‘hidden’ emotional expressions. Soc. Cogn. Affect. Neurosci. 2012, 8, 741–749. [Google Scholar] [CrossRef]
- Hovey, D.; Martens, L.; Laeng, B.; Leknes, S.; Westberg, L. The effect of intranasal oxytocin on visual processing and salience of human faces. Transl. Psychiatry 2020, 10, 318. [Google Scholar] [CrossRef]
- Andari, E.; Richard, N.; Leboyer, M.; Sirigu, A. Adaptive coding of the value of social cues with oxytocin, an fMRI study in autism spectrum disorder. Cortex 2016, 76, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, P.; Esslinger, C.; Chen, Q.; Mier, D.; Lis, S.; Siddhanti, S.; Gruppe, H.; Mattay, V.S.; Gallhofer, B.; Meyer-Lindenberg, A. Oxytocin Modulates Neural Circuitry for Social Cognition and Fear in Humans. J. Neurosci. 2005, 25, 11489–11493. [Google Scholar] [CrossRef]
- Peltola, M.J.; Strathearn, L.; Puura, K. Oxytocin promotes face-sensitive neural responses to infant and adult faces in mothers. Psychoneuroendocrinology 2018, 91, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Huffmeijer, R.; Alink, L.R.; Tops, M.; Grewen, K.M.; Light, K.C.; Bakermans-Kranenburg, M.J.; van Ijzendoorn, M.H. The impact of oxytocin administration and maternal love withdrawal on event-related potential (ERP) responses to emotional faces with performance feedback. Horm. Behav. 2013, 63, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Tillman, R.; Gordon, I.; Naples, A.; Rolison, M.; Leckman, J.F.; Feldman, R.; Pelphrey, K.A.; McPartland, J.C. Oxytocin Enhances the Neural Efficiency of Social Perception. Front. Hum. Neurosci. 2019, 13, 71. [Google Scholar] [CrossRef] [PubMed]
- Adrian, E.D.; Matthews, B.H.C. The berger rhythm: Potential changes from the occipital lobes in man. Brain 1934, 57, 355–385. [Google Scholar] [CrossRef]
- Norcia, A.M.; Appelbaum, L.; Ales, J.M.; Cottereau, B.R.; Rossion, B. The steady-state visual evoked potential in vision research: A review. J. Vis. 2015, 15, 4. [Google Scholar] [CrossRef]
- Vettori, S.; Dzhelyova, M.; Van der Donck, S.; Jacques, C.; Van Wesemael, T.; Steyaert, J.; Rossion, B.; Boets, B. Combined frequency-tagging EEG and eye tracking reveal reduced social bias in boys with autism spectrum disorder. Cortex 2019, 125, 135–148. [Google Scholar] [CrossRef]
- Vettori, S.; Dzhelyova, M.; Van Der Donck, S.; Jacques, C.; Steyaert, J.; Rossion, B.; Boets, B. Frequency-Tagging Electroencephalography of Superimposed Social and Non-Social Visual Stimulation Streams Reveals Reduced Saliency of Faces in Autism Spectrum Disorder. Front. Psychiatry 2020, 11, 332. [Google Scholar] [CrossRef]
- Pei, F.; Pettet, M.W.; Norcia, A.M. Neural correlates of object-based attention. J. Vis. 2002, 2, 588–596. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Seth, A.K.; Gally, J.A.; Edelman, G.M. The power of human brain magnetoencephalographic signals can be modulated up or down by changes in an attentive visual task. Proc. Natl. Acad. Sci. USA 2003, 100, 3501–3506. [Google Scholar] [CrossRef]
- Müller, M.M.; Andersen, S.; Trujillo, N.J.; Valdés-Sosa, P.; Malinowski, P.; Hillyard, S.A. Feature-selective attention enhances color signals in early visual areas of the human brain. Proc. Natl. Acad. Sci. USA 2006, 103, 14250–14254. [Google Scholar] [CrossRef]
- Andersen, S.K.; Fuchs, S.; Müller, M.M. Effects of Feature-selective and Spatial Attention at Different Stages of Visual Processing. J. Cogn. Neurosci. 2011, 23, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Störmer, V.S.; Winther, G.N.; Li, S.-C.; Andersen, S.K. Sustained Multifocal Attentional Enhancement of Stimulus Processing in Early Visual Areas Predicts Tracking Performance. J. Neurosci. 2013, 33, 5346–5351. [Google Scholar] [CrossRef] [PubMed]
- Baldauf, D.; Desimone, R. Neural Mechanisms of Object-Based Attention. Science 2014, 344, 424–427. [Google Scholar] [CrossRef] [PubMed]
- de Heering, A.; Rossion, B. Rapid categorization of natural face images in the infant right hemisphere. eLife 2015, 4, e06564. [Google Scholar] [CrossRef]
- Van der Donck, S.; Dzhelyova, M.; Vettori, S.; Thielen, H.; Steyaert, J.; Rossion, B.; Boets, B. Fast Periodic Visual Stimulation EEG Reveals Reduced Neural Sensitivity to Fearful Faces in Children with Autism. J. Autism Dev. Disord. 2019, 49, 4658–4673. [Google Scholar] [CrossRef]
- Van Der Donck, S.; Dzhelyova, M.; Vettori, S.; Mahdi, S.S.; Claes, P.; Steyaert, J.; Boets, B. Rapid neural categorization of angry and fearful faces is specifically impaired in boys with autism spectrum disorder. J. Child Psychol. Psychiatry 2020, 61, 1019–1029. [Google Scholar] [CrossRef]
- Vettori, S.; Dzhelyova, M.; Van der Donck, S.; Jacques, C.; Steyaert, J.; Rossion, B.; Boets, B. Reduced neural sensitivity to rapid individual face discrimination in autism spectrum disorder. NeuroImage: Clin. 2018, 21, 101613. [Google Scholar] [CrossRef]
- Leleu, A.; Favre, E.; Yailian, A.; Fumat, H.; Klamm, J.; Amado, I.; Baudouin, J.-Y.; Franck, N.; Demily, C. An implicit and reliable neural measure quantifying impaired visual coding of facial expression: Evidence from the 22q11.2 deletion syndrome. Transl. Psychiatry 2019, 9, 67. [Google Scholar] [CrossRef] [Green Version]
- Poncet, F.; Baudouin, J.-Y.; Dzhelyova, M.P.; Rossion, B.; Leleu, A. Rapid and automatic discrimination between facial expressions in the human brain. Neuropsychologia 2019, 129, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Van der Donck, S.; Moerkerke, M.; Dlhosova, T.; Vettori, S.; Dzhelyova, M.; Alaerts, K.; Boets, B. Monitoring the effect of oxytocin on the neural sensitivity to emotional faces via frequency-tagging EEG: A double-blind, cross-over study. Psychophysiol. 2022, 59, e14026. [Google Scholar] [CrossRef] [PubMed]
- Grill-Spector, K.; Weiner, K.S.; Kay, K.; Gomez, J. The Functional Neuroanatomy of Human Face Perception. Annu. Rev. Vis. Sci. 2017, 3, 167–196. [Google Scholar] [CrossRef] [PubMed]
- Haxby, J.V.; Hoffman, E.A.; Gobbini, M.I. The distributed human neural system for face perception. Trends Cogn. Sci. 2000, 4, 223–233. [Google Scholar] [CrossRef]
- Kanwisher, N.; McDermott, J.; Chun, M. The Fusiform Face Area: A Module in Human Extrastriate Cortex Specialized for Face Perception. J. Neurosci. 1997, 17, 4302–4311. [Google Scholar] [CrossRef]
- Puce, A.; Allison, T.; Gore, J.C.; McCarthy, G. Face-sensitive regions in human extrastriate cortex studied by functional MRI. J. Neurophysiol. 1995, 74, 1192–1199. [Google Scholar] [CrossRef]
- Epstein, R.; Kanwisher, N. A cortical representation of the local visual environment. Nature 1998, 392, 598–601. [Google Scholar] [CrossRef]
- Kadipasaoglu, C.M.; Conner, C.R.; Whaley, M.L.; Baboyan, V.G.; Tandon, N. Category-Selectivity in Human Visual Cortex Follows Cortical Topology: A Grouped icEEG Study. PLoS ONE 2016, 11, e0157109. [Google Scholar] [CrossRef]
- Weiner, K.S.; Grill-Spector, K. Sparsely-distributed organization of face and limb activations in human ventral temporal cortex. NeuroImage 2010, 52, 1559–1573. [Google Scholar] [CrossRef] [PubMed]
- Jacques, C.; Retter, T.L.; Rossion, B. A single glance at natural face images generate larger and qualitatively different category-selective spatio-temporal signatures than other ecologically-relevant categories in the human brain. NeuroImage 2016, 137, 21–33. [Google Scholar] [CrossRef]
- Domes, G.; Lischke, A.; Berger, C.; Grossmann, A.; Hauenstein, K.; Heinrichs, M.; Herpertz, S.C. Effects of intranasal oxytocin on emotional face processing in women. Psychoneuroendocrinology 2010, 35, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, K.S. Sex, Receptors, and Attachment: A Review of Individual Factors Influencing Response to Oxytocin. Front. Behav. Neurosci. 2013, 6, 194. [Google Scholar] [CrossRef]
- Graustella, A.J.; MacLeod, C. A critical review of the influence of oxytocin nasal spray on social cognition in humans: Evidence and future directions. Horm. Behav. 2012, 61, 410–418. [Google Scholar] [CrossRef]
- Quintana, D.S.; Lischke, A.; Grace, S.; Scheele, D.; Ma, Y.; Becker, B. Advances in the field of intranasal oxytocin research: Lessons learned and future directions for clinical research. Mol. Psychiatry 2020, 26, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Daughters, K.; Manstead, A.; Hubble, K.; Rees, A.; Thapar, A.; Van Goozen, S.H.M. Salivary Oxytocin Concentrations in Males following Intranasal Administration of Oxytocin: A Double-Blind, Cross-Over Study. PLoS ONE 2015, 10, e0145104. [Google Scholar] [CrossRef]
- Striepens, N.; Kendrick, K.M.; Hanking, V.; Landgraf, R.; Wüllner, U.; Maier, W.; Hurlemann, R. Elevated cerebrospinal fluid and blood concentrations of oxytocin following its intranasal administration in humans. Sci. Rep. 2013, 3, 3440. [Google Scholar] [CrossRef] [PubMed]
- Retter, T.L.; Rossion, B. Uncovering the neural magnitude and spatio-temporal dynamics of natural image categorization in a fast visual stream. Neuropsychologia 2016, 91, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Makeig, S.; Bell, A.J.; Jung, T.-P.; Sejnowski, T.J. Independent Component Analysis of Electroencephalographic Data. In Advances in Neural Information Processing Systems 8; Touretzky, D.S., Mozer, M.C., Hasselmo, M.E., Eds.; MIT Press: Cambridge, United Kingdom, 1996; pp. 145–151. [Google Scholar]
- Regan, D. Human Brain Electrophysiology: Evoked Potentials and Evoked Magnetic Fields in Science and Medicine; Elsevier: Amsterdam, The Netherlands, 1989. [Google Scholar]
- Rossion, B.; Torfs, K.; Jacques, C.; Liu-Shuang, J. Fast periodic presentation of natural images reveals a robust face-selective electrophysiological response in the human brain. J. Vis. 2015, 15, 18. [Google Scholar] [CrossRef]
- Dzhelyova, M.; Jacques, C.; Rossion, B. At a Single Glance: Fast Periodic Visual Stimulation Uncovers the Spatio-Temporal Dynamics of Brief Facial Expression Changes in the Human Brain. Cereb. Cortex 2016, 27, 4106–4123. [Google Scholar] [CrossRef] [PubMed]
- Liu-Shuang, J.; Norcia, A.M.; Rossion, B. An objective index of individual face discrimination in the right occipito-temporal cortex by means of fast periodic oddball stimulation. Neuropsychologia 2013, 52, 57–72. [Google Scholar] [CrossRef]
- Singmann, H.; Bolker, B.; Westfall, J.; Aust, F.; Ben- Shachar, M.S. afex: Analysis of Factorial Experiments. R Package Version 1.1-0. 2022. Available online: https://CRAN.R-project.org/package=afex (accessed on 29 July 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 29 July 2021).
- Lenth, R. emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.7.3. 2022. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 29 July 2021).
- Morey, R.D. and Rouder, J.N. BayesFactor: Computation of Bayes Factors for Common Designs. R Package Version 0.9.12-4.3. 2021. Available online: https://CRAN.R-project.org/package=BayesFactor (accessed on 29 July 2021).
- Lee, M.D.; Wagenmakers, E.-J. Bayesian Model Comparison. In Bayesian Cognitive Modeling: A Practical Course; Cambridge University Press: Cambridge, UK, 2014; pp. 101–117. [Google Scholar] [CrossRef]
- Jeffreys, H. Theory of Probabilit, 3rd ed.; Oxford University Press: Oxford, UK, 1961. [Google Scholar]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Hagen, S.; Jacques, C.; Maillard, L.; Colnat-Coulbois, S.; Rossion, B.; Jonas, J. Spatially Dissociated Intracerebral Maps for Face- and House-Selective Activity in the Human Ventral Occipito-Temporal Cortex. Cereb. Cortex 2020, 30, 4026–4043. [Google Scholar] [CrossRef]
- Epstein, R.A.; Kveraga, K.; Bar, M. Neural Systems for Visual Scene Recognition. In Scene Vision: Making Sense of What We See; MIT Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Jacques, C.; Witthoft, N.; Weiner, K.S.; Foster, B.L.; Rangarajan, V.; Hermes, D.; Miller, K.J.; Parvizi, J.; Grill-Spector, K. Corresponding ECoG and fMRI category-selective signals in human ventral temporal cortex. Neuropsychologia 2015, 83, 14–28. [Google Scholar] [CrossRef]
- Dzhelyova, M.; Jacques, C.; Dormal, G.; Michel, C.; Schiltz, C.; Rossion, B. High test-retest reliability of a neural index of rapid automatic discrimination of unfamiliar individual faces. Vis. Cogn. 2019, 27, 127–141. [Google Scholar] [CrossRef]
- Jeste, S.S.; Frohlich, J.; Loo, S.K. Electrophysiological biomarkers of diagnosis and outcome in neurodevelopmental disorders. Curr. Opin. Neurol. 2015, 28, 110–116. [Google Scholar] [CrossRef]
- Winterton, A.; Westlye, L.T.; Steen, N.E.; Andreassen, O.A.; Quintana, D.S. Improving the precision of intranasal oxytocin research. Nat. Hum. Behav. 2020, 5, 9–18. [Google Scholar] [CrossRef]
- Tabak, B.A.; Teed, A.R.; Castle, E.; Dutcher, J.M.; Meyer, M.L.; Bryan, R.; Irwin, M.R.; Lieberman, M.D.; Eisenberger, N.I. Null results of oxytocin and vasopressin administration across a range of social cognitive and behavioral paradigms: Evidence from a randomized controlled trial. Psychoneuroendocrinology 2019, 107, 124–132. [Google Scholar] [CrossRef]
- Bartz, J.A.; Zaki, J.; Bolger, N.; Hollander, E.; Ludwig, N.; Kolevzon, A.; Ochsner, K.N. Oxytocin Selectively Improves Empathic Accuracy. Psychol. Sci. 2010, 21, 1426–1428. [Google Scholar] [CrossRef]
- Feeser, M.; Fan, Y.; Weigand, A.; Hahn, A.; Gärtner, M.; Böker, H.; Grimm, S.; Bajbouj, M. Oxytocin improves mentalizing–Pronounced effects for individuals with attenuated ability to empathize. Psychoneuroendocrinology 2015, 53, 223–232. [Google Scholar] [CrossRef]
- Harari-Dahan, O.; Bernstein, A. A general approach-avoidance hypothesis of Oxytocin: Accounting for social and non-social effects of oxytocin. Neurosci. Biobehav. Rev. 2014, 47, 506–519. [Google Scholar] [CrossRef]
- Harari-Dahan, O.; Bernstein, A. Oxytocin attenuates social and non-social avoidance: Re-thinking the social specificity of Oxytocin. Psychoneuroendocrinology 2017, 81, 105–112. [Google Scholar] [CrossRef]
- Alaerts, K.; Taillieu, A.; Daniels, N.; Soriano, J.R.; Prinsen, J. Oxytocin enhances neural approach towards social and non-social stimuli of high personal relevance. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Trilla, I.; Drimalla, H.; Bajbouj, M.; Dziobek, I. The Influence of Reward on Facial Mimicry: No Evidence for a Significant Effect of Oxytocin. Front. Behav. Neurosci. 2020, 14, 88. [Google Scholar] [CrossRef]
- Melkonyan, A.; Liu, L.; Brown, E.C.; Meyer, W.; Madipakkam, A.R.; Ringelmann, L.; Lange, F.; Schmid, S.M.; Münte, T.F.; Park, S.Q. Unchanged food approach-avoidance behaviour of healthy men after oxytocin administration. J. Neuroendocr. 2020, 32, e12923. [Google Scholar] [CrossRef]
- Lane, A.; Luminet, O.; Nave, G.; Mikolajczak, M. Is there a Publication Bias in Behavioural Intranasal Oxytocin Research on Humans? Opening the File Drawer of One Laboratory. J. Neuroendocr. 2016, 28, 1–15. [Google Scholar] [CrossRef]
- Bürkner, P.-C.; Williams, D.R.; Simmons, T.C.; Woolley, J.D. Intranasal Oxytocin May Improve High-Level Social Cognition in Schizophrenia, But Not Social Cognition or Neurocognition in General: A Multilevel Bayesian Meta-analysis. Schizophr. Bull. 2017, 43, 1291–1303. [Google Scholar] [CrossRef]
- Huang, M.; Liu, K.; Wei, Z.; Feng, Z.; Chen, J.; Yang, J.; Zhong, Q.; Wan, G.; Kong, X.-J. Serum Oxytocin Level Correlates With Gut Microbiome Dysbiosis in Children With Autism Spectrum Disorder. Front. Neurosci. 2021, 15, 1320. [Google Scholar] [CrossRef]
- Peled-Avron, L.; Abu-Akel, A.; Shamay-Tsoory, S. Exogenous effects of oxytocin in five psychiatric disorders: A systematic review, meta-analyses and a personalized approach through the lens of the social salience hypothesis. Neurosci. Biobehav. Rev. 2020, 114, 70–95. [Google Scholar] [CrossRef]
- Ma, Y.; Shamay-Tsoory, S.; Han, S.; Zink, C.F. Oxytocin and Social Adaptation: Insights from Neuroimaging Studies of Healthy and Clinical Populations. Trends Cogn. Sci. 2016, 20, 133–145. [Google Scholar] [CrossRef]
- Walum, H.; Waldman, I.D.; Young, L.J. Statistical and Methodological Considerations for the Interpretation of Intranasal Oxytocin Studies. Biol. Psychiatry 2015, 79, 251–257. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Effect | F Value | p Value | Post-Hoc Test | |
|---|---|---|---|---|
| Treatment condition | F(1, 31) = 0.03 | p = 0.856 | / | |
| Stimulus type | F(1, 30) = 0.16 | p = 0.696 | / | |
| ROI | F(2, 40) = 48.03 | p < 0.001 | MO (2.20 µV) > LOT (1.22 µV), p < 0.001 | |
| MO (2.20 µV) > ROT (1.61 µV), p < 0.001 | ||||
| ROT (1.61 µV) > LOT (1.22 µV), p = 0.012 | ||||
| Session order | F(1, 29) = 0.46 | p = 0.501 | / | |
| Treatment condition and stimulus type | F(1, 551) = 0.08 | p = 0.775 | / | |
| Treatment condition and ROI | F(2, 552) = 0.28 | p = 0.759 | / | |
| Stimulus type and ROI | F(2, 557) = 184.26 | p < 0.001 | face | ROT > LOT, p = 0.009 |
| MO > LOT, p = 0.006 | ||||
| house | MO > LOT, p < 0.001 | |||
| MO > ROT, p < 0.001 | ||||
| in LOT in ROT | face > house, p = 0.022 | |||
| face > house, p < 0.001 | ||||
| in MO | house > face, p < 0.001 | |||
| Treatment condition, stimulus type and ROI | F(2, 553) = 0.24 | p = 0.788 | / | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, Z.; Van der Donck, S.; Moerkerke, M.; Dlhosova, T.; Vettori, S.; Dzhelyova, M.; van Winkel, R.; Alaerts, K.; Boets, B. Frequency-Tagging EEG of Superimposed Social and Non-Social Visual Stimulation Streams Provides No Support for Social Salience Enhancement after Intranasal Oxytocin Administration. Brain Sci. 2022, 12, 1224. https://doi.org/10.3390/brainsci12091224
Qiao Z, Van der Donck S, Moerkerke M, Dlhosova T, Vettori S, Dzhelyova M, van Winkel R, Alaerts K, Boets B. Frequency-Tagging EEG of Superimposed Social and Non-Social Visual Stimulation Streams Provides No Support for Social Salience Enhancement after Intranasal Oxytocin Administration. Brain Sciences. 2022; 12(9):1224. https://doi.org/10.3390/brainsci12091224
Chicago/Turabian StyleQiao, Zhiling, Stephanie Van der Donck, Matthijs Moerkerke, Tereza Dlhosova, Sofie Vettori, Milena Dzhelyova, Ruud van Winkel, Kaat Alaerts, and Bart Boets. 2022. "Frequency-Tagging EEG of Superimposed Social and Non-Social Visual Stimulation Streams Provides No Support for Social Salience Enhancement after Intranasal Oxytocin Administration" Brain Sciences 12, no. 9: 1224. https://doi.org/10.3390/brainsci12091224
APA StyleQiao, Z., Van der Donck, S., Moerkerke, M., Dlhosova, T., Vettori, S., Dzhelyova, M., van Winkel, R., Alaerts, K., & Boets, B. (2022). Frequency-Tagging EEG of Superimposed Social and Non-Social Visual Stimulation Streams Provides No Support for Social Salience Enhancement after Intranasal Oxytocin Administration. Brain Sciences, 12(9), 1224. https://doi.org/10.3390/brainsci12091224