
Dopamine and Beyond: Implications of Psychophysical Studies of Intracranial Self-Stimulation for the Treatment of Depression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Animal Models of Depression: The Bright Side
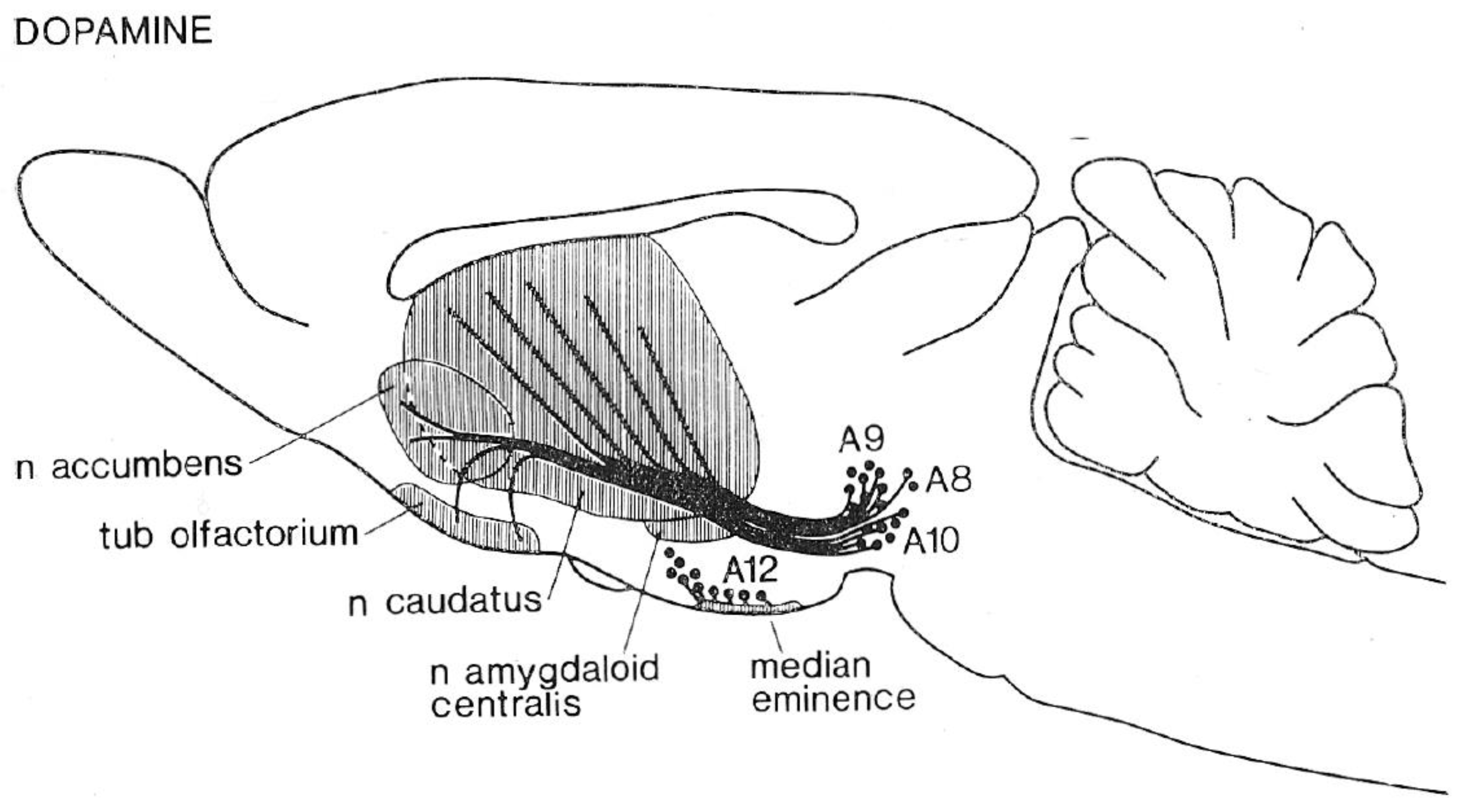
3. Dopamine Transmission and Depression
4. Brain Stimulation Reward
4.1. Phase 1: Direct Stimulation of Dopaminergic Axons
4.2. Phase 2: The Series-Circuit Model
4.3. Phase 3: The Convergence Model
5. Contribution of Midbrain Dopamine Neurons to the Treatment of Depression: One Pathway among Many?
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McGirr, A.; Renaud, J.; Seguin, M.; Alda, M.; Benkelfat, C.; Lesage, A.; Turecki, G. An Examination of DSM-IV Depressive Symptoms and Risk for Suicide Completion in Major Depressive Disorder: A Psychological Autopsy Study. J. Affect. Disord. 2007, 97, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Bretschneider, J.; Janitza, S.; Jacobi, F.; Thom, J.; Hapke, U.; Kurth, T.; Maske, U.E. Time Trends in Depression Prevalence and Health-Related Correlates: Results from Population-Based Surveys in Germany 1997–1999 vs. 2009–2012. BMC Psychiatry 2018, 18, 394. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, B.H. Depression as a Disease of Modernity: Explanations for Increasing Prevalence. J. Affect. Disord. 2012, 140, 205. [Google Scholar] [CrossRef]
- Murphy, J.M.; Laird, N.M.; Monson, R.R.; Sobol, A.M.; Leighton, A.H. A 40-Year Perspective on the Prevalence of Depression: The Stirling County Study. Arch. Gen. Psychiatry 2000, 57, 209–215. [Google Scholar] [CrossRef]
- Duffy, M.E.; Twenge, J.M.; Joiner, T.E. Trends in Mood and Anxiety Symptoms and Suicide-Related Outcomes Among U.S. Undergraduates, 2007-2018: Evidence From Two National Surveys. J. Adolesc. Health Off. Publ. Soc. Adolesc. Med. 2019, 65, 590–598. [Google Scholar] [CrossRef]
- Santomauro, D.F.; Herrera, A.M.M.; Shadid, J.; Zheng, P.; Ashbaugh, C.; Pigott, D.M.; Abbafati, C.; Adolph, C.; Amlag, J.O.; Aravkin, A.Y.; et al. Global Prevalence and Burden of Depressive and Anxiety Disorders in 204 Countries and Territories in 2020 Due to the COVID-19 Pandemic. Lancet 2021, 398, 1700–1712. [Google Scholar] [CrossRef]
- López-Muñoz, F.; D’Ocón, P.; Romero, A.; Guerra, J.A.; Álamo, C. Role of Serendipity in the Discovery of Classical Antidepressant Drugs: Applying Operational Criteria and Patterns of Discovery. World J. Psychiatry 2022, 12, 588–602. [Google Scholar] [CrossRef]
- Filatova, E.V.; Shadrina, M.I.; Slominsky, P.A.; Faron-Górecka, A.; Dziedzicka-Wasylewska, M. Major Depression: One Brain, One Disease, One Set of Intertwined Processes. Cells 2021, 10, 1283. [Google Scholar] [CrossRef]
- Gaynes, B.N.; Lux, L.; Gartlehner, G.; Asher, G.; Forman-Hoffman, V.; Green, J.; Boland, E.; Weber, R.P.; Randolph, C.; Bann, C.; et al. Defining Treatment-Resistant Depression. Depress. Anxiety 2020, 37, 134–145. [Google Scholar] [CrossRef]
- Drobisz, D.; Damborská, A. Deep Brain Stimulation Targets for Treating Depression. Behav. Brain Res. 2019, 359, 266–273. [Google Scholar] [CrossRef]
- Coenen, V.A.; Bewernick, B.H.; Kayser, S.; Kilian, H.; Boström, J.; Greschus, S.; Hurlemann, R.; Klein, M.E.; Spanier, S.; Sajonz, B.; et al. Superolateral Medial Forebrain Bundle Deep Brain Stimulation in Major Depression: A Gateway Trial. Neuropsychopharmacology 2019, 44, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Fenoy, A.J.; Quevedo, J.; Soares, J.C. Deep Brain Stimulation of the “Medial Forebrain Bundle”: A Strategy to Modulate the Reward System and Manage Treatment-Resistant Depression. Mol. Psychiatry 2021, 27, 574–592. [Google Scholar] [CrossRef]
- Coenen, V.A.; Panksepp, J.; Hurwitz, T.A.; Urbach, H.; Mädler, B. Human Medial Forebrain Bundle (MFB) and Anterior Thalamic Radiation (ATR): Imaging of Two Major Subcortical Pathways and the Dynamic Balance of Opposite Affects in Understanding Depression. J. Neuropsychiatry Clin. Neurosci. 2012, 268, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Abbafati, C.; Abbas, K.M.; Abbasi-Kangevari, M.; Abd-Allah, F.; Abdelalim, A.; Abdollahi, M.; Abdollahpour, I.; Abegaz, K.H.; Abolhassani, H.; Aboyans, V.; et al. Global Burden of 369 Diseases and Injuries in 204 Countries and Territories, 1990–2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- Döbrössy, M.D.; Ramanathan, C.; Vajari, D.A.; Tong, Y.; Schlaepfer, T.; Coenen, V.A. Neuromodulation in Psychiatric Disorders: Experimental and Clinical Evidence for Reward and Motivation Network Deep Brain Stimulation: Focus on the Medial Forebrain Bundle. Eur. J. Neurosci. 2021, 53, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Pallikaras, V.; Shizgal, P. The Convergence Model of Brain Reward Circuitry: Implications for Relief of Treatment-Resistant Depression by Deep-Brain Stimulation of the Medial Forebrain Bundle. Front. Behav. Neurosci. 2022, 16, 851067. [Google Scholar] [CrossRef]
- Mineka, S.; Zinbarg, R. Animal Models of Psychopathology. Walker, C.E., Ed.; In Clinical Psychology. Applied Clinical Psychology; Springer: Boston, MA Boston, MA, USA, 1991; pp. 51–86. [Google Scholar] [CrossRef]
- Staay, F.J. van der Animal Models of Behavioral Dysfunctions: Basic Concepts and Classifications, and an Evaluation Strategy. Brain Res. Rev. 2006, 52, 131–159. [Google Scholar] [CrossRef]
- Maximino, C.; Staay, F.J.V.D. Behavioral Models in Psychopathology: Epistemic and Semantic Considerations. Behav. Brain Funct. 2019, 15, 1. [Google Scholar] [CrossRef]
- Pound, P.; Ritskes-Hoitinga, M. Is It Possible to Overcome Issues of External Validity in Preclinical Animal Research? Why Most Animal Models Are Bound to Fail. J. Transl. Med. 2018, 16, 304. [Google Scholar] [CrossRef]
- Lafollette, H.; Shanks, N. Two models of models in biomedical research. Philos. Q. 1995, 45, 141–160. [Google Scholar] [CrossRef]
- Davis, M.T.; Holmes, S.E.; Pietrzak, R.H.; Esterlis, I. Neurobiology of Chronic Stress-Related Psychiatric Disorders: Evidence from Molecular Imaging Studies. Chronic Stress 2017, 1, 29862379. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.V.; Sterlemann, V.; Müller, M.B. Chronic Stress and Individual Vulnerability. Ann. N. Y. Acad. Sci. 2008, 1148, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Fleshner, M.; Maier, S.F.; Lyons, D.M.; Raskind, M.A. The Neurobiology of the Stress-Resistant Brain. Stress 2011, 14, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, I.I.; Gould, T.D. The Endophenotype Concept in Psychiatry: Etymology and Strategic Intentions. Am. J. Psychiatry 2003, 160, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Halahakoon, D.C.; Kieslich, K.; O’Driscoll, C.; Nair, A.; Lewis, G.; Roiser, J.P. Reward-Processing Behavior in Depressed Participants Relative to Healthy Volunteers: A Systematic Review and Meta-Analysis. JAMA Psychiatry 2020, 77, 1286–1295. [Google Scholar] [CrossRef] [PubMed]
- Keren, H.; O’Callaghan, G.; Vidal-Ribas, P.; Buzzell, G.A.; Brotman, M.A.; Leibenluft, E.; Pan, P.M.; Meffert, L.; Kaiser, A.; Wolke, S.; et al. Reward Processing in Depression: A Conceptual and Meta-Analytic Review across FMRI and EEG Studies. Am. J. Psychiatry 2018, 175, 1111–1120. [Google Scholar] [CrossRef]
- Olino, T.M.; McMakin, D.L.; Morgan, J.K.; Silk, J.S.; Birmaher, B.; Axelson, D.A.; Williamson, D.E.; Dahl, R.E.; Ryan, N.D.; Forbes, E.E. Reduced Reward Anticipation in Youth at High-Risk for Unipolar Depression: A Preliminary Study. Dev. Cogn. Neurosci. 2014, 8, 55–64. [Google Scholar] [CrossRef]
- Willner, P. The Validity of Animal Models of Depression. Psychopharmacology 1984, 83, 1–16. [Google Scholar] [CrossRef]
- Lopez-Munoz, F.; Alamo, C. Monoaminergic Neurotransmission: The History of the Discovery of Antidepressants from 1950s Until Today. Curr. Pharm. Des. 2009, 15, 1563–1586. [Google Scholar] [CrossRef]
- Carroll, B.J. Monoamine Precursors in the Treatment of Depression. Clin. Pharmacol. Ther. 1971, 12, 743–761. [Google Scholar] [CrossRef]
- Goodwin, S. Affective Disorders: The Catecholamine Hypothesis Revisited. Psychopharmacol. Bull. 1973, 10, 1157–1164. [Google Scholar] [CrossRef]
- Brown, A.S.; Gershon, S. Dopamine and Depression. J. Neural Transm. Gen. Sect. JNT 1993, 91, 75–109. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, B.W.; Nemeroff, C.B. The Role of Dopamine in the Pathophysiology of Depression. Arch. Gen. Psychiatry 2007, 64, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Hegerl, U.; Hensch, T. Why Do Stimulants Not Work in Typical Depression? Aust. N. Z. J. Psychiatry 2017, 51, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Malhi, G.S.; Byrow, Y.; Bassett, D.; Boyce, P.; Hopwood, M.; Lyndon, W.; Mulder, R.; Porter, R.; Singh, A.; Murray, G. Stimulants for Depression: On the up and Up? Aust. N. Z. J. Psychiatry 2016, 50, 203–207. [Google Scholar] [CrossRef]
- Gnanavel, S.; Sharma, P.; Kaushal, P.; Hussain, S. Attention Deficit Hyperactivity Disorder and Comorbidity: A Review of Literature. World J. Clin. Cases 2019, 7, 2420. [Google Scholar] [CrossRef]
- Mattingly, G.W.; Wilson, J.; Ugarte, L.; Glaser, P. Individualization of Attention-Deficit/Hyperactivity Disorder Treatment: Pharmacotherapy Considerations by Age and Co-Occurring Conditions. CNS Spectr. 2021, 26, 202–221. [Google Scholar] [CrossRef]
- Patkar, A.A.; Masand, P.S.; Pae, C.U.; Peindl, K.; Hooper-Wood, C.; Mannelli, P.; Ciccone, P. A Randomized, Double-Blind, Placebo-Controlled Trial of Augmentation with an Extended Release Formulation of Methylphenidate in Outpatients with Treatment-Resistant Depression. J. Clin. Psychopharmacol. 2006, 26, 653–656. [Google Scholar] [CrossRef]
- Trujillo-Pisanty, I.; Conover, K.; Solis, P.; Palacios, D.; Shizgal, P. Dopamine Neurons Do Not Constitute an Obligatory Stage in the Final Common Path for the Evaluation and Pursuit of Brain Stimulation Reward. PLoS ONE 2020, 15, e0226722. [Google Scholar] [CrossRef]
- Olds, J.; Milner, P. Positive Reinforcement Produced by Electrical Stimulation of Septal Area and Other Regions of Rat Brain. J. Comp. Physiol. Psychol. 1954, 47, 419–427. [Google Scholar] [CrossRef]
- Treadway, M.T.; Buckholtz, J.W.; Schwartzman, A.N.; Lambert, W.E.; Zald, D.H. Worth the “EEfRT”? The Effort Expenditure for Rewards Task as an Objective Measure of Motivation and Anhedonia. PLoS ONE 2009, 4, e6598. [Google Scholar] [CrossRef] [PubMed]
- Treadway, M.T.; Bossaller, N.A.; Shelton, R.C.; Zald, D.H. Effort-Based Decision-Making in Major Depressive Disorder: A Translational Model of Motivational Anhedonia. J. Abnorm. Psychol. 2012, 121, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Gallistel, C.R. Motivating Effects in Self-Stimulation. J. Comp. Physiol. Psychol. 1966, 62, 95–101. [Google Scholar] [CrossRef]
- Edmonds, D.E.; Gallistel, C.R. Parametric Analysis of Brain Stimulation Reward in the Rat: III. Effect of Performance Variables on the Reward Summation Function. J. Comp. Physiol. Psychol. 1974, 87, 876–883. [Google Scholar] [CrossRef]
- Sax, L.; Gallistel, C.R. Characteristics of Spatiotemporal Integration in the Priming and Rewarding Effects of Medial Forebrain Bundle Stimulation. Behav. Neurosci. 1991, 105, 884–900. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, J.A.; Howarth, C.I. Some Tests of a Theory of Intracranial Self-Stimulation. Psychol. Rev. 1963, 70, 444–460. [Google Scholar] [CrossRef]
- Huys, Q.J.M.; Daw, N.D.; Dayan, P. Depression: A Decision-Theoretic Analysis. Annu. Rev. Neurosci. 2015, 38, 1–23. [Google Scholar] [CrossRef]
- Wasserman, E.M.; Gomita, Y.; Gallistel, C.R. Pimozide Blocks Reinforcement but Not Priming from MFB Stimulation in the Rat. Pharm. Biochem Behav 1982, 17, 783–787. [Google Scholar] [CrossRef]
- Coenen, V.; Hurwitz, T.; Panksepp, J.; Mädler, B.; Honey, C. Medial Forebrain Bundle Stimulation Elicits Psychotropic Side Effects in Subthalamic Nucleus Deep Brain Stimulation for PD—New Insights through Diffusion Tensor Imaging. Akt. Neurol. 2009, 36, 749. [Google Scholar] [CrossRef]
- Coenen, V.A.; Schlaepfer, T.E.; Maedler, B.; Panksepp, J. Cross-Species Affective Functions of the Medial Forebrain Bundle—Implications for the Treatment of Affective Pain and Depression in Humans. Neurosci. Biobehav. Rev. 2011, 35, 1971–1981. [Google Scholar] [CrossRef]
- Dahlström, A.; Fuxe, K. A Method for the Demonstration of Monoamine-Containing Nerve Fibres in the Central Nervous System. Acta Physiol. Scand. 1964, 60, 293–294. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Dahlström, A.; Höistad, M.; Marcellino, D.; Jansson, A.; Rivera, A.; Diaz-Cabiale, Z.; Jacobsen, K.; Tinner-Staines, B.; Hagman, B.; et al. From the Golgi–Cajal Mapping to the Transmitter-Based Characterization of the Neuronal Networks Leading to Two Modes of Brain Communication: Wiring and Volume Transmission. Brain Res. Rev. 2007, 55, 17–54. [Google Scholar] [CrossRef] [PubMed]
- Crow, T.J. The Relation between Electrical Self-Stimulation Sites and Catecholamine-Containing Neurons in the Rat Mesencephalon. Experientia 1971, 27, 662. [Google Scholar] [CrossRef]
- German, D.C.; Bowden, D.M. Catecholamine Systems as the Neural Substrate for Intracranial Self-Stimulation: A Hypothesis. Brain Res. 1974, 73, 381–419. [Google Scholar] [CrossRef]
- Wise, R.A. Catecholamine Theories of Reward: A Critical Review. Brain Res. 1978, 152, 215–247. [Google Scholar] [CrossRef]
- Corbett, D.; Wise, R.A. Intracranial Self-Stimulation in Relation to the Ascending Dopaminergic Systems of the Midbrain: A Moveable Electrode Mapping Study. Brain Res. 1980, 185, 1–15. [Google Scholar] [CrossRef]
- Corbett, D.; Wise, R.A. Intracranial Self-Stimulation in Relation to the Ascending Noradrenergic Fiber Systems of the Pontine Tegmentum and Caudal Midbrain: A Moveable Electrode Mapping Study. Brain Res. 1979, 177, 423–436. [Google Scholar] [CrossRef]
- Ungerstedt, U. Stereotaxic Mapping of the Monoamine Pathways in the Rat Brain. Acta Physiol. Scand. Suppl. 1971, 82, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Bielajew, C.; Shizgal, P. Behaviorally Derived Measures of Conduction Velocity in the Substrate for Rewarding Medial Forebrain Bundle Stimulation. Brain Res. 1982, 237, 107–119. [Google Scholar] [CrossRef]
- Bielajew, C.; Shizgal, P. Evidence Implicating Descending Fibers in Self-Stimulation of the Medial Forebrain Bundle. J. Neurosci. 1986, 6, 919–929. [Google Scholar] [CrossRef]
- Gallistel, C.; Shizgal, P.; Yeomans, J. A Portrait of the Substrate for Self-Stimulation. Psychol. Rev. 1981, 88, 228–273. [Google Scholar] [CrossRef] [PubMed]
- Shizgal, P.; Bielajew, C.; Corbett, D.; Skelton, R.; Yeomans, J. Behavioral Methods for Inferring Anatomical Linkage between Rewarding Brain Stimulation Sites. J. Comp. Physiol. Psychol. 1980, 94, 227–237. [Google Scholar] [CrossRef]
- Yeomans, J.S. The Absolute Refractory Periods of Self-Stimulation Neurons. Physiol. Behav. 1979, 22, 911–919. [Google Scholar] [CrossRef]
- Yeomans, J.S. Quantitative Measurement of Neural Post-Stimulation Excitability with Behavioral Methods. Physiol. Behav. 1975, 15, 593–602. [Google Scholar] [CrossRef]
- Anderson, R.M.; Fatigati, M.D.; Rompre, P.-P. Estimates of the Axonal Refractory Period of Midbrain Dopamine Neurons: Their Relevance to Brain Stimulation Reward. Brain Res. 1996, 718, 83–88. [Google Scholar] [CrossRef]
- Feltz, P.; Albe-Fessard, D. A Study of an Ascending Nigro-Caudate Pathway. Electroencephalogr. Clin. Neurophysiol. 1972, 33, 179–193. [Google Scholar] [CrossRef]
- German, D.C.; Dalsass, M.; Kiser, R.S. Electrophysiological Examination of the Ventral Tegmental (A10) Area in the Rat. Brain Res. 1980, 181, 191–197. [Google Scholar] [CrossRef]
- Maeda, H.; Mogenson, G.J. An Electrophysiological Study of Inputs to Neurons of the Ventral Tegmental Area from the Nucleus Accumbens and Medial Preoptic-Anterior Hypothalamic Areas. Brain Res. 1980, 197, 365–377. [Google Scholar] [CrossRef]
- Takigawa, M.; Mogenson, G.J. A Study of Inputs to Antidromically Identified Neurons of the Locus Coeruleus. Brain Res. 1977, 135, 217–230. [Google Scholar] [CrossRef]
- Guyenet, P.G.; Aghajanian, G.K. Antidromic Identification of Dopaminergic and Other Output Neurons of the Rat Substantia Nigra. Brain Res. 1978, 150, 69–84. [Google Scholar] [CrossRef]
- Yim, C.Y.; Mogenson, G.J. Electrophysiological Studies of Neurons in the Ventral Tegmental Area of Tsai. Brain Res. 1980, 181, 301–313. [Google Scholar] [CrossRef]
- Wise, R.A. Action of Drugs of Abuse on Brain Reward Systems. Pharm. Biochem. Behav. 1980, 13 (Suppl. S1), 213–223. [Google Scholar] [CrossRef]
- Yeomans, J.S. Role of Tegmental Cholinergic Neurons in Dopaminergic Activation, Antimuscarinic Psychosis and Schizophrenia. Neuropsychopharmacology 1995, 12, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Yeomans, J.S.; Takeuchi, J.; Baptista, M.; Flynn, D.D.; Lepik, K.; Nobrega, J.; Fulton, J.; Ralph, M.R. Brain-Stimulation Reward Thresholds Raised by an Antisense Oligonucleotide for the M5 Muscarinic Receptor Infused near Dopamine Cells. J. Neurosci 2000, 20, 8861–8867. [Google Scholar] [CrossRef] [PubMed]
- Yetnikoff, L.; Lavezzi, H.N.; Reichard, R.A.; Zahm, D.S. An Update on the Connections of the Ventral Mesencephalic Dopaminergic Complex. Neuroscience 2014, 282, 23–48. [Google Scholar] [CrossRef]
- Matsuda, W.; Furuta, T.; Nakamura, K.C.; Hioki, H.; Fujiyama, F.; Arai, R.; Kaneko, T. Single Nigrostriatal Dopaminergic Neurons Form Widely Spread and Highly Dense Axonal Arborizations in the Neostriatum. J. Neurosci. 2009, 29, 444–453. [Google Scholar] [CrossRef]
- Watabe-Uchida, M.; Zhu, L.; Ogawa, S.K.; Vamanrao, A.; Uchida, N. Whole-Brain Mapping of Direct Inputs to Midbrain Dopamine Neurons. Neuron 2012, 74, 858–873. [Google Scholar] [CrossRef]
- Huston, J.P.; Borbély, A.A. Operant Conditioning in Forebrain Ablated Rats by Use of Rewarding Hypothalamic Stimulation. Brain Res. 1973, 50, 467–472. [Google Scholar] [CrossRef]
- Huston, J.P. Searching for the Neural Mechanism of Reinforcement (of “Stamping In”). In The Neural Basis of Feeding and Reward; Hoebel, B.G., Novin, D., Eds.; Haer Institute for Electrophysiological Research: Brunswick, ME, USA, 1982; pp. 75–83. [Google Scholar]
- Johnson, P.I.; Stellar, J.R. N-Methyl-d-Aspartic Acid-Induced Lesions of the Nucleus Accumbens and/or Ventral Pallidum Fail to Attenuate Lateral Hypothalamic Self-Stimulation Reward. Brain Res. 1994, 646, 73–84. [Google Scholar] [CrossRef]
- Cossette, M.-P.; Conover, K.; Shizgal, P. The Neural Substrates for the Rewarding and Dopamine-Releasing Effects of Medial Forebrain Bundle Stimulation Have Partially Discrepant Frequency Responses. Behav. Brain Res. 2016, 297, 345–358. [Google Scholar] [CrossRef]
- Arvanitogiannis, A.; Shizgal, P. The Reinforcement Mountain: Allocation of Behavior as a Function of the Rate and Intensity of Rewarding Brain Stimulation. Behav. Neurosci. 2008, 122, 1126–1138. [Google Scholar] [CrossRef]
- Breton, Y.-A.; Mullett, A.; Conover, K.; Shizgal, P. Validation and Extension of the Reward-Mountain Model. Front. Behav. Neurosci. 2013, 7, 125. [Google Scholar] [CrossRef]
- Hernandez, G.; Breton, Y.-A.; Conover, K.; Shizgal, P. At What Stage of Neural Processing Does Cocaine Act to Boost Pursuit of Rewards? PLoS ONE 2010, 5, e15081. [Google Scholar] [CrossRef] [PubMed]
- Gallistel, C.R. Self-Stimulation in the Rat: Quantitative Characteristics of the Reward Pathway. J. Comp. Physiol. Psychol. 1978, 92, 977–998. [Google Scholar] [CrossRef]
- Sonnenschein, B.; Conover, K.; Shizgal, P. Growth of Brain Stimulation Reward as a Function of Duration and Stimulation Strength. Behav. Neurosci. 2003, 117, 978–994. [Google Scholar] [CrossRef]
- Hernandez, G.; Trujillo-Pisanty, I.; Cossette, M.-P.; Conover, K.; Shizgal, P. Role of Dopamine Tone in the Pursuit of Brain Stimulation Reward. J. Neurosci 2012, 32, 11032–11041. [Google Scholar] [CrossRef]
- Trujillo-Pisanty, I.; Conover, K.; Shizgal, P. A New View of the Effect of Dopamine Receptor Antagonism on Operant Performance for Rewarding Brain Stimulation in the Rat. Psychopharmacology 2014, 231, 1351–1364. [Google Scholar] [CrossRef]
- Trujillo-Pisanty, I.; Hernandez, G.; Moreau-Debord, I.; Cossette, M.-P.; Conover, K.; Cheer, J.F.; Shizgal, P. Cannabinoid Receptor Blockade Reduces the Opportunity Cost at Which Rats Maintain Operant Performance for Rewarding Brain Stimulation. J. Neurosci. 2011, 31, 5426–5435. [Google Scholar] [CrossRef]
- Nieuwenhuys, R.; Geeraedts, L.M.; Veening, J.G. The Medial Forebrain Bundle of the Rat. I. General Introduction. J. Comp. Neurol. 1982, 206, 49–81. [Google Scholar] [CrossRef]
- Nauta, W.J.; Domesick, V.B. Neural Associations of the Limbic System. In The Neural Basis of Behavior; Beckman, A.L., Ed.; SP Medical & Scientific Books: Tokyo, Japan, 1982; pp. 175–206. ISBN 0-89335-132-6. [Google Scholar]
- Kiss, I.; Shizgal, P. Compound Action Potentials Recorded in the Ventral Tegmental Area, Substantia Nigra, and Periaqueductal Gray Following Rewarding Stimulation of the Lateral Hypothalamus in the Rat. Psychobiology 1990, 18, 205–214. [Google Scholar] [CrossRef]
- Coenen, V.A.; Döbrössy, M.D.; Teo, S.J.; Wessolleck, J.; Sajonz, B.E.A.; Reinacher, P.C.; Thierauf-Emberger, A.; Spittau, B.; Leupold, J.; von Elverfeldt, D.; et al. Diverging Prefrontal Cortex Fiber Connection Routes to the Subthalamic Nucleus and the Mesencephalic Ventral Tegmentum Investigated with Long Range (Normative) and Short Range (Ex-Vivo High Resolution) 7T DTI. Brain Struct. Funct. 2021, 227, 23–47. [Google Scholar] [CrossRef] [PubMed]
- Haber, S.N.; Yendiki, A.; Jbabdi, S. Four Deep Brain Stimulation Targets for Obsessive-Compulsive Disorder: Are They Different? Biol. Psychiatry 2021, 90, 667–677. [Google Scholar] [CrossRef]
- Grisot, G.; Haber, S.N.; Yendiki, A. Diffusion MRI and Anatomic Tracing in the Same Brain Reveal Common Failure Modes of Tractography. NeuroImage 2021, 239, 118300. [Google Scholar] [CrossRef] [PubMed]
- Haber, S.N.; Liu, H.; Seidlitz, J.; Bullmore, E. Prefrontal Connectomics: From Anatomy to Human Imaging. Neuropsychopharmacol. 2022, 47, 20–40. [Google Scholar] [CrossRef]
- Arsenault, M.-Y.; Parent, A.; Séguéla, P.; Descarries, L. Distribution and Morphological Characteristics of Dopamine-Immunoreactive Neurons in the Midbrain of the Squirrel Monkey (Saimiri Sciureus): Dopamine neurons in primate midbrain. J. Comp. Neurol. 1988, 267, 489–506. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pallikaras, V.; Shizgal, P. Dopamine and Beyond: Implications of Psychophysical Studies of Intracranial Self-Stimulation for the Treatment of Depression. Brain Sci. 2022, 12, 1052. https://doi.org/10.3390/brainsci12081052
Pallikaras V, Shizgal P. Dopamine and Beyond: Implications of Psychophysical Studies of Intracranial Self-Stimulation for the Treatment of Depression. Brain Sciences. 2022; 12(8):1052. https://doi.org/10.3390/brainsci12081052
Chicago/Turabian StylePallikaras, Vasilios, and Peter Shizgal. 2022. "Dopamine and Beyond: Implications of Psychophysical Studies of Intracranial Self-Stimulation for the Treatment of Depression" Brain Sciences 12, no. 8: 1052. https://doi.org/10.3390/brainsci12081052
APA StylePallikaras, V., & Shizgal, P. (2022). Dopamine and Beyond: Implications of Psychophysical Studies of Intracranial Self-Stimulation for the Treatment of Depression. Brain Sciences, 12(8), 1052. https://doi.org/10.3390/brainsci12081052