
Acute Vagus Nerve Stimulation Facilitates Short Term Memory and Cognitive Flexibility in Rats



Abstract
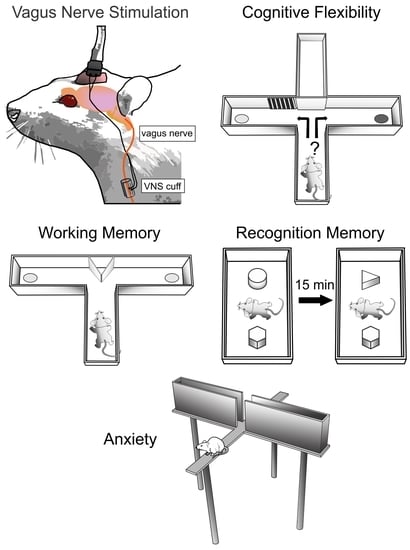
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Surgical Procedures
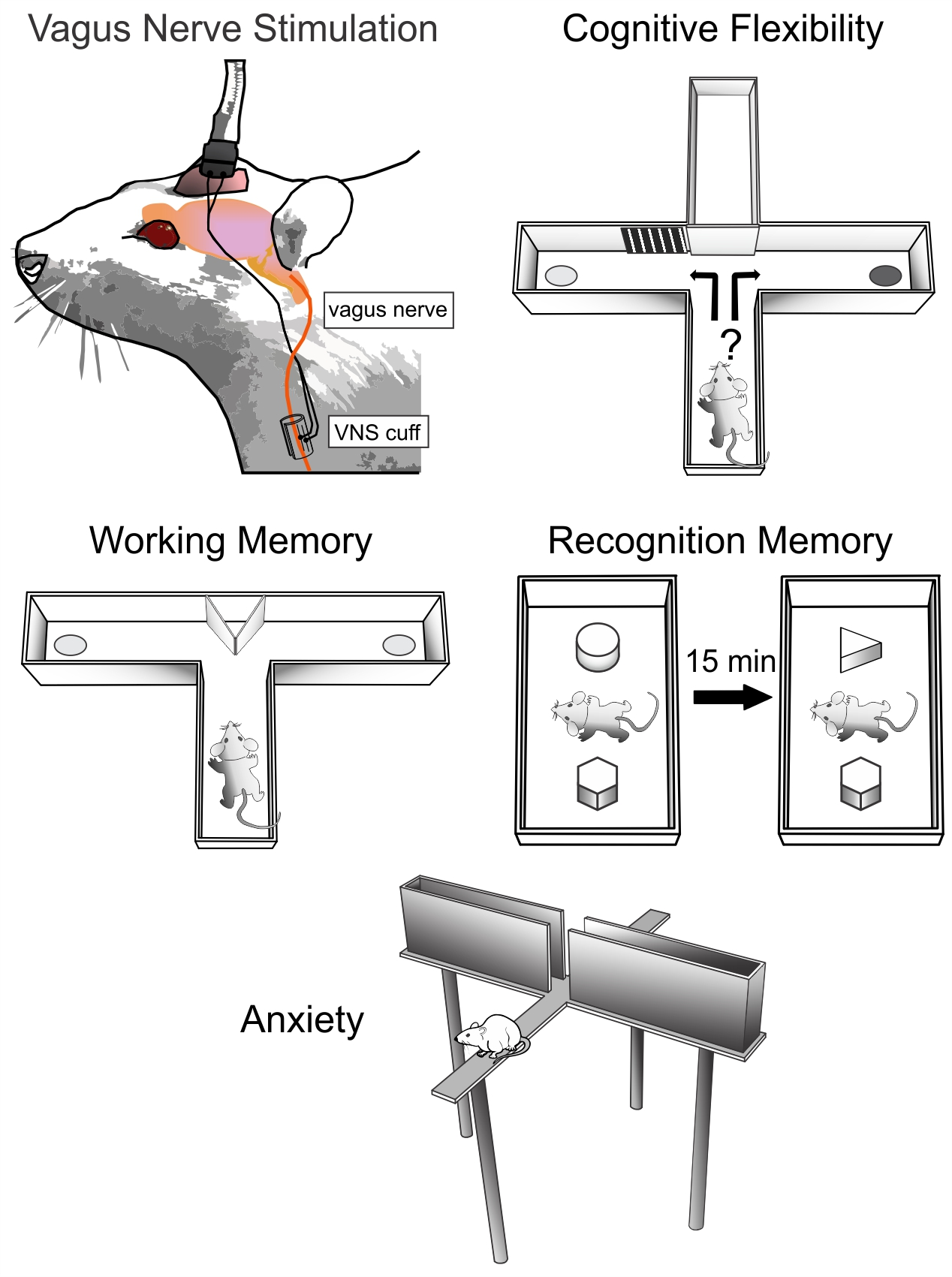
2.2. Cross-Maze Rule-Shifting Task (CMRST)
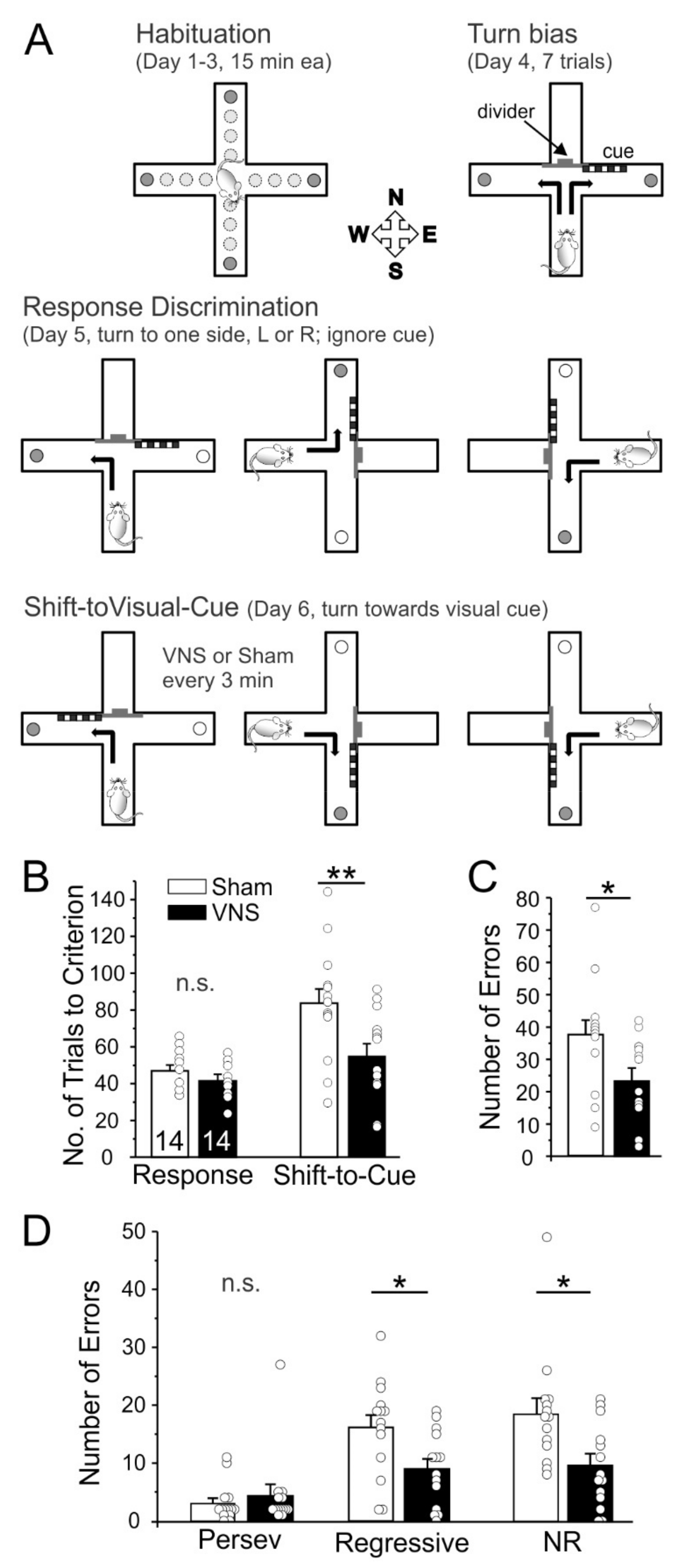
2.3. Delayed Spontaneous Alternation
2.4. Novel Object Recognition
2.5. Elevated plus Maze
2.6. Data Analysis
3. Results
3.1. Cross-Maze Rule Shifting Task
3.2. Delayed Spontaneous Alternation
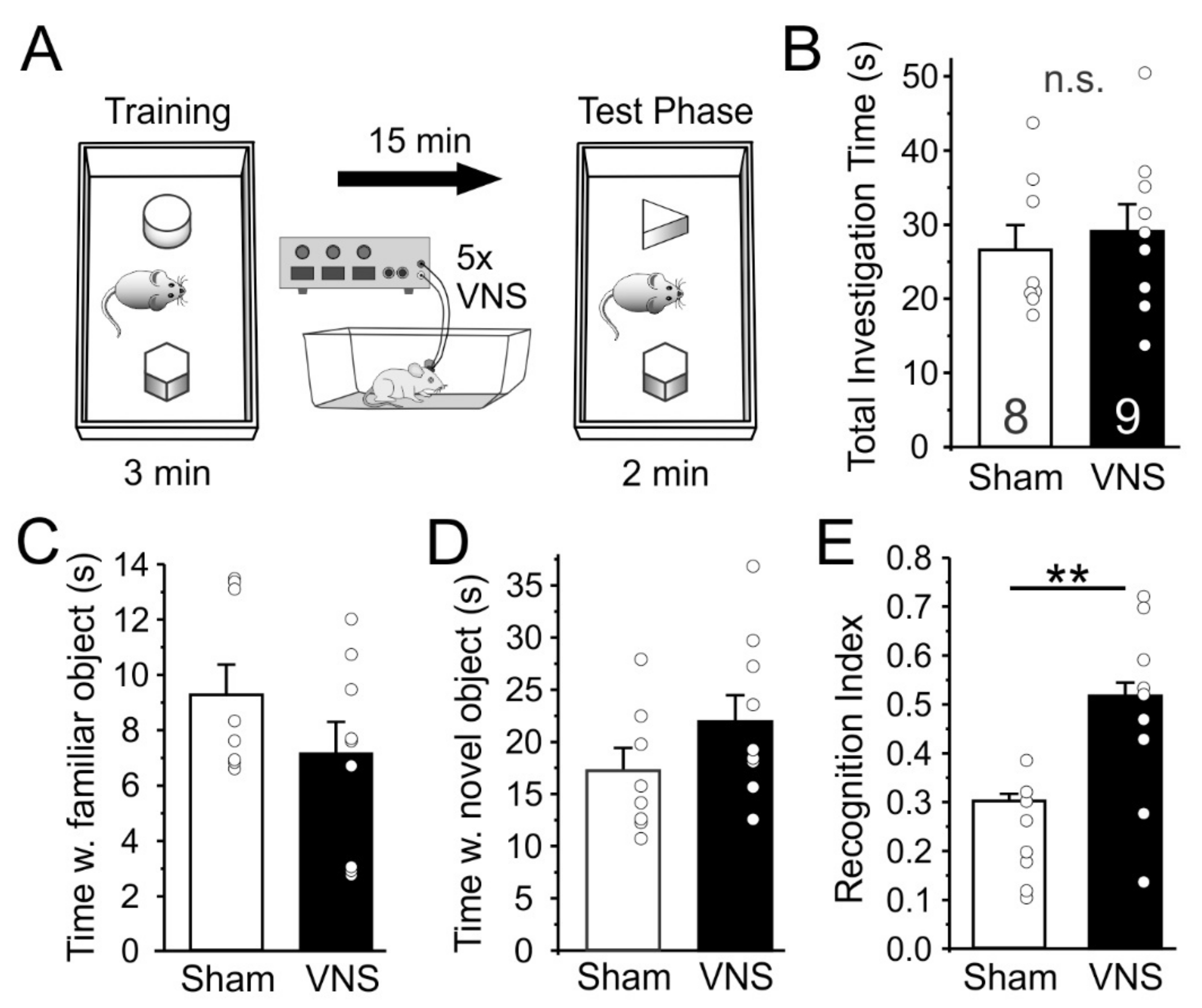
3.3. Novel Object Recognition
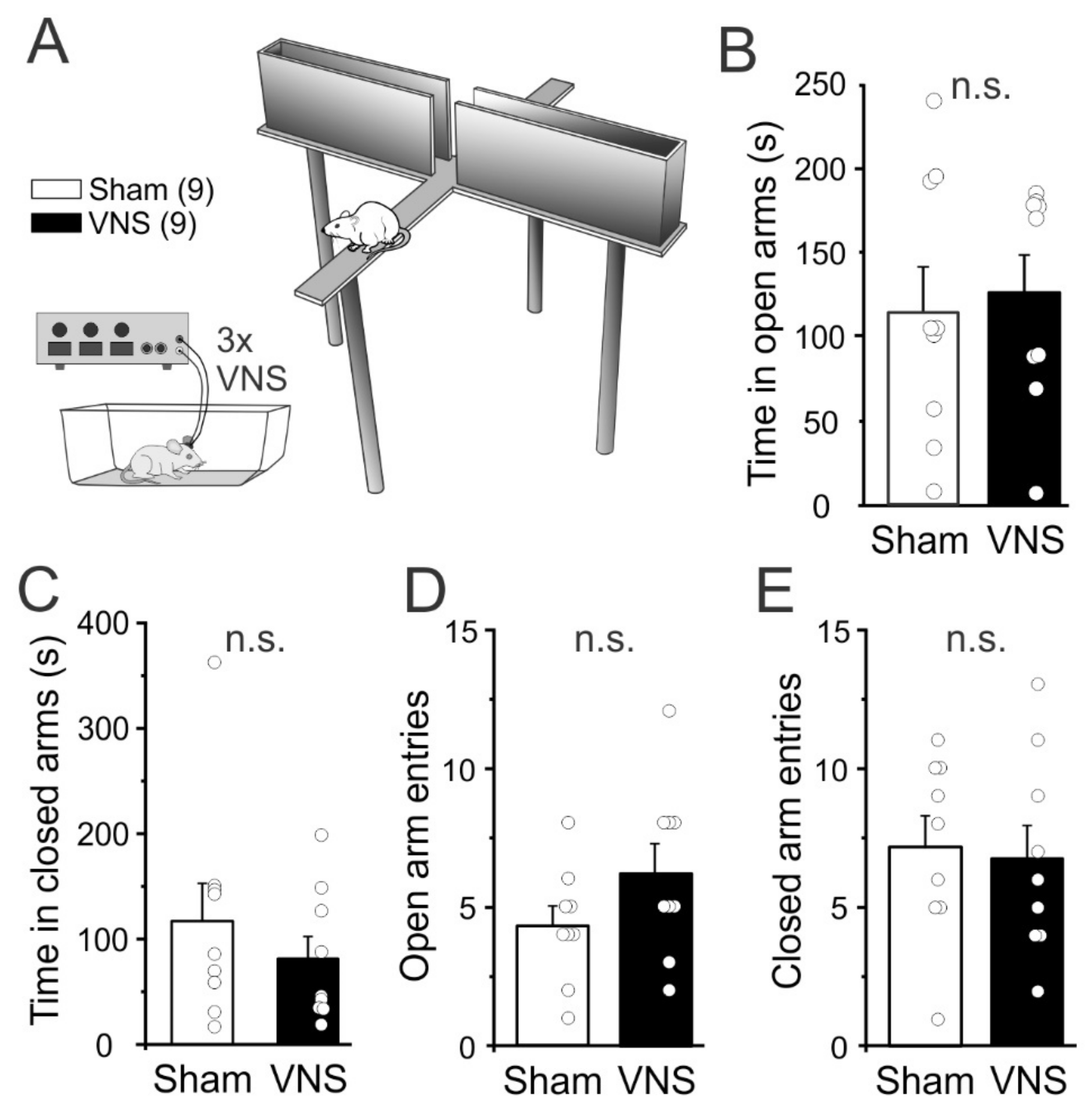
3.4. Elevated Plus Maze
4. Discussion
4.1. Rule Shifting
4.2. Short-Term or Working Memory
4.3. Novel Object Recognition
4.4. Elevated plus Maze
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ondicova, K.; Pecenák, J.; Mravec, B. The role of the vagus nerve in depression. Neuro Endocrinol. Lett. 2010, 31, 602–608. [Google Scholar] [PubMed]
- Marangell, L.B.; Martinez, M.; Jurdi, R.A.; Zboyan, H. Neurostimulation therapies in depression: A review of new modalities. Acta Psychiatr. Scand. 2007, 116, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.E.; Morsi, A.; Kalhorn, S.P.; Marcus, J.; Sellin, J.; Kang, M.; Silverberg, A.; Rivera, E.; Geller, E.; Carlson, C.; et al. Vagus nerve stimulation in 436 consecutive patients with treatment-resistant epilepsy: Long-term outcomes and predictors of response. Epilepsy Behav. 2011, 20, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Colzato, L.S.; Jongkees, B.; De Wit, M.; Van Der Molen, M.J.W.; Steenbergen, L. Variable heart rate and a flexible mind: Higher resting-state heart rate variability predicts better task-switching. Cogn. Affect. Behav. Neurosci. 2018, 18, 730–738. [Google Scholar] [CrossRef]
- Vonck, K.; Raedt, R.; Naulaerts, J.; De Vogelaere, F.; Thiery, E.; Van Roost, D.; Aldenkamp, B.; Miatton, M.; Boon, P. Vagus nerve stimulation…25 years later! What do we know about the effects on cognition? Neurosci. Biobehav. Rev. 2014, 45, 63–71. [Google Scholar] [CrossRef]
- Ricardo, J.A.; Koh, E.T. Anatomical evidence of direct projections from the nucleus of the solitary tract to the hypothalamus, amygdala, and other forebrain structures in the rat. Brain Res. 1978, 153, 1–26. [Google Scholar] [CrossRef]
- Hassert, D.L.; Miyashita, T.; Williams, C.L. The Effects of Peripheral Vagal Nerve Stimulation at a Memory-Modulating Intensity on Norepinephrine Output in the Basolateral Amygdala. Behav. Neurosci. 2004, 118, 79–88. [Google Scholar] [CrossRef]
- Dorr, A.E.; Debonnel, G. Effect of Vagus Nerve Stimulation on Serotonergic and Noradrenergic Transmission. J. Pharmacol. Exp. Ther. 2006, 318, 890–898. [Google Scholar] [CrossRef]
- Manta, S.; Dong, J.; Debonnel, G.; Blier, P. Enhancement of the function of rat serotonin and norepinephrine neurons by sustained vagus nerve stimulation. J. Psychiatry Neurosci. 2009, 34, 272–280. [Google Scholar]
- Manta, S.; El Mansari, M.; Debonnel, G.; Blier, P. Electrophysiological and neurochemical effects of long-term vagus nerve stimulation on the rat monoaminergic systems. Int. J. Neuropsychopharmacol. 2013, 16, 459–470. [Google Scholar] [CrossRef]
- Roosevelt, R.W.; Smith, D.C.; Clough, R.W.; Jensen, R.A.; Browning, R.A. Increased extracellular concentrations of norepinephrine in cortex and hippocampus following vagus nerve stimulation in the rat. Brain Res. 2006, 1119, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.C.; Jo, B.G.; Lee, C.-Y.; Lee, K.-W.; Namgung, U. Hippocampal activation of 5-HT1B receptors and BDNF production by vagus nerve stimulation in rats under chronic restraint stress. Eur. J. Neurosci. 2019, 50, 1820–1830. [Google Scholar] [CrossRef] [PubMed]
- Furmaga, H.; Shah, A.; Frazer, A. Serotonergic and Noradrenergic Pathways Are Required for the Anxiolytic-like and Antidepressant-like Behavioral Effects of Repeated Vagal Nerve Stimulation in Rats. Biol. Psychiatry 2011, 70, 937–945. [Google Scholar] [CrossRef]
- Nichols, J.; Nichols, A.; Smirnakis, S.; Engineer, N.; Kilgard, M.; Atzori, M. Vagus nerve stimulation modulates cortical synchrony and excitability through the activation of muscarinic receptors. Neuroscience 2011, 189, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Chase, M.; Sterman, M.; Clemente, C. Cortical and subcortical patterns of response to afferent vagal stimulation. Exp. Neurol. 1966, 16, 36–49. [Google Scholar] [CrossRef]
- Krahl, S.E.; Clark, K.B. Vagus nerve stimulation for epilepsy: A review of central mechanisms. Surg. Neurol. Int. 2012, 3, 255–259. [Google Scholar] [CrossRef]
- Cao, B.; Wang, J.; Shahed, M.; Jelfs, B.; Chan, R.H.M.; Li, Y. Vagus Nerve Stimulation Alters Phase Synchrony of the Anterior Cingulate Cortex and Facilitates Decision Making in Rats. Sci. Rep. 2016, 6, 35135. [Google Scholar] [CrossRef]
- Cao, J.; Lu, K.-H.; Powley, T.L.; Liu, Z. Vagal nerve stimulation triggers widespread responses and alters large-scale functional connectivity in the rat brain. PLoS ONE 2017, 12, e0189518. [Google Scholar] [CrossRef]
- Collins, L.; Boddington, L.; Steffan, P.J.; McCormick, D. Vagus nerve stimulation induces widespread cortical and behavioral activation. Curr. Biol. 2021, 31, 2088–2098.e3. [Google Scholar] [CrossRef]
- Clark, K.; Krahl, S.; Smith, D.; Jensen, R. Post-training Unilateral Vagal Stimulation Enhances Retention Performance in the Rat. Neurobiol. Learn. Mem. 1995, 63, 213–216. [Google Scholar] [CrossRef]
- Clark, K.; Smith, D.; Hassert, D.; Browning, R.; Naritoku, D.; Jensen, R. Posttraining Electrical Stimulation of Vagal Afferents with Concomitant Vagal Efferent Inactivation Enhances Memory Storage Processes in the Rat. Neurobiol. Learn. Mem. 1998, 70, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Sanders, T.H.; Weiss, J.; Hogewood, L.; Chen, L.; Paton, C.; Mcmahan, R.L.; Sweatt, J.D. Cognition-Enhancing Vagus Nerve Stimulation Alters the Epigenetic Landscape. J. Neurosci. 2019, 39, 3454–3469. [Google Scholar] [CrossRef]
- Childs, J.E.; DeLeon, J.; Nickel, E.; Kroener, S. Vagus nerve stimulation reduces cocaine seeking and alters plasticity in the extinction network. Learn. Mem. 2017, 24, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Peña, D.F.; Engineer, N.D.; McIntyre, C.K. Rapid Remission of Conditioned Fear Expression with Extinction Training Paired with Vagus Nerve Stimulation. Biol. Psychiatry 2013, 73, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Engineer, N.D.; Riley, J.; Seale, J.D.; Vrana, W.A.; Shetake, J.A.; Sudanagunta, S.P.; Borland, M.S.; Kilgard, M. Reversing pathological neural activity using targeted plasticity. Nature 2011, 470, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.B.; Naritoku, D.K.; Smith, D.C.; Browning, R.A.; Jensen, R.A. Enhanced recognition memory following vagus nerve stimulation in human subjects. Nat. Neurosci. 1999, 2, 94–98. [Google Scholar] [CrossRef]
- Sun, L.; Peräkylä, J.; Holm, K.; Haapasalo, J.; Lehtimäki, K.; Ogawa, K.H.; Peltola, J.; Hartikainen, K.M. Vagus nerve stimulation improves working memory performance. J. Clin. Exp. Neuropsychol. 2017, 39, 954–964. [Google Scholar] [CrossRef]
- Altidor, L.K.-P.; Bruner, M.M.; Deslauriers, J.F.; Garman, T.S.; Ramirez, S.; Dirr, E.W.; Olczak, K.P.; Maurer, A.P.; Lamb, D.G.; Otto, K.J.; et al. Acute vagus nerve stimulation enhances reversal learning in rats. Neurobiol. Learn. Mem. 2021, 184, 107498. [Google Scholar] [CrossRef]
- Childs, J.E.; Alvarez-Dieppa, A.C.; McIntyre, C.K.; Kroener, S. Vagus Nerve Stimulation as a Tool to Induce Plasticity in Pathways Relevant for Extinction Learning. J. Vis. Exp. 2015, 102, e53032. [Google Scholar] [CrossRef]
- Childs, J.E.; Kim, S.; Driskill, C.M.; Hsiu, E.; Kroener, S. Vagus nerve stimulation during extinction learning reduces conditioned place preference and context-induced reinstatement of cocaine seeking. Brain Stimul. 2019, 12, 1448–1455. [Google Scholar] [CrossRef]
- Butler, A.G.; O’Callaghan, E.L.; Allen, A.M.; McDougall, S.J. Use of a physiological reflex to standardize vagal nerve stimulation intensity improves data reproducibility in a memory extinction assay. Brain Stimul. 2021, 14, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Jeevakumar, V.; Driskill, C.; Paine, A.; Sobhanian, M.; Vakil, H.; Morris, B.; Ramos, J.; Kroener, S. Ketamine administration during the second postnatal week induces enduring schizophrenia-like behavioral symptoms and reduces parvalbumin expression in the medial prefrontal cortex of adult mice. Behav. Brain Res. 2015, 282, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Phensy, A.; Duzdabanian, H.E.; Brewer, S.; Panjabi, A.; Driskill, C.; Berz, A.; Peng, G.; Kroener, S. Antioxidant Treatment with N-acetyl Cysteine Prevents the Development of Cognitive and Social Behavioral Deficits that Result from Perinatal Ketamine Treatment. Front. Behav. Neurosci. 2017, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef]
- George, M.S.; Ward, H.E.; Ninan, P.; Pollack, M.; Nahas, Z.; Anderson, B.; Kose, S.; Howland, R.H.; Goodman, W.K.; Ballenger, J.C. A pilot study of vagus nerve stimulation (VNS) for treatment-resistant anxiety disorders. Brain Stimul. 2008, 1, 112–121. [Google Scholar] [CrossRef]
- Noble, L.J.; Chuah, A.; Callahan, K.K.; Souza, R.R.; McIntyre, C.K. Peripheral effects of vagus nerve stimulation on anxiety and extinction of conditioned fear in rats. Learn. Mem. 2019, 26, 245–251. [Google Scholar] [CrossRef]
- Noble, L.J.; Meruva, V.B.; Hays, S.A.; Rennaker, R.L.; Kilgard, M.; McIntyre, C.K. Vagus nerve stimulation promotes generalization of conditioned fear extinction and reduces anxiety in rats. Brain Stimul. 2019, 12, 9–18. [Google Scholar] [CrossRef]
- Porter, B.A.; Khodaparast, N.; Fayyaz, T.; Cheung, R.J.; Ahmed, S.S.; Vrana, W.A.; Rennaker, I.R.L.; Kilgard, M. Repeatedly Pairing Vagus Nerve Stimulation with a Movement Reorganizes Primary Motor Cortex. Cereb. Cortex 2012, 22, 2365–2374. [Google Scholar] [CrossRef]
- Khodaparast, N.; Hays, S.A.; Sloan, A.M.; Hulsey, D.R.; Ruiz, A.; Pantoja, M.; Rennaker, R.L.; Kilgard, M.P. Vagus nerve stimulation during rehabilitative training improves forelimb strength following ischemic stroke. Neurobiol. Dis. 2013, 60, 80–88. [Google Scholar] [CrossRef]
- Bowles, S.; Hickman, J.; Peng, X.; Williamson, W.R.; Huang, R.; Washington, K.; Donegan, D.; Welle, C.G. Vagus nerve stimulation drives selective circuit modulation through cholinergic reinforcement. Neuron 2022, 110, 1–19. [Google Scholar] [CrossRef]
- Penã, D.F.; Childs, J.E.; Willett, S.; Vital, A.; McIntyre, C.K.; Kroener, S.; Peña, D.F. Vagus nerve stimulation enhances extinction of conditioned fear and modulates plasticity in the pathway from the ventromedial prefrontal cortex to the amygdala. Front. Behav. Neurosci. 2014, 8, 327. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, L.; Sellaro, R.; Stock, A.-K.; Verkuil, B.; Beste, C.; Colzato, L.S. RETRACTED: Transcutaneous vagus nerve stimulation (tVNS) enhances response selection during action cascading processes. Eur. Neuropsychopharmacol. 2015, 25, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Floresco, S.B.; Magyar, O.; Ghods-Sharifi, S.; Vexelman, C.; Tse, M.T.L. Multiple Dopamine Receptor Subtypes in the Medial Prefrontal Cortex of the Rat Regulate Set-Shifting. Neuropsychopharmacology 2006, 31, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Young, J.W.; Powell, S.B.; Risbrough, V.; Marston, H.M.; Geyer, M.A. Using the MATRICS to guide development of a preclinical cognitive test battery for research in schizophrenia. Pharmacol. Ther. 2009, 122, 150–202. [Google Scholar] [CrossRef]
- Han, W.; Tellez, L.A.; Perkins, M.H.; Perez, I.O.; Qu, T.; Ferreira, J.; Ferreira, T.L.; Quinn, D.; Liu, Z.-W.; Gao, X.-B.; et al. A Neural Circuit for Gut-Induced Reward. Cell 2018, 175, 887–888. [Google Scholar] [CrossRef]
- Dehais, F.; Hodgetts, H.M.; Causse, M.; Behrend, J.; Durantin, G.; Tremblay, S. Momentary lapse of control: A cognitive continuum approach to understanding and mitigating perseveration in human error. Neurosci. Biobehav. Rev. 2019, 100, 252–262. [Google Scholar] [CrossRef]
- Fuster, J.M. The Prefrontal Cortex, 5th ed.; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Floresco, S.B.; Zhang, Y.; Enomoto, T. Neural circuits subserving behavioral flexibility and their relevance to schizophrenia. Behav. Brain Res. 2009, 204, 396–409. [Google Scholar] [CrossRef]
- Birrell, J.M.; Brown, V.J. Medial Frontal Cortex Mediates Perceptual Attentional Set Shifting in the Rat. J. Neurosci. 2000, 20, 4320–4324. [Google Scholar] [CrossRef]
- Bissonette, G.B.; Powell, E.M.; Roesch, M.R. Neural structures underlying set-shifting: Roles of medial prefrontal cortex and anterior cingulate cortex. Behav. Brain Res. 2013, 250, 91–101. [Google Scholar] [CrossRef]
- Ragozzino, M.E.; Detrick, S.; Kesner, R.P. Involvement of the Prelimbic–Infralimbic Areas of the Rodent Prefrontal Cortex in Behavioral Flexibility for Place and Response Learning. J. Neurosci. 1999, 19, 4585–4594. [Google Scholar] [CrossRef]
- Borodovitsyna, O.; Flamini, M.; Chandler, D. Noradrenergic Modulation of Cognition in Health and Disease. Neural Plast. 2017, 2017, 6031478. [Google Scholar] [CrossRef] [PubMed]
- Robbins, T.; Arnsten, A. The Neuropsychopharmacology of Fronto-Executive Function: Monoaminergic Modulation. Annu. Rev. Neurosci. 2009, 32, 267–287. [Google Scholar] [CrossRef] [PubMed]
- Thayer, J.F.; Hansen, A.L.; Saus-Rose, E.; Johnsen, B.H. Heart Rate Variability, Prefrontal Neural Function, and Cognitive Performance: The Neurovisceral Integration Perspective on Self-regulation, Adaptation, and Health. Ann. Behav. Med. 2009, 37, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Colzato, L.; Beste, C. A literature review on the neurophysiological underpinnings and cognitive effects of transcutaneous vagus nerve stimulation: Challenges and future directions. J. Neurophysiol. 2020, 123, 1739–1755. [Google Scholar] [CrossRef]
- Klarer, M.; Weber-Stadlbauer, U.; Arnold, M.; Langhans, W.; Meyer, U. Cognitive effects of subdiaphragmatic vagal deafferentation in rats. Neurobiol. Learn. Mem. 2017, 142, 190–199. [Google Scholar] [CrossRef]
- Stuchlik, A.; Sumiyoshi, T. Cognitive Deficits in Schizophrenia and Other Neuropsychiatric Disorders: Convergence of Preclinical and Clinical Evidence. Front. Behav. Neurosci. 2014, 8, 444. [Google Scholar] [CrossRef]
- Van Holst, R.J.; Schilt, T. Drug-related decrease in neuropsychological functions of abstinent drug users. Curr. Drug Abus. Rev. 2011, 4, 42–56. [Google Scholar] [CrossRef]
- Fernández-Serrano, M.J.; Pérez-García, M.; Verdejo-García, A. What are the specific vs. generalized effects of drugs of abuse on neuropsychological performance? Neurosci. Biobehav. Rev. 2011, 35, 377–406. [Google Scholar] [CrossRef]
- Solomon, M.; Gordon, A.; Iosif, A.; Geddert, R.; Krug, M.K.; Mundy, P.; Hessl, D. Using the NIH Toolbox to Assess Cognition in Adolescents and Young Adults with Autism Spectrum Disorders. Autism Res. 2021, 14, 500–511. [Google Scholar] [CrossRef]
- LaLonde, R. The neurobiological basis of spontaneous alternation. Neurosci. Biobehav. Rev. 2002, 26, 91–104. [Google Scholar] [CrossRef]
- Dudchenko, P.A. An overview of the tasks used to test working memory in rodents. Neurosci. Biobehav. Rev. 2004, 28, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, K.C. A test of two explanations of spontaneous alternation. J. Comp. Physiol. Psychol. 1952, 45, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Dudchenko, P.A. How do animals actually solve the T maze? Behav Neurosci. 2001, 115, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Korotkova, T.; Fuchs, E.C.; Ponomarenko, A.; von Engelhardt, J.; Monyer, H. NMDA Receptor Ablation on Parvalbumin-Positive Interneurons Impairs Hippocampal Synchrony, Spatial Representations, and Working Memory. Neuron 2010, 68, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Groves, D.A.; Bowman, E.; Brown, V. Recordings from the rat locus coeruleus during acute vagal nerve stimulation in the anaesthetised rat. Neurosci. Lett. 2005, 379, 174–179. [Google Scholar] [CrossRef]
- Follesa, P.; Biggio, F.; Gorini, G.; Caria, S.; Talani, G.; Dazzi, L.; Puligheddu, M.; Marrosu, F.; Biggio, G. Vagus nerve stimulation increases norepinephrine concentration and the gene expression of BDNF and bFGF in the rat brain. Brain Res. 2007, 1179, 28–34. [Google Scholar] [CrossRef]
- Huffman, W.J.; Subramaniyan, S.; Rodriguiz, R.M.; Wetsel, W.C.; Grill, W.M.; Terrando, N. Modulation of neuroinflammation and memory dysfunction using percutaneous vagus nerve stimulation in mice. Brain Stimul. 2019, 12, 19–29. [Google Scholar] [CrossRef]
- Vázquez-Oliver, A.; Brambilla-Pisoni, C.; Domingo-Gainza, M.; Maldonado, R.; Ivorra, A.; Ozaita, A. Auricular transcutaneous vagus nerve stimulation improves memory persistence in naïve mice and in an intellectual disability mouse model. Brain Stimul. 2020, 13, 494–498. [Google Scholar] [CrossRef]
- Ghacibeh, G.A.; Shenker, J.I.; Shenal, B.; Uthman, B.M.; Heilman, K.M. The Influence of Vagus Nerve Stimulation on Memory. Cogn. Behav. Neurol. 2006, 19, 119–122. [Google Scholar] [CrossRef]
- Mathew, E.; Tabet, M.N.; Robertson, N.M.; Hays, S.A.; Rennaker, R.L.; Kilgard, M.P.; McIntyre, C.K.; Souza, R.R. Vagus nerve stimulation produces immediate dose-dependent anxiolytic effect in rats. J. Affect. Disord. 2020, 265, 552–557. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Hulsey, D.R.; Shedd, C.M.; Sarker, S.F.; Kilgard, M.P.; Hays, S.A. Norepinephrine and serotonin are required for vagus nerve stimulation directed cortical plasticity. Exp. Neurol. 2019, 320, 112975. [Google Scholar] [CrossRef] [PubMed]
- Usami, K.; Kano, R.; Kawai, K.; Noda, T.; Shiramatsu, T.I.; Saito, N.; Takahashi, H. Modulation of cortical synchrony by vagus nerve stimulation in adult rats. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2013, 2013, 5348–5351. [Google Scholar] [CrossRef] [PubMed]
- Schuman, E.M. Neurotrophin regulation of synaptic transmission. Curr. Opin. Neurobiol. 1999, 9, 105–109. [Google Scholar] [CrossRef]
- Schinder, A. The neurotrophin hypothesis for synaptic plasticity. Trends Neurosci. 2000, 23, 639–645. [Google Scholar] [CrossRef]
- Bai, J.; Otani, S. Neuroplasticity Regulation by Noradrenaline in Mammalian Brain. Curr. Neuropharmacol. 2009, 7, 286–295. [Google Scholar] [CrossRef]
- Palacios-Filardo, J.; Mellor, J.R. Neuromodulation of hippocampal long-term synaptic plasticity. Curr. Opin. Neurobiol. 2019, 54, 37–43. [Google Scholar] [CrossRef]
- Zuo, Y.; Smith, D.C.; Jensen, R.A. Vagus nerve stimulation potentiates hippocampal LTP in freely-moving rats. Physiol. Behav. 2007, 90, 583–589. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Shipley, M.; Chouvet, G.; Ennis, M.; van Bockstaele, E.; Pieribone, V.; Shiekhattar, R.; Akaoka, H.; Drolet, G.; Astier, B.; et al. Afferent regulation of locus coeruleus neurons: Anatomy, physiology and pharmacology. Prog. Brain Res. 1991, 88, 47–75. [Google Scholar] [CrossRef]
- Hulsey, D.R.; Riley, J.R.; Loerwald, K.W.; Rennaker, R.L.; Kilgard, M.P.; Hays, S.A. Parametric characterization of neural activity in the locus coeruleus in response to vagus nerve stimulation. Exp. Neurol. 2017, 289, 21–30. [Google Scholar] [CrossRef]
- Van Bockstaele, E.J.; Peoples, J.; Telegan, P. Efferent projections of the nucleus of the solitary tract to peri-locus coeruleus dendrites in rat brain: Evidence for a monosynaptic pathway. J. Comp. Neurol. 1999, 412, 410–428. [Google Scholar] [CrossRef]
- Mello-Carpes, P.B.; Izquierdo, I. The Nucleus of the Solitary Tract → Nucleus Paragigantocellularis→Locus Coeruleus→CA1 region of dorsal hippocampus pathway is important for consolidation of object recognition memory. Neurobiol. Learn. Mem. 2013, 100, 56–63. [Google Scholar] [CrossRef]
- Clayton, E.C.; Williams, C.L. Adrenergic activation of the nucleus tractus solitarius potentiates amygdala norepinephrine release and enhances retention performance in emotionally arousing and spatial memory tasks. Behav. Brain Res. 2000, 112, 151–158. [Google Scholar] [CrossRef]
- Clayton, E.C.; Williams, C.L. Noradrenergic Receptor Blockade of the NTS Attenuates the Mnemonic Effects of Epinephrine in an Appetitive Light–Dark Discrimination Learning Task. Neurobiol. Learn. Mem. 2000, 74, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.L.; Men, D.; Clayton, E.C. The effects of noradrenergic activation of the nucleus tractus solitarius on memory and in potentiating norepinephrine release in the amygdala. Behav. Neurosci. 2000, 114, 1131–1144. [Google Scholar] [CrossRef]
- Beste, C.; Steenbergen, L.; Sellaro, R.; Grigoriadou, S.; Zhang, R.; Chmielewski, W.; Stock, A.-K.; Colzato, L. Effects of Concomitant Stimulation of the GABAergic and Norepinephrine System on Inhibitory Control—A Study Using Transcutaneous Vagus Nerve Stimulation. Brain Stimul. 2016, 9, 811–818. [Google Scholar] [CrossRef]
- Dietrich, S.; Smith, J.; Scherzinger, C.; Hofmann-Preiss, K.; Freitag, T.; Eisenkolb, A.; Ringler, R. A novel transcutaneous vagus nerve stimulation leads to brainstem and cerebral activations measured by functional MRI. Biomed. Tech. Biomed. Eng. 2008, 53, 104–111. [Google Scholar] [CrossRef]
- Idiaquez, J.; Roman, G.C. Autonomic dysfunction in neurodegenerative dementias. J. Neurol. Sci. 2011, 305, 22–27. [Google Scholar] [CrossRef]
- Allan, L.M.; Ballard, C.; Allen, J.; Murray, A.; Davidson, A.W.; McKeith, I.; Kenny, R.A. Autonomic dysfunction in dementia. J. Neurol. Neurosurg. Psychiatry 2006, 78, 671–677. [Google Scholar] [CrossRef]
- Kemp, A.H.; Quintana, D.S.; Gray, M.A.; Felmingham, K.L.; Brown, K.; Gatt, J.M. Impact of Depression and Antidepressant Treatment on Heart Rate Variability: A Review and Meta-Analysis. Biol. Psychiatry 2010, 67, 1067–1074. [Google Scholar] [CrossRef]
- Jandackova, V.K.; Britton, A.; Malik, M.; Steptoe, A. Heart rate variability and depressive symptoms: A cross-lagged analysis over a 10-year period in the Whitehall II study. Psychol. Med. 2016, 46, 2121–2131. [Google Scholar] [CrossRef]
- Rush, A.J.; Marangell, L.B.; Sackeim, H.A.; George, M.S.; Brannan, S.K.; Davis, S.M.; Howland, R.; Kling, M.A.; Rittberg, B.R.; Burke, W.J.; et al. Vagus Nerve Stimulation for Treatment-Resistant Depression: A Randomized, Controlled Acute Phase Trial. Biol. Psychiatry 2005, 58, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Guo, Y.-M.; Ning, Y.-P.; Cao, L.-P.; Rao, Y.-H.; Sun, J.-Q.; Qing, M.-J.; Zheng, W. Adjunctive vagus nerve stimulation for treatment-resistant depression: A preliminary study. Int. J. Psychiatry Clin. Pract. 2022, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Jodoin, V.D.; Richer, F.; Miron, J.-P.; Fournier-Gosselin, M.-P.; Lespérance, P. Long-term Sustained Cognitive Benefits of Vagus Nerve Stimulation in Refractory Depression. J. ECT 2018, 34, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, H.I.; Riphagen, J.M.; Razat, C.M.; Wiese, S.; Sack, A. Transcutaneous vagus nerve stimulation boosts associative memory in older individuals. Neurobiol. Aging 2015, 36, 1860–1867. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Ventura-Bort, C.; Hamm, A.; Weymar, M. Transcutaneous vagus nerve stimulation (tVNS) enhances conflict-triggered adjustment of cognitive control. Cogn. Affect. Behav. Neurosci. 2018, 18, 680–693. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Driskill, C.M.; Childs, J.E.; Itmer, B.; Rajput, J.S.; Kroener, S. Acute Vagus Nerve Stimulation Facilitates Short Term Memory and Cognitive Flexibility in Rats. Brain Sci. 2022, 12, 1137. https://doi.org/10.3390/brainsci12091137
Driskill CM, Childs JE, Itmer B, Rajput JS, Kroener S. Acute Vagus Nerve Stimulation Facilitates Short Term Memory and Cognitive Flexibility in Rats. Brain Sciences. 2022; 12(9):1137. https://doi.org/10.3390/brainsci12091137
Chicago/Turabian StyleDriskill, Christopher M., Jessica E. Childs, Bemisal Itmer, Jai S. Rajput, and Sven Kroener. 2022. "Acute Vagus Nerve Stimulation Facilitates Short Term Memory and Cognitive Flexibility in Rats" Brain Sciences 12, no. 9: 1137. https://doi.org/10.3390/brainsci12091137
APA StyleDriskill, C. M., Childs, J. E., Itmer, B., Rajput, J. S., & Kroener, S. (2022). Acute Vagus Nerve Stimulation Facilitates Short Term Memory and Cognitive Flexibility in Rats. Brain Sciences, 12(9), 1137. https://doi.org/10.3390/brainsci12091137