
Beneficial Influence of Exendin-4 on Specific Organs and Mechanisms Favourable for the Elderly with Concomitant Obstructive Lung Diseases

Abstract
1. Introduction
2. General Characteristics of Ex-4
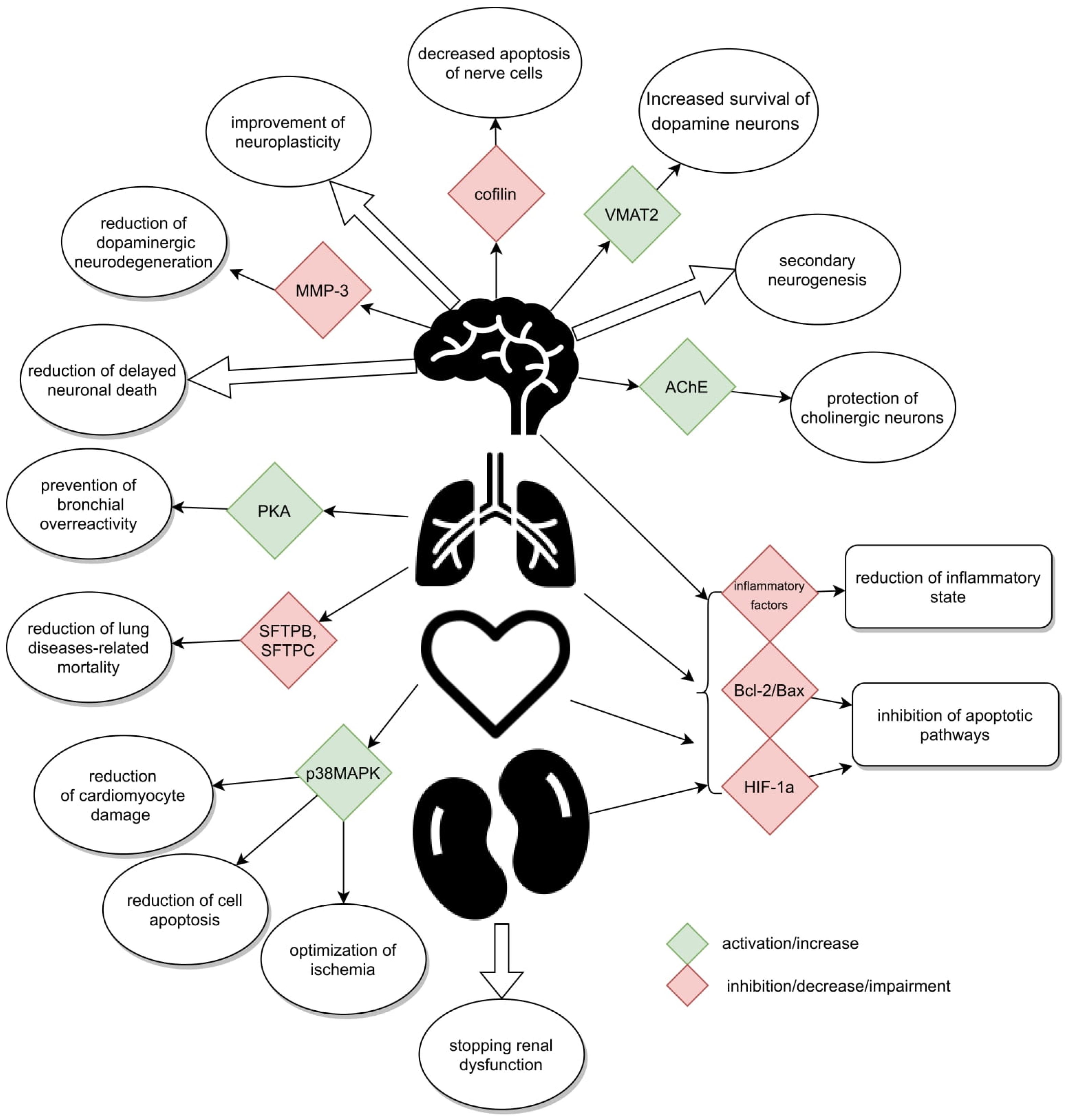
3. Actions against Neurodegeneration of Cerebral Tissue
4. Protective Action of Ex-4 in Hypoxia of Various Tissues
5. Anti-Inflammatory Properties of Ex-4
6. Beneficial Effects in Chronic Obstructive Pulmonary Disease (COPD)
7. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MMP-3 | metalloproteinase 3 |
| TNF-α | tumour necrosis factor α |
| IL-1β | interleukin 1β |
| VMAT2 | vesicular monoamine transporter 2 |
| LTP | long-term potential |
| ALS | amyotrophic lateral sclerosis |
| p38MAPK | p38 mitogen-activated protein kinase |
| LDH | lactate dehydrogenase |
| CK-MB | creatine kinase MB |
| GLUT-4 | glucose transporter 4 |
| GPxR | glutathione peroxidase |
| TRxR | thioredoxin reductase |
| GLP-1R | receptor for GLP-1 |
| ROS | reactive oxygen species |
| JNK-1 | c-Jun N-terminal kinase 1 |
| TLR-2 | Toll-like receptor-2 |
| TLR-4 | Toll-like receptor-4 |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| SOCS-3 | suppressor of cytokine signalling 3 |
| PKA | cAMP-dependent protein kinase A |
| COPD | chronic obstructive pulmonary disease |
| MCP-1 | monocyte chemoattractant protein-1 |
| IL-6 | interleukin 6 |
| IL-10 | interleukin 10 |
| SFTPB, SFTPC, SFTPD | surfactant proteins B, C, D |
| GLP-1 | glucagon-like protein-1 |
| PI3K | phosphoinositide 3-kinase |
| cAMP | cyclic adenosine monophosphate |
| CREB | cAMP-response element-binding protein |
| Bcl-2/Bax | B-cell lymphoma-2 |
| AKT | protein kinase b |
| HIF-1a | Hypoxia-inducible factor-1α |
| BP | blood pressure |
| HR | heart rate |
| AChE | acetylcholinesterase |
References
- Athauda, D.; Maclagan, K.; Skene, S.S.; Bajwa-Joseph, M.; Letchford, D.; Chowdhury, K.; Hibbert, S.; Budnik, N.; Zampedri, L.; Dickson, J.; et al. Exenatide once weekly versus placebo in Parkinson’s disease: A randomised, double-blind, placebo-controlled trial. Lancet 2017, 390, 1664–1675. [Google Scholar] [CrossRef]
- Tysnes, O.-B.; Storstein, A. Epidemiology of Parkinson’s disease. J. Neural Transm. 2017, 124, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.; Lee, B.Y.; Hane, F.T. Recent Progress in Alzheimer’s Disease Research, Part 2: Genetics and Epidemiology. J. Alzheimer’s Dis. 2017, 57, 317–330. [Google Scholar] [CrossRef]
- McCormack, P.L. Exenatide Twice Daily: A Review of Its Use in the Management of Patients with Type 2 Diabetes Mellitus. Drugs 2014, 74, 325–351. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chigurupati, S.; Holloway, H.W.; Mughal, M.; Tweedie, D.; Bruestle, D.A.; Mattson, M.P.; Wang, Y.; Harvey, B.K.; Ray, B.; et al. Exendin-4 Ameliorates Motor Neuron Degeneration in Cellular and Animal Models of Amyotrophic Lateral Sclerosis. PLoS ONE 2012, 7, e32008. [Google Scholar] [CrossRef]
- Eng, J.; Kleinman, W.A.; Singh, L.; Singh, G.; Raufman, J.P. Isolation and Characterization of Exendin-4, an Exendin-3 Analogue, from Heloderma Suspectum Venom: Further Evidence for an Exendin Receptor on Dispersed Acini from Guinea Pig Pancreas. J. Biol. Chem. 1992, 267, 7402–7405. [Google Scholar] [CrossRef]
- Göke, R.; Fehmann, H.C.; Linn, T.; Schmidt, H.; Eng, J.; Göke, B. Exendin 4 Is a High-Potency Agonist and Truncated Exendin (9-39)Amide an Antagonist at the GLP-1 (7-36)Amide Receptor of Insulin-Secreting Beta-Cells. Digestion 1993, 54, 341–347. [Google Scholar] [CrossRef]
- Kastin, A.J.; Akerstrom, V. Entry of exendin-4 into brain is rapid but may be limited at high doses. Int. J. Obes. 2003, 27, 313–318. [Google Scholar] [CrossRef]
- Hui, H.; Farilla, L.; Merkel, P.; Perfetti, R. The Short Half-Life of Glucagon-like Peptide-1 in Plasma Does Not Reflect Its Long-Lasting Beneficial Effects. Eur. J. Endocrinol. 2002, 146, 863–869. [Google Scholar] [CrossRef]
- Greiner, T.U.; Bäckhed, F. Microbial regulation of GLP-1 and L-cell biology. Mol. Metab. 2016, 5, 753–758. [Google Scholar] [CrossRef]
- Tian, L.; Jin, T. The Incretin Hormone GLP-1 and Mechanisms Underlying Its Secretion: GLP-1. J. Diabetes 2016, 8, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Young, A.A.; Gedulin, B.R.; Bhavsar, S.; Bodkin, N.; Jodka, C.; Hansen, B.; Denaro, M. Glucose-lowering and insulin-sensitizing actions of exendin-4: Studies in obese diabetic (ob/ob, db/db) mice, diabetic fatty Zucker rats, and diabetic rhesus monkeys (Macaca mulatta). Diabetes 1999, 48, 1026–1034. [Google Scholar] [CrossRef] [PubMed]
- Greig, N.H.; Holloway, H.W.; De Ore, K.A.; Jani, D.; Wang, Y.; Zhou, J.; Garant, M.J.; Egan, J.M. Once daily injection of exendin-4 to diabetic mice achieves long-term beneficial effects on blood glucose concentrations. Diabetologia 1999, 42, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Thorens, B.; Porret, A.; Buhler, L.E.O.; Deng, S.; Morel, P.; Widmann, C. Cloning and Fungctional Expression of the Human Islet GLP-1 Receptor. Diabetes 1993, 42, 1678–1682. [Google Scholar] [CrossRef]
- Yamamoto, H.; Lee, C.E.; Marcus, J.N.; Williams, T.D.; Overton, J.M.; Lopez, M.E.; Hollenberg, A.N.; Baggio, L.; Saper, C.B.; Drucker, D.J.; et al. Glucagon-like peptide-1 receptor stimulation increases blood pressure and heart rate and activates autonomic regulatory neurons. J. Clin. Investig. 2002, 110, 43–52. [Google Scholar] [CrossRef]
- Richter, G.; Feddersen, O.; Wagner, U.; Barth, P.; Göke, R.; Göke, B. GLP-1 stimulates secretion of macromolecules from airways and relaxes pulmonary artery. Am. J. Physiol. Cell. Mol. Physiol. 1993, 265, L374–L381. [Google Scholar] [CrossRef]
- Brierley, D.I.; de Lartigue, G. Reappraising the role of the vagus nerve in GLP-1-mediated regulation of eating. J. Cereb. Blood Flow Metab. 2021, 179, 584–599. [Google Scholar] [CrossRef]
- Gong, N.; Xiao, Q.; Zhu, B.; Zhang, C.-Y.; Wang, Y.-X.; Fan, H.; Ma, A.-N. Activation of Spinal Glucagon-Like Peptide-1 Receptors Specifically Suppresses Pain Hypersensitivity. J. Neurosci. 2014, 34, 5322–5334. [Google Scholar] [CrossRef]
- Zhang, D.; Lv, G. Therapeutic potential of spinal GLP-1 receptor signaling. Peptides 2018, 101, 89–94. [Google Scholar] [CrossRef]
- Ma, L.; Ju, P.; Wang, W.; Wei, J.; Wang, W.; Zhao, M.; Ahmad, K.A.; Wang, Y.; Chen, J. Microglial Activation of GLP-1R Signaling in Neuropathic Pain Promotes Gene Expression Adaption Involved in Inflammatory Responses. Neural Plast. 2021, 2021, 9923537. [Google Scholar] [CrossRef]
- Wagner, U.; Bredenbröker, D.; Storm, B.; Tackenberg, B.; Fehmann, H.-C.; von Wichert, P. Effects of VIP and Related Peptides on Airway Mucus Secretion from Isolated Rat Trachea. Peptides 1998, 19, 241–245. [Google Scholar] [CrossRef]
- Rogliani, P.; Calzetta, L.; Capuani, B.; Facciolo, F.; Cazzola, M.; Lauro, D.; Matera, M.G. Glucagon-Like Peptide 1 Receptor: A Novel Pharmacological Target for Treating Human Bronchial Hyperresponsiveness. Am. J. Respir. Cell Mol. Biol. 2016, 55, 804–814. [Google Scholar] [CrossRef]
- Wei, Y.; Mojsov, S. Tissue-specific expression of the human receptor for glucagon-like peptide-I: Brain, heart and pancreatic forms have the same deduced amino acid sequences. FEBS Lett. 1995, 358, 219–224. [Google Scholar] [CrossRef]
- Roan, J.-N.; Hsu, C.-H.; Fang, S.-Y.; Tsai, H.-W.; Luo, C.-Y.; Huang, C.-C.; Lam, C.-F. Exendin-4 improves cardiovascular function and survival in flow-induced pulmonary hypertension. J. Thorac. Cardiovasc. Surg. 2018, 155, 1661–1669.e4. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.; Candeias, E.; Correia, S.; Santos, R.; Carvalho, C.; Cardoso, S.; Plácido, A.; Santos, M.; Oliveira, C.; Moreira, P. Crosstalk between diabetes and brain: Glucagon-like peptide-1 mimetics as a promising therapy against neurodegeneration. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2013, 1832, 527–541. [Google Scholar] [CrossRef]
- Luciani, P.; Deledda, C.; Benvenuti, S.; Cellai, I.; Squecco, R.; Monici, M.; Cialdai, F.; Luciani, G.; Danza, G.; Di Stefano, C.; et al. Differentiating effects of the glucagon-like peptide-1 analogue exendin-4 in a human neuronal cell model. Experientia 2010, 67, 3711–3723. [Google Scholar] [CrossRef]
- Foltynie, T.; Aviles-Olmos, I. Exenatide as a potential treatment for patients with Parkinson’s disease: First steps into the clinic. Alzheimer’s Dement. 2014, 10, S38–S46. [Google Scholar] [CrossRef]
- Kanse, S.M.; Kreymann, B.; Ghatei, M.A.; Bloom, S.R. Identification and characterization of glucagon-like peptide-1 7-36 amide-binding sites in the rat brain and lung. FEBS Lett. 1988, 241, 209–212. [Google Scholar] [CrossRef]
- Ayoub, W.A.; Kumar, A.A.; Naguib, H.S.; Taylor, H.C. Exenatide-Induced Acute Pancreatitis. Endocr. Pract. 2010, 16, 80–83. [Google Scholar] [CrossRef]
- Kim, D.S.; Choi, H.-I.; Wang, Y.; Luo, Y.; Hoffer, B.J.; Greig, N.H. A New Treatment Strategy for Parkinson’s Disease through the Gut–Brain Axis. Cell Transplant. 2017, 26, 1560–1571. [Google Scholar] [CrossRef]
- Hamilton, A.; Patterson, S.; Porter, D.; Gault, V.; Holscher, C. Novel GLP-1 mimetics developed to treat type 2 diabetes promote progenitor cell proliferation in the brain. J. Neurosci. Res. 2011, 89, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Moon, M.; Park, S. Exendin-4 protects dopaminergic neurons by inhibition of microglial activation and matrix metalloproteinase-3 expression in an animal model of Parkinson’s disease. J. Endocrinol. 2009, 202, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Mogi, M.; Harada, M.; Kondo, T.; Riederer, P.; Inagaki, H.; Minami, M.; Nagatsu, T. Interleukin-1β, interleukin-6, epidermal growth factor and transforming growth factor-α are elevated in the brain from parkinsonian patients. Neurosci. Lett. 1994, 180, 147–150. [Google Scholar] [CrossRef]
- Hunot, S.; Boissière, F.; Faucheux, B.; Brugg, B.; Mouatt-Prigent, A.; Agid, Y.; Hirsch, E. Nitric oxide synthase and neuronal vulnerability in parkinson’s disease. Neuroscience 1996, 72, 355–363. [Google Scholar] [CrossRef]
- Bessler, H.; Djaldetti, R.; Salman, H.; Bergman, M.; Djaldetti, M. IL-1β, IL-2, IL-6 and TNF-α production by peripheral blood mononuclear cells from patients with Parkinson’s disease. Biomed. Pharmacother. 1999, 53, 141–145. [Google Scholar] [CrossRef]
- Bertilsson, G.; Patrone, C.; Zachrisson, O.; Andersson, A.; Dannaeus, K.; Heidrich, J.; Kortesmaa, J.; Mercer, A.; Nielsen, E.; Rönnholm, H.; et al. Peptide hormone exendin-4 stimulates subventricular zone neurogenesis in the adult rodent brain and induces recovery in an animal model of parkinson’s disease. J. Neurosci. Res. 2007, 86, 326–338. [Google Scholar] [CrossRef]
- Chen, S.; Yu, S.-J.; Li, Y.; Lecca, D.; Glotfelty, E.; Kim, H.K.; Choi, H.-I.; Hoffer, B.J.; Greig, N.H.; Kim, D.S.; et al. Post-treatment with PT302, a long-acting Exendin-4 sustained release formulation, reduces dopaminergic neurodegeneration in a 6-Hydroxydopamine rat model of Parkinson’s disease. Sci. Rep. 2018, 8, 10722. [Google Scholar] [CrossRef]
- Li, Y.; Tweedie, D.; Mattson, M.P.; Holloway, H.W.; Greig, N.H. Enhancing the GLP-1 receptor signaling pathway leads to proliferation and neuroprotection in human neuroblastoma cells. J. Neurochem. 2010, 113, 1621–1631. [Google Scholar] [CrossRef]
- Li, H.; Lee, C.H.; Yoo, K.-Y.; Choi, J.H.; Park, O.K.; Yan, B.C.; Byun, K.; Lee, B.; Hwang, I.K.; Won, M.-H. Chronic treatment of exendin-4 affects cell proliferation and neuroblast differentiation in the adult mouse hippocampal dentate gyrus. Neurosci. Lett. 2010, 486, 38–42. [Google Scholar] [CrossRef]
- Perry, T.; Haughey, N.J.; Mattson, M.P.; Egan, J.M.; Greig, N.H. Protection and Reversal of Excitotoxic Neuronal Damage by Glucagon-Like Peptide-1 and Exendin-4. J. Pharmacol. Exp. Ther. 2002, 302, 881–888. [Google Scholar] [CrossRef]
- Wenger, R.H.; Gassmann, M. Oxygen(es) and the hypoxia-inducible factor-1. Biol. Chem. 1997, 378, 609–616. [Google Scholar] [PubMed]
- Jin, J.; Kang, H.-M.; Jung, J.; Jeong, J.-W.; Park, C. Related expressional change of HIF-1α to the neuroprotective activity of exendin-4 in transient global ischemia. NeuroReport 2014, 25, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Yan, B.; Yoo, K.-Y.; Choi, J.H.; Kwon, S.-H.; Her, S.; Sohn, Y.; Hwang, I.K.; Cho, J.H.; Kim, Y.-M.; et al. Ischemia-induced changes in glucagon-like peptide-1 receptor and neuroprotective effect of its agonist, exendin-4, in experimental transient cerebral ischemia. J. Neurosci. Res. 2011, 89, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Lü, K.; Chang, G.; Ye, L.; Zhang, P.; Li, Y.; Zhang, D. Protective effects of extendin-4 on hypoxia/reoxygenation-induced injury in H9c2 cells. Mol. Med. Rep. 2015, 12, 3007–3016. [Google Scholar] [CrossRef]
- Padmasekar, M.; Lingwal, N.; Samikannu, B.; Chen, C.; Sauer, H.; Linn, T. Exendin-4 Protects Hypoxic Islets From Oxidative Stress and Improves Islet Transplantation Outcome. Endocrinology 2013, 154, 1424–1433. [Google Scholar] [CrossRef]
- Chen, Y.-T.; Tsai, T.-H.; Yang, C.-C.; Sun, C.-K.; Chang, L.-T.; Chen, H.-H.; Chang, C.-L.; Sung, P.-H.; Zhen, Y.-Y.; Leu, S.; et al. Exendin-4 and sitagliptin protect kidney from ischemia-reperfusion injury through suppressing oxidative stress and inflammatory reaction. J. Transl. Med. 2013, 11, 270. [Google Scholar] [CrossRef]
- Waddell, J.A.; Emerson, P.A.; Gunstone, R.F. Hypoxia in bronchial asthma. BMJ 1967, 2, 402–404. [Google Scholar] [CrossRef][Green Version]
- Ahmad, T.; Kumar, M.; Mabalirajan, U.; Pattnaik, B.; Aggarwal, S.; Singh, R.; Singh, S.; Mukerji, M.; Ghosh, B.; Agrawal, A. Hypoxia Response in Asthma: Differential modulation on inflammation and epithelial injury. Am. J. Respir. Cell Mol. Biol. 2012, 47, 1–10. [Google Scholar] [CrossRef]
- Chaudhuri, A.; Ghanim, H.; Vora, M.; Sia, C.L.; Korzeniewski, K.; Dhindsa, S.; Makdissi, A.; Dandona, P. Exenatide Exerts a Potent Antiinflammatory Effect. J. Clin. Endocrinol. Metab. 2012, 97, 198–207. [Google Scholar] [CrossRef]
- Nguyen, D.-V.; Linderholm, A.; Haczku, A.; Kenyon, N. Obesity-related, metabolic asthma: A new role for glucagon-like peptide 1 agonists. Lancet Respir. Med. 2017, 5, 162–164. [Google Scholar] [CrossRef]
- Rogliani, P.; Calzetta, L.; Ora, J.; Cazzola, M.; Lauro, D.; Matera, M.G.; Coppola, A. Effect of exendin-4 on lung function of diabetic patients: A preliminary report. Eur. Respir. J. 2017, 50, PA1039. [Google Scholar] [CrossRef]
- Viby, N.-E.; Isidor, M.S.L.; Buggeskov, K.B.; Poulsen, S.S.; Hansen, J.; Kissow, H. Glucagon-Like Peptide-1 (GLP-1) Reduces Mortality and Improves Lung Function in a Model of Experimental Obstructive Lung Disease in Female Mice. Endocrinology 2013, 154, 4503–4511. [Google Scholar] [CrossRef] [PubMed]
- Vara, E.; Arias-díaz, J.; Garcia, C.; Balibrea, J.L.; Blázquez, E. Surfactant Secretion in Human Type II Pneumocytes. Crit. Care Med. 2001, 1, 1–7. [Google Scholar]
- Rooney, S.A.; Young, S.L.; Mendelson, C.R. Molecular and cellular processing of lung surfactant. FASEB J. 1994, 8, 957–967. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristics | GLP-1 | Ex-4 |
|---|---|---|
| Origin | Endogenous | Exogenous |
| Site of generation | L-cells in intestines | Salivary glands of Heloderma suspectum lizard |
| Structure | - | 53% similar to GLP-1 |
| Receptor | GLP-1 | GLP-1, VIP proteins |
| Affinity to GLP-1R | - | Higher than GLP-1 |
| Half-time | 1.5–5 min [9] | 120 min |
| Amount of cAMP secretion | - | Three times higher |
| Beneficial Effects | Adverse Effects |
|---|---|
| Stimulation of proinsulin gene expression at the transcription level | Increased blood pressure |
| Release of insulin reserves from pancreatic cells | Increased heart rate |
| Inhibition of glucagon secretion by the liver | Aversion to taste |
| Improvement in insulin sensitivity | Delayed gastric emptying |
| Reduction of glycated haemoglobin levels | Slackening of peristaltic movements of the stomach and intestines |
| Body weight loss | Pancreatitis |
| No action at low glycaemic values |
| Mechanism of Action | Potential Therapeutic Procedures |
|---|---|
| Prevention and treatment of neurodegenerative diseases |
| ALS prevention and treatment |
| Treatment of complications due to organ ischemia |
| Treatment of ischemic strokes |
| Prevention of myocardial infarcts |
| Prevention of complications of thromboembolism, vascular atherosclerosis |
| Improvement in asthma control |
| COPD treatment |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figat, M.; Kardas, G.; Kuna, P.; Panek, M.G. Beneficial Influence of Exendin-4 on Specific Organs and Mechanisms Favourable for the Elderly with Concomitant Obstructive Lung Diseases. Brain Sci. 2022, 12, 1090. https://doi.org/10.3390/brainsci12081090
Figat M, Kardas G, Kuna P, Panek MG. Beneficial Influence of Exendin-4 on Specific Organs and Mechanisms Favourable for the Elderly with Concomitant Obstructive Lung Diseases. Brain Sciences. 2022; 12(8):1090. https://doi.org/10.3390/brainsci12081090
Chicago/Turabian StyleFigat, Magdalena, Grzegorz Kardas, Piotr Kuna, and Michał G. Panek. 2022. "Beneficial Influence of Exendin-4 on Specific Organs and Mechanisms Favourable for the Elderly with Concomitant Obstructive Lung Diseases" Brain Sciences 12, no. 8: 1090. https://doi.org/10.3390/brainsci12081090
APA StyleFigat, M., Kardas, G., Kuna, P., & Panek, M. G. (2022). Beneficial Influence of Exendin-4 on Specific Organs and Mechanisms Favourable for the Elderly with Concomitant Obstructive Lung Diseases. Brain Sciences, 12(8), 1090. https://doi.org/10.3390/brainsci12081090