
Dopamine, Erectile Function and Male Sexual Behavior from the Past to the Present: A Review



Abstract
1. Introduction
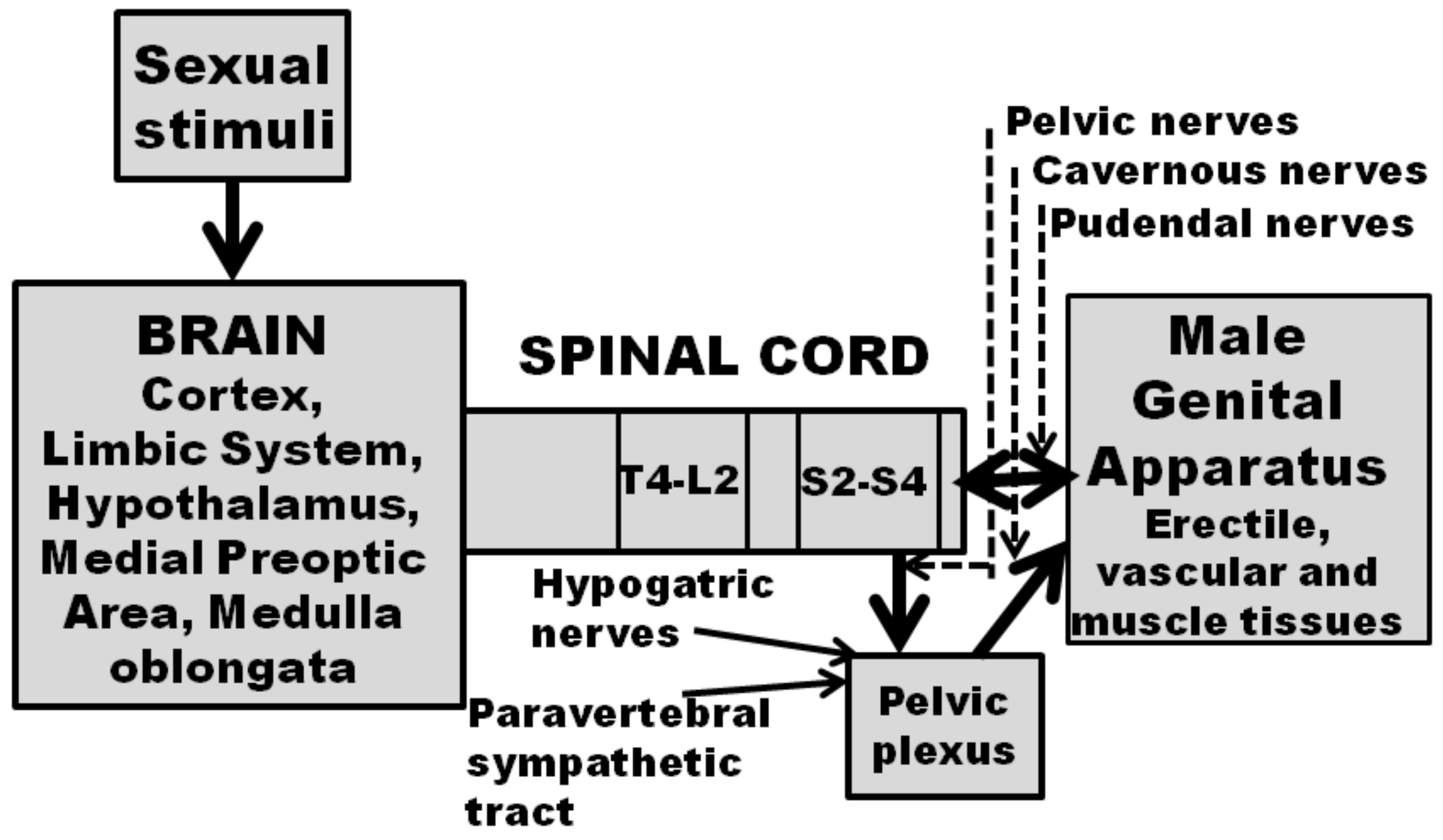
2. Dopamine and Erectile Function
2.1. Dopamine D2-Like Receptor Agonists Given Systemically Induce Penile Erection in Rats, Mice, Monkeys and also in Man
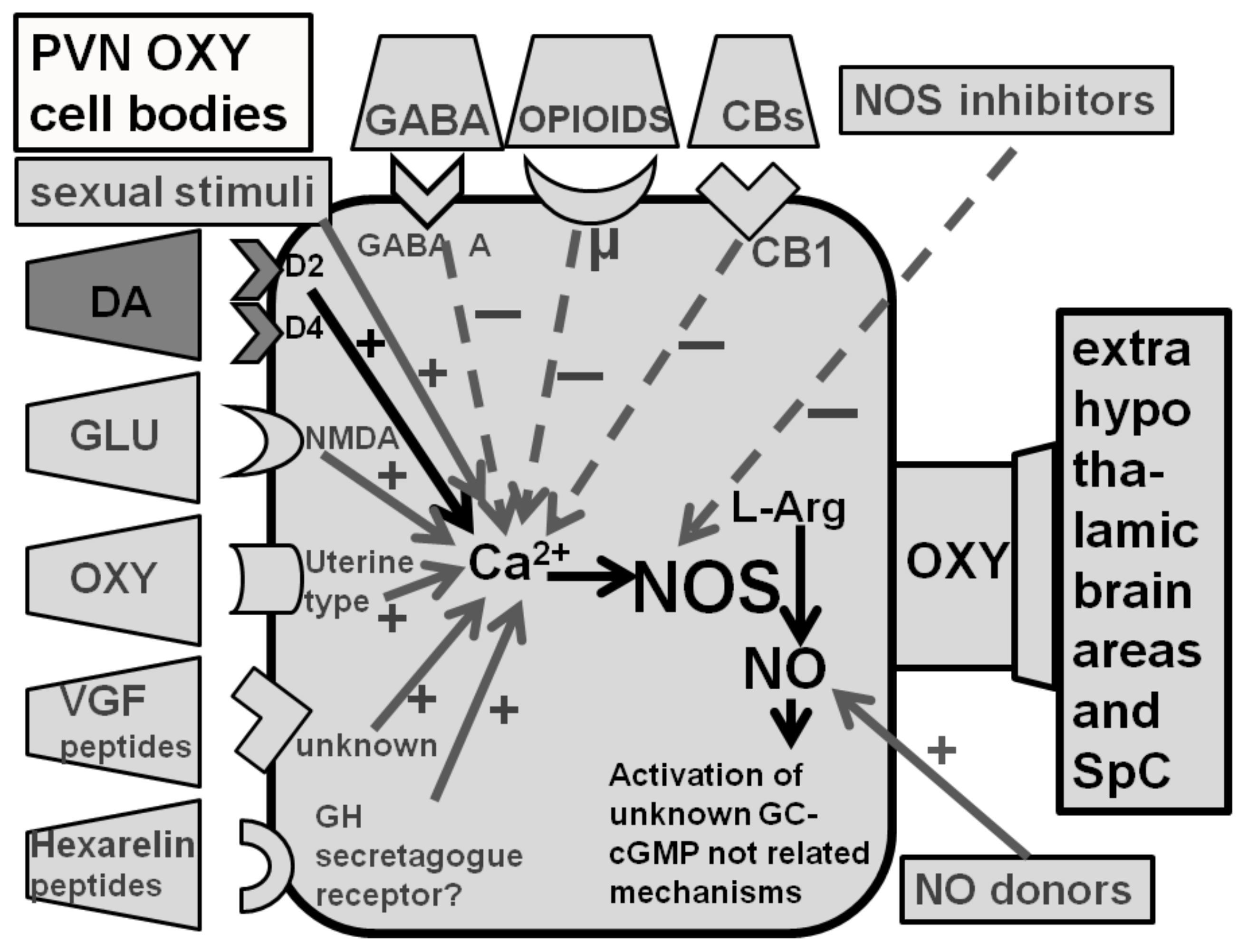
2.2. Dopamine D2-Like Receptor Agonists Induce Penile Erection in Male Rats by Activating PVN Oxytocinergic Neurons Projecting to Extrahypothalamic Brain Areas and the Spinal Cord
2.3. Dopamine Agonists of the D1 and D2 Type Receptors Exert an Opposite Role on Penile Reflexes in Male Rats at the Level of the Medial Preoptic Area and the PVN
2.4. Dopamine D2 and D4, but Not D3 Receptor Subtypes, Mediate Dopamine Agonist-Induced Penile Erection by Activating PVN Oxytocinergic Neurons That Project to Extrahypothalamic Brain Areas and the Spinal Cord
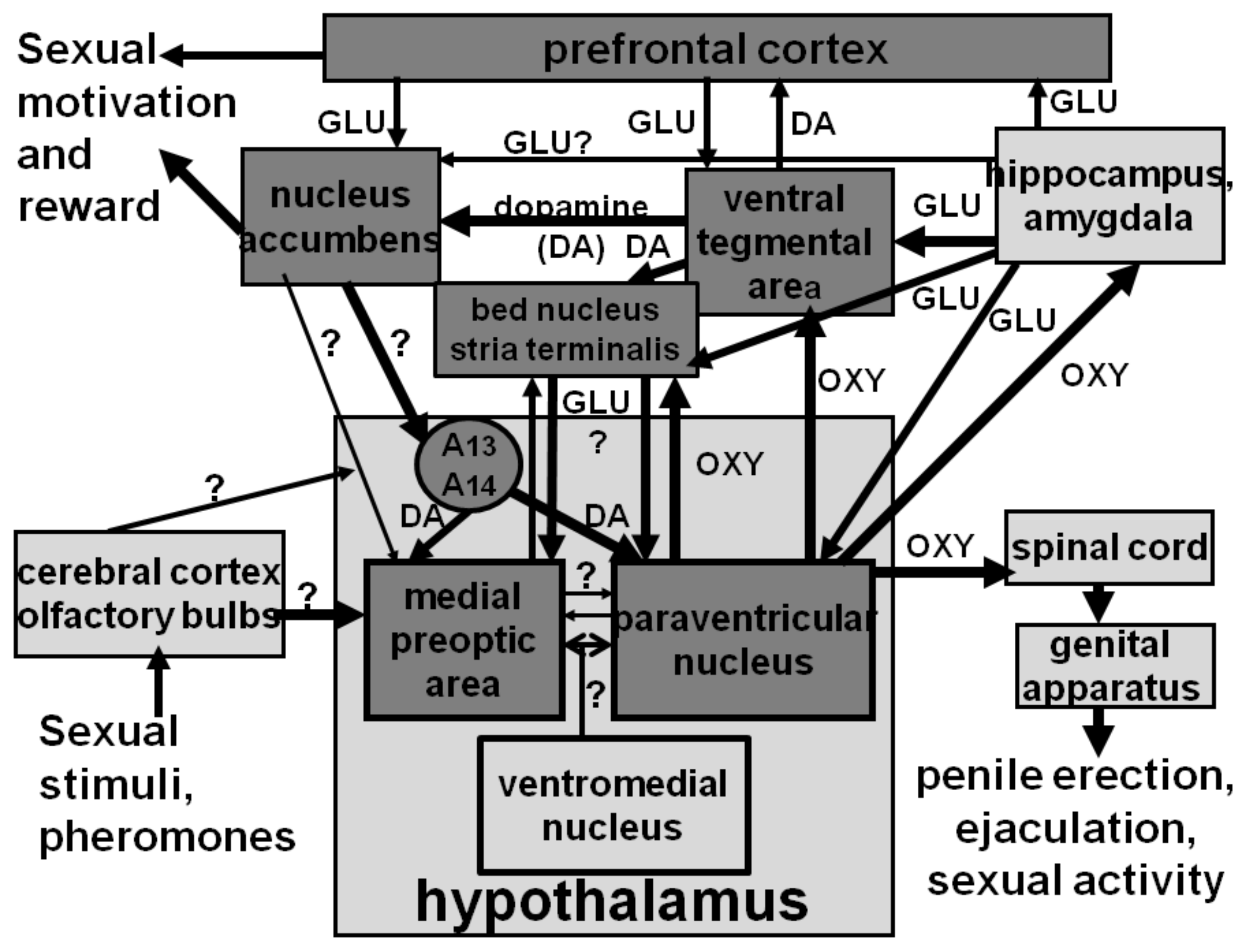
3. Dopamine Released in the Nucleus Accumbens Also Facilitates Penile Erection
4. Dopamine Released in the Bed Nucleus of the Stria Terminalis Also Facilitates Penile Erection
5. Dopamine and Sexual Behavior
5.1. Dopamine and Male Sexual Behavior
5.2. Dopamine Receptors and Male Sexual Behavior
5.3. D4 Dopamine Receptors Agonists and Antagonists Improve and Impair Sexual Behavior in Male Rats: Comparison with Mixed D2-Like Dopamine Receptor Agonists and Antagonists
5.4. Differences between Mixed Dopamine D2-like Receptor Agonists and D4 Receptor Agonists on Penile Erection and Copulatory Behavior
6. Rats with a Higher Dopaminergic Tone Than Their Control Counterparts Obtained by Selective Breeding Protocols over Multiple Generations or by Genetic Mutations Show Improved Erectile Function and Copulatory Behavior
6.1. Erectile Function and Sexual Behavior in Rats Selectively Bred That Show a Higher Dopaminergic Tone than Their Lower Tone Counterparts
6.2. RHA and RLA Rats Have a Different Central Dopaminergic Tone
6.3. RHA and RLA Rats, Erectile Function and Dopamine
6.4. RHA and RLA rats, Sexual Behavior and Dopamine
6.5. Experimental Evidence Supporting the Role of a Different Dopaminergic Tone in the Sexual Differences between RHA and RLA Rats
6.5.1. Autoradiography, Binding and Behavioral Studies
6.5.2. Microdialysis and Copulatory Behavior Combined Experiments
6.6. BNEHR and BNELR Rats, Sexual Behavior and Dopamine
6.7. LY and HY Rats, Erectile Function, Sexual Behavior and Dopamine
6.8. Erectile Function and Sexual Behavior in Rats with a Genetically-Induced Increase in Dopaminergic Tone
6.9. DAT-KO, DAT-HET and DAT-WT Rats, Sexual Behavior and Dopamine
7. Dopaminergic Systems, Sexual Motivation, Reward and Performance
8. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Blaschko, H. Catecholamines 1922–1971. In Catcholamines. Handbuch der Experimentellen Pharmakologie; Blaschko, H., Muscholl, E., Eds.; Springer: Berlin, Germany, 1972; Volume 33, pp. 1–15. ISBN 0-387-05517-7. [Google Scholar]
- Blaschko, H. A half-century of research (1922–1972) on catecholamine biosynthesis. J. App. Cardiol. 1987, 2, 171–183. [Google Scholar]
- Axelrod, J. Purification and properties of phenylethanolamine-N-methyl transferase. J. Biol. Chem. 1962, 237, 1657–1660. [Google Scholar] [CrossRef]
- Axelrod, J. O-Methylation of epinephrine and other catechols. Science 1962, 126, 400–401. [Google Scholar] [CrossRef] [PubMed]
- Stjärne, L.; Hedqvist, P.; Lagercrantz, H.; Wennmalm, Å. (Eds.) Chemical Neurotransmission 75 Years; Academic Press: London, UK, 1981; p. 13. [Google Scholar]
- Eisenhofer, G.; Kopin, I.J.; Goldstein, D.S. Catecholamine metabolism: A contemporary view with implications for physiology and medicine. Pharmacol. Rev. 2004, 56, 331–349. [Google Scholar] [CrossRef]
- Donnerer, J.; Lembeck, F. Adrenaline, noradrenaline and dopamine: The catecholamines. In The Chemical Languages of the Nervous System; Karger: Basel, Switzerland, 2006; pp. 150–160. [Google Scholar]
- Carlsson, A. The occurrence, distribution and physiological role of catcholamines in the nervous system. Pharmacol. Rev. 1959, 11, 490–493. [Google Scholar]
- Carlsson, A. Thirthy years of dopamine research. Adv. Neurol. 1993, 60, 1–10. [Google Scholar]
- Bertler, Å.; Rosengren, E. Occurrence and distribution of dopamine in brain and other tissues. Experientia 1959, 15, 10–11. [Google Scholar] [CrossRef]
- Dahlstroem, A.; Fuxe, K. Evidence for the existence of monoamine neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain stem neurons. Acta Physiol. Scand. Suppl. 1964, 62, 1–55. [Google Scholar]
- Kebabian, J.W.; Calne, D.B. Multiple receptors for dopamine. Nature 1979, 277, 93–96. [Google Scholar] [CrossRef]
- Seeman, P.; Van Tol, H.H. Dopamine receptor pharmacology. Trends Pharmacol. Sci. 1994, 15, 264–270. [Google Scholar] [CrossRef]
- Sokoloff, P.; Schwartz, J.-C. Novel dopamine receptors half a decade later. Trends Pharmacol. Sci. 1995, 16, 270–275. [Google Scholar] [CrossRef]
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef] [PubMed]
- Martel, J.C.; McArthur, S.G. Dopamine receptor subtypes, physiology and pharmacology: New ligands and concepts in schizophrenia. Front. Pharmacol. 2020, 11, 1003. [Google Scholar] [CrossRef]
- Hieble, J.P. Adrenergic Receptors. In Encyclopedia of Neuroscience; Squire, L.R., Ed.; Academic Press: Oxford, MS, USA, 2009; pp. 135–139. [Google Scholar]
- Qu, L.; Zhou, Q.; Xu, Y.; Guo, Y.; Chen, X.; Yao, D.; Han, G.W.; Liu, Z.-J.; Stevens, R.C.; Zhong, G.; et al. Structural basis of the diversity of adrenergic receptors. Cell Rep. 2019, 29, 2929–2935. [Google Scholar] [CrossRef]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef]
- Aghajanian, G.K.; Sanders-Bush, E. Serotonin. In Psychopharmacology: The Fifth Generation of Progress; Davis, K.L., Charney, D., Coyle, J.T., Nemeroff, C., Eds.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2002; pp. 15–35. [Google Scholar]
- Iversen, S.D.; Iversen, L.L. Dopamine: 50 years in perspective. Trends Neurosci. 2007, 30, 188–193. [Google Scholar] [CrossRef]
- Ehringer, H.; Hornykiewicz, O. Verteilung von Noradrenalin und Dopamin (3-Hydroxytyramin) im Gehirn des Menschen und ihr Verhalten bei Erkrankungen des Extrapyramidalen Systems. Klin. Wochenschr. 1960, 38, 1236–1239. [Google Scholar] [CrossRef]
- Cotzias, G.C.; Van Woert, M.H.; Schiffer, L.M. Aromatic amino acids and modification of parkinsonism. N. Engl. J. Med. 1967, 276, 374–379. [Google Scholar] [CrossRef]
- Hornykiewicz, O. From dopamine to Parkinson’s disease: A personal research record. In The Neurosciences: Paths of Discovery II; Samson, F., Adelman, G., Eds.; Birkhäuser: Basel, Switzerland, 1992; pp. 125–147. [Google Scholar]
- Wise, R.A.; Robble, M.A. Dopamine and Addiction. Annu. Rev. Psychol. 2020, 71, 79–106. [Google Scholar] [CrossRef]
- Wise, R.A.; Jordan, C.J. Dopamine, behavior, and addiction. J. Biomed. Sci. 2021, 28, 83. [Google Scholar] [CrossRef]
- Seeman, P.; Lee, T.; Chau-Wong, M.; Wong, K. Antipsychotic drug doses and neuroleptic/dopamine receptors. Nature 1976, 261, 717–719. [Google Scholar] [CrossRef] [PubMed]
- Seeman, M.V. History of the dopamine hypothesis of antipsychotic action. World J. Psychiatry 2021, 11, 355–364. [Google Scholar] [CrossRef] [PubMed]
- McCann, S.M.; Ojeda, S.R.; Fawcett, C.P.; Krulich, L. Catecholaminergic control of gonadotropin and prolactin secretion with particular reference to the possible participation of dopamine. Adv. Neurol. 1974, 5, 435–445. [Google Scholar]
- MacLeod, R.M.; Login, I. Control of prolactin secretion by the hypothalamic catecholamines. Adv. Sex Horm. Res. 1976, 2, 211–231. [Google Scholar] [PubMed]
- Flückiger, E. Dopaminergic control of prolactin secretion. Bull. Schweiz Akad. Med. Wiss. 1978, 34, 191–196. [Google Scholar] [PubMed]
- Soulairac, A.; Soulairac, M.L. Action de l’amphetamine, de l’adrenaline et de l’atropine sur le comportement sexuel du rat male. J. Physiol. (Paris) 1957, 49, 381–385. [Google Scholar]
- Tagliamonte, A.; Tagliamonte, P.; Gessa, G.L.; Brodie, B.B. Compulsive sexual activity induced by p-chlorophenylalanine in normal and pinealectomized male rats. Science 1969, 166, 1433–1435. [Google Scholar] [CrossRef]
- Tagliamonte, A.; Tagliamonte, P.; Gessa, G.L. Reversal of pargyline-induced inhibition of sexual behaviour in male rats by p-chlorophenylalanine. Nature 1971, 230, 244–245. [Google Scholar] [CrossRef]
- Tagliamonte, A.; Fratta, W.; Del Fiacco, M.; Gessa, G. Possible stimulatory role of brain dopamine in the copulatory behavior of male rats. Pharmacol. Biochem. Behav. 1974, 2, 257–260. [Google Scholar] [CrossRef]
- Tagliamonte, A.; Fratta, W.; Gessa, G.L. Aphrodisiac effect ofl-DOPA and apomorphine in male sexually sluggish rats. Exp. 1974, 30, 381–382. [Google Scholar] [CrossRef]
- Da Prada, M.; Carruba, M.; Saner, A.; O’Brien, R.; Pletscher, A. The action ofl-DOPA on sexual behaviour of male rats. Brain Res. 1973, 55, 383–389. [Google Scholar] [CrossRef]
- Gessa, G.L.; Tagliamonte, A. Role of brain monoamines in male sexual behavior. Life Sci. 1974, 14, 425–436. [Google Scholar] [CrossRef]
- Gray, G.D.; Davis, H.N.; Dewsbury, D.A. Effects of L-DOPA on the heterosexual copulatory behavior of male rats. Eur. J. Pharmacol. 1974, 27, 367–370. [Google Scholar] [CrossRef]
- Hliňák, Z.; Madlafousek, J.; Krejčí, I. Effects of lisuride on precopulatory and copulatory behaviour of adult male rats. Psychopharmacology 1983, 79, 231–235. [Google Scholar] [CrossRef]
- Meyerson, B.J. The effect of neuropharmacological agents on hormone-activated estrous behavior in ovariectomized rats. Arch. Int. Pharmacodyn. Ther. 1964, 160, 4–33. [Google Scholar]
- Ahlenius, S.; Engel, J.; Eriksson, H.; Modigh, K. Importance of central catecholamines in the mediation of lordosis behaviour in ovariectomized rats treated with estrogen and inhibitors of monoamine synthesis. J. Neural Transm. 1972, 33, 247–255. [Google Scholar] [CrossRef]
- Everitt, B.J.; Fuxe, K.; Hökfelt, T. Inhibitory role of dopamine and 5-hydroxytryptamine in the sexual behaviour of female rats. Eur. J. Pharmacol. 1974, 29, 187–191. [Google Scholar] [CrossRef]
- Everitt, B.J.; Fuxe, K. Dopamine and sexual behaviour in female rats. Effects of dopamine receptor agonists and sulpiride. Neurosci. Lett. 1977, 4, 209–213. [Google Scholar] [CrossRef]
- Tsuruta, K.; Frey, E.A.; Grewe, C.W.; Cote, T.E.; Eskay, R.L.; Kebabian, J.W. Evidence that LY 171555 specifically stimulates the D-2 dopamine receptor. Nature 1981, 292, 463–465. [Google Scholar] [CrossRef]
- Melis, M.R.; Argiolas, A.; Gessa, G.L. Apomorphine-induced penile erection and yawning: Site of action in brain. Brain Res. 1987, 415, 98–102. [Google Scholar] [CrossRef]
- Hull, E.; Bitran, D.; Pehek, E.A.; Warner, R.K.; Band, L.C.; Holmes, G.M. Dopaminergic control of male sex behavior in rats: Effects of an intracerebrally-infused agonist. Brain Res. 1986, 370, 73–81. [Google Scholar] [CrossRef]
- Hull, E.M.; Eaton, R.C.; Markowski, V.P.; Moses, J.; Lumley, L.A.; Loucks, J.A. Opposite influence of medial preoptic D1 and D2 receptors on genital reflexes: Implications for copulation. Life Sci. 1992, 51, 1705–1713. [Google Scholar] [CrossRef]
- Hull, E.M.; Warner, R.K.; Bazzett, T.J.; Eaton, R.C.; Thompson, J.T.; Scaletta, L.L. D2/D1 ratio in the medial preoptic area affects copulation of male rats. J. Pharmacol. Exp. Ther. 1989, 251, 422–427. [Google Scholar] [PubMed]
- Markowski, V.P.; Eaton, R.C.; Lumley, L.A.; Moses, J.; Hull, E.M. A D1 agonist in the MPOA facilitates copulation in male rats. Pharmacol. Biochem. Behav. 1994, 47, 483–486. [Google Scholar] [CrossRef][Green Version]
- Collins, G.T.; Truccone, A.; Haji-Abdi, F.; Newman, A.H.; Grundt, P.; Rice, K.C.; Husbands, S.M.; Greedy, B.M.; Enguehard-Gueiffier, C.; Gueiffier, A.; et al. Proerectile effects of dopamine D2-like agonists are mediated by the D3 receptor in rats and mice. J. Pharmacol. Exp. Ther. 2009, 329, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Depoortère, R.; Bardin, L.; Rodrigues, M.; Abrial, E.; Aliaga, M.; Newman-Tancredi, A. Penile erection and yawning induced by dopamine D2-like receptor agonists in rats: Influence of strain and contribution of dopamine D2, but not D3 and D4 receptors. Behav. Pharmacol. 2009, 20, 303–311. [Google Scholar] [CrossRef]
- Ferrari, F.; Giuliani, D. Behavioral effects induced by the dopamine D3 agonist 7-OH-DPAT in sexually-active and -inactive male rats. Neuropharmacology 1996, 35, 279–284. [Google Scholar] [CrossRef]
- Clément, P.; Pozzato, C.; Heidbreder, C.; Alexandre, L.; Giuliano, F.; Melotto, S. Delay of ejaculation induced by SB-277011, a selective dopamine D3 receptor antagonist, in the rat. J. Sex. Med. 2009, 6, 980–988. [Google Scholar] [CrossRef]
- Hsieh, G.C.; Hollingsworth, P.R.; Martino, B.; Chang, R.; Terranova, M.A.; O’Neill, A.B.; Lynch, J.J.; Moreland, R.B.; Donnelly-Roberts, D.L.; Kolasa, T.; et al. Central mechanisms regulating penile erection in conscious rats: The dopaminergic systems related to the proerectile effect of apomorphine. J. Pharmacol. Exp. Ther. 2004, 308, 330–338. [Google Scholar] [CrossRef]
- Mierau, J.; Schneider, F.J.; Ensinger, H.A.; Chio, C.L.; Lajiness, M.E.; Huff, R.M. Pramipexole binding and activation of cloned and expressed dopamine D2, D3 and D4 receptors. Eur. J. Pharmacol. Mol. Pharmacol. 1995, 290, 29–36. [Google Scholar] [CrossRef]
- Glase, S.A.; Akunne, H.C.; Georgic, L.M.; Heffner, T.G.; MacKenzie, R.G.; Manley, P.J.; Pugsley, A.T.A.; Wise, L.D. Substituted [(4-Phenylpiperazinyl)-methyl] benzamides: Selective dopamine D4 agonists. J. Med. Chem. 1997, 40, 1771–1772. [Google Scholar] [CrossRef] [PubMed]
- Brioni, J.D.; Moreland, R.B.; Cowart, M.; Hsieh, G.C.; Stewart, A.O.; Hedlund, P.; Donnelly-Roberts, D.L.; Nakane, M.; Lynch, J.J., III; Kolasa, T.; et al. Activation of dopamine D4 receptors by ABT-724 induces penile erection in rats. Proc. Natl. Acad. Sci. USA 2004, 101, 6758–6763. [Google Scholar] [CrossRef] [PubMed]
- Enguehard-Gueiffier, C.; Hübner, H.; El Hakmaoui, A.; Allouchi, H.; Gmeiner, P.; Argiolas, A.; Melis, M.R.; Gueiffier, A. 2-[(4-phenylpiperazin-1-yl)methyl]imidazo(di)azines as selective D4-ligands. induction of penile erection by 2-[4-(2-methoxyphenyl)piperazin-1-ylmethyl]imidazo[1,2-a]pyridine (PIP3EA), a potent and selective D4 partial agonist. J. Med. Chem. 2006, 49, 3938–3947. [Google Scholar] [CrossRef] [PubMed]
- Löber, S.; Tschammer, N.; Hübner, H.; Melis, M.R.; Argiolas, A.; Gmeiner, P. the azulene framework as a novel arene bioisostere: Design of potent dopamine D4 receptor ligands inducing penile erection. Chem. Med. Chem. 2009, 4, 325–328. [Google Scholar] [CrossRef]
- Löber, S.; Hübner, H.; Buschauer, A.; Sanna, F.; Argiolas, A.; Melis, M.R.; Gmeiner, P. Novel azulene derivatives for the treatment of erectile dysfunction. Bioorganic Med. Chem. Lett. 2012, 22, 7151–7154. [Google Scholar] [CrossRef]
- Moreland, R.B.; Patel, M.; Hsieh, G.C.; Wetter, J.M.; Marsh, K.; Brioni, J.D. A-412997 is a selective dopamine D4 receptor agonist in rats. Pharmacol. Biochem. Behav. 2005, 82, 140–147. [Google Scholar] [CrossRef]
- Zorn, S.H.; Jackson, E.; Johnon, C.; Lewis, L.; Fliri, A. CP-226269 is a selective dopamine D4 agonist. Soc. Neurosci. Abstr. 1997, 23, 685. [Google Scholar]
- Kulagowski, J.J.; Broughton, H.B.; Curtis, N.R.; Mawer, I.M.; Ridgill, M.P.; Baker, R.; Emms, F.; Freedman, S.B.; Marwood, R.; Patel, S.; et al. 3-[[4-(4-chlorophenyl)piperazin-1-yl]methyl]-1H-pyrrolo[2,3-b]pyridine: An antagonist with high affinity and selectivity for the human dopamine D4 receptor. J. Med. Chem. 1996, 39, 1941–1942. [Google Scholar] [CrossRef]
- Reavill, C.; Taylor, S.G.; Wood, M.D.; Ashmeade, T.; Austin, N.E.; Avenell, K.Y.; Boyfield, I.; Branch, C.L.; Cilia, J.; Coldwell, M.C.; et al. Pharmacological actions of a novel, high-affinity, and selective human dopamine D3 receptor antagonist, SB-277011A. J. Pharmacol. Exp. Ther. 2000, 294, 1154–1165. [Google Scholar]
- Bettinetti, L.; Schlotter, K.; Hübner, H.; Gmeiner, P. Interactive SAR studies: Rational discovery of super-potent and highly selective dopamine D3 receptor antagonists and partial agonists. J. Med. Chem. 2002, 45, 4594–4597. [Google Scholar] [CrossRef]
- Tice, M.; Hashemi, T.; Taylor, L.; Duffy, R.; McQuade, R. Characterization of the binding of SCH 39166 to the five cloned dopamine receptor subtypes. Pharmacol. Biochem. Behav. 1994, 49, 567–571. [Google Scholar] [CrossRef]
- Millan, M.J.; Newman-Tancredi, A.; Brocco, M.; Gobert, A.; Lejeune, F.; Audinot, V.; Rivet, J.M.; Schreiber, R.; Dekeyne, A.; Spedding, M.; et al. S 18126 ([2-[4-(2,3-dihydrobenzo[1,4]dioxin-6-yl)piperazin-1-yl methyl]indan-2-yl]), a potent, selective and competitive antagonist at dopamine D4 receptors: An in vitro and in vivo comparison with L 745,870 (3-(4-[4-chlorophenyl]piperazin-1-yl)methyl-1H-pyrrolo[2, 3b]pyridine) and raclopride. J. Pharmacol. Exp. Ther. 1998, 287, 167–186. [Google Scholar] [PubMed]
- Melis, M.R.; Argiolas, A. Dopamine and sexual behavior. Neurosci. Biobehav. Rev. 1995, 19, 19–38. [Google Scholar] [CrossRef]
- Sanna, F.; Contini, A.; Melis, M.R.; Argiolas, A. Role of dopamine D4 receptors in copulatory behavior: Studies with selective D4 agonists and antagonists. Pharmacol. Biochem. Behav. 2015, 137, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Cagiano, R.; Barfield, R.J.; White, N.R.; Pleim, E.T.; Cuomo, V. Mediation of rat postejaculatory 22 kHz ultrasonic vocalization by dopamine D2 receptors. Pharmacol. Biochem. Behav. 1989, 34, 53–58. [Google Scholar] [CrossRef]
- Beck, J.; Biały, M.; Kostowski, W. Effects of D1 receptor agonist SKF 38393 on male rat sexual behavior and postcopulatory departure in the goal compartment–runway paradigm. Physiol. Behav. 2002, 76, 91–97. [Google Scholar] [CrossRef]
- Guadarrama-Bazante, I.L.; Canseco-Alba, A.; Rodríguez-Manzo, G. Dopamine receptors play distinct roles in sexual behavior expression of rats with a different sexual motivational tone. Behav. Pharmacol. 2014, 25, 684–694. [Google Scholar] [CrossRef]
- Sanna, F.; Succu, S.; Hübner, H.; Gmeiner, P.; Argiolas, A.; Melis, M.R. Dopamine D2-like receptor agonists induce penile erection in male rats: Differential role of D2, D3 and D4 receptors in the paraventricular nucleus of the hypothalamus. Behav. Brain Res. 2011, 225, 169–176. [Google Scholar] [CrossRef]
- Sanna, F.; Succu, S.; Melis, M.R.; Argiolas, A. Dopamine agonist-induced penile erection and yawning: Differential role of D2 receptor subtypes and correlation with nitric oxide production in the paraventricular nucleus of the hypothalamus of male rats. Behav. Brain Res. 2012, 230, 355–364. [Google Scholar] [CrossRef]
- Ferrari, F.; Pelloni, F.; Giuliani, D. Behavioural evidence that different neurochemical mechanisms underly stretching-yawning and penile erection induced in male rats by SND 919, a new selective D2 dopamine receptor agonist. Psychopharmacology 1993, 113, 172–176. [Google Scholar] [CrossRef]
- Ferrari, F.; Giuliani, D. Sexual attraction and copulation in male rats: Effects of the dopamine agonist SND 919. Pharmacol. Biochem. Behav. 1995, 50, 29–34. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Mascia, M.S.; Argiolas, A. PD-168,077, a selective dopamine D4 receptor agonist, induces penile erection when injected into the paraventricular nucleus of male rats. Neurosci. Lett. 2005, 379, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Melis, M.R.; Succu, S.; Sanna, F.; Melis, T.; Mascia, M.S.; Enguehard-Gueiffier, C.; Hübner, H.; Gmeiner, P.; Gueiffier, A.; Argiolas, A. PIP3EA and PD168077, two selective dopamine D4 receptor agonists, induce penile erection in male rats: Site and mechanism of action in the brain. Eur. J. Neurosci. 2006, 24, 2021–2030. [Google Scholar] [CrossRef]
- Succu, S.; Sanna, F.; Melis, T.; Boi, A.; Argiolas, A.; Melis, M.R. Stimulation of dopamine receptors in the paraventricular nucleus of the hypothalamus of male rats induces penile erection and increases extra-cellular dopamine in the nucleus accumbens: Involvement of central oxytocin. Neuropharmacology 2007, 52, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Corda, M.G.; Melis, M.R.; Piludu, M.A.; Löber, S.; Hübner, H.; Gmeiner, P.; Argiolas, A.; Giorgi, O. Dopamine agonist-induced penile erection and yawning: A comparative study in outbred Roman high- and low-avoidance rats. Pharmacol. Biochem. Behav. 2013, 109, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Argiolas, A.; Melis, M.R. Neuromodulation of penile erection: An overview of the role of neurotransmitters and neuropeptides. Prog. Neurobiol. 1995, 47, 235–255. [Google Scholar] [CrossRef]
- Argiolas, A. Male erectile dysfunction: Chemical pharmacology of penile erection. Drug Discov. Today Ther. Strat. 2005, 2, 31–36. [Google Scholar] [CrossRef]
- Andersson, K.E.; Wagner, G. Physiology of penile erection. Physiol. Rev. 1995, 75, 191–236. [Google Scholar] [CrossRef]
- McKenna, K.E. Central control of penile erection. Int. J. Impot. Res. 1998, 10, S25–S34. [Google Scholar]
- Sato, Y.; Christ, G.J.; Horita, H.; Adachi, H.; Suzuki, N.; Tsukamoto, T. The effects of alterations in nitric oxide levels in the paraventricular nucleus on copulatory behavior and reflexive erections in male rats. J. Urol. 1999, 162, 2182–2185. [Google Scholar] [CrossRef]
- Meisel, R.L.; Sachs, B.D. The physiology of male sexual behavior. In The Physiology of Reproduction; Knobil, E., Neil, J., Eds.; Raven Press: New York, NY, USA, 1994; pp. 3–96. [Google Scholar]
- McKenna, K. Some proposals regarding the organization of the central nervous system control of penile erection. Neurosci. Biobehav. Rev. 2000, 24, 535–540. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.R. The neuropharmacology of yawning. Eur. J. Pharmacol. 1998, 343, 1–16. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Mauri, A.; Argiolas, A. Nitric oxide production is increased in the paraventricular nucleus of the hypothalamus of male rats during non-contact penile erections and copulation. Eur. J. Neurosci. 1998, 10, 1968–1974. [Google Scholar] [CrossRef] [PubMed]
- Argiolas, A. Nitric oxide is a central mediator of penile erection. Neuropharmacology 1994, 33, 1339–1344. [Google Scholar] [CrossRef]
- Sachs, B.D. Contextual approaches to the physiology and classification of erectile function, erectile dysfunction, and sexual arousal. Neurosci. Biobehav. Rev. 2000, 24, 541–560. [Google Scholar] [CrossRef]
- Giuliano, F.; Rampin, O. Central neural regulation of penile erection. Neurosci. Biobehav. Rev. 2000, 24, 517–533. [Google Scholar] [CrossRef]
- Chitaley, K.; Wingard, C.; Webb, R.C.; Branam, H.; Stopper, V.S.; Lewis, R.W.; Mills, T.M. Antagonism of Rho-kinase stimulates rat penile erection via a nitric oxide-independent pathway. Nat. Med. 2001, 7, 119–122. [Google Scholar] [CrossRef]
- Giuliano, F.; Allard, J. Dopamine and sexual function. Int. J. Impot. Res. 2001, 13, S18–S28. [Google Scholar] [CrossRef]
- Melis, M.R.; Argiolas, A. Central oxytocinergic neurotransmission: A drug target for the therapy of psychogenic erectile dysfunction. Curr. Drug Targets 2003, 4, 55–66. [Google Scholar] [CrossRef]
- Heaton, J.P. Central neuropharmacological agents and mechanisms in erectile dysfunction: The role of dopamine. Neurosci. Biobehav. Rev. 2000, 24, 561–569. [Google Scholar] [CrossRef]
- Giuliano, F.; Rampin, O. Neural control of erection. Physiol. Behav. 2004, 83, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.-E. Mechanisms of penile erection and basis for pharmacological treatment of erectile dysfunction. Pharmacol. Rev. 2011, 63, 811–859. [Google Scholar] [CrossRef] [PubMed]
- Argiolas, A.; Melis, M.R. Neuropeptides and central control of sexual behaviour from the past to the present: A review. Prog. Neurobiol. 2013, 108, 80–107. [Google Scholar] [CrossRef] [PubMed]
- Melis, M.R.; Argiolas, A. Oxytocin, erectile function and sexual behavior: Last discoveries and possible advances. Int. J. Mol. Sci. 2021, 22, 10376. [Google Scholar] [CrossRef]
- Melis, M.R.; Argiolas, A. Erectile function and sexual behavior: A review of the role of nitric oxide in the central nervous system. Biomolecules 2021, 11, 1866. [Google Scholar] [CrossRef]
- Mogilnicka, E.; Klimek, V. Drugs affecting dopamine neurons and yawning behavior. Pharmacol. Biochem. Behav. 1977, 7, 303–305. [Google Scholar] [CrossRef]
- Benassi-Benelli, A.; Ferrari, F.; Pellegrino-Quarantotti, B. Penile erections induced by apomorphine and N-propyl-norapomorphine in rats. Arch. Int. Pharmacodyn. Ther. 1979, 242, 241–247. [Google Scholar]
- Bertolini, A.; Genedani, S.; Castelli, M. Behavioural effects of naloxone in rats. Experientia 1978, 34, 771–772. [Google Scholar] [CrossRef]
- Baggio, G.; Ferrari, F. The role of dopaminergic receptors in the behavioral effects induced by lisuride in male rats. Psychopharmacology 1983, 80, 38–42. [Google Scholar] [CrossRef]
- Gower, A.J.; Berendsen, H.H.G.; Princen, N.N.; Broekkamp, C.L. Antagonism of drug-induced yawning and penile erection in rats. Eur. J. Pharmacol. 1984, 122, 239–244. [Google Scholar] [CrossRef]
- Lal, S. Apomorphine in the evaluation of dopaminergic function in man. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1988, 12, 117–164. [Google Scholar] [CrossRef]
- Lal, S.; Tesfaye, Y.; Thavundayil, J.X.; Thompson, T.R.; Kiely, M.E.; Nair, N.; Grassino, A.; Dubrovsky, B. Apomorphine: Clinical studies on erectile impotence and yawning. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1989, 13, 329–339. [Google Scholar] [CrossRef]
- Pomerantz, S.M. Apomorphine facilities male sexual behavior of rhesus monkeys. Pharmacol. Biochem. Behav. 1990, 35, 659–664. [Google Scholar] [CrossRef]
- Pomerantz, S.M. Quinelorane (LY 163502), a D2 dopamine receptor agonist, acts centrally to facilitate penile erections of male rhesus monkeys. Pharmacol. Biochem. Behav. 1991, 39, 123–128. [Google Scholar] [CrossRef]
- Lal, S.; Kiely, M.E.; Thavundayil, J.X.; Stewart, J.D.; Assalian, P.; Ackman, C.F.D. Effect of bromocriptine in patients with apomorphine-responsive erectile impotence: An open study. J. Psychiatry Neurosci. 1991, 16, 262–266. [Google Scholar]
- Pomerantz, S.M. Dopaminergic influences on male sexual behavior of rhesus monkeys: Effects of dopamine agonists. Pharmacol. Biochem. Behav. 1992, 41, 511–517. [Google Scholar] [CrossRef]
- Laduron, P.M.; Leysen, J.E. Domperidone, a specific in vitro dopamine antagonist, devoid of in vivo central dopaminergic activity. Biochem. Pharmacol. 1979, 28, 2161–2165. [Google Scholar] [CrossRef]
- Brown, G.M.; Verhaegan, H.; Greidanus, T.B.; Brugmans, J. Endocrine effects of domperidone: A peripheral dopamine blocking agent. Clin. Endocrinol. 1981, 15, 275–282. [Google Scholar] [CrossRef]
- Pehek, E.A.; Thompson, J.T.; Hull, E.M. The effects of intrathecal administration of the dopamine agonist apomorphine on penile reflexes and copulation in the male rat. Psychopharmacology 1989, 99, 304–308. [Google Scholar] [CrossRef]
- Stefanick, M.L.; Smith, E.R.; Clark, J.T.; Davidson, J.M. Effects of a potent dopamine receptor agonist, RDS-127, on penile reflexes and seminal emission in intact and spinally transected rats. Pharmacol. Biochem. Behav. 1982, 29, 973–978. [Google Scholar] [CrossRef]
- Melis, M.R.; Mauri, A.; Argiolas, A. Apomorphine- and oxytocin-induced penile erection and yawning in intact and castrated male rats: Effect of sexual steroids. Neuroendocrinology 1994, 59, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Pehek, E.A.; Thompson, J.T.; Hull, E.M. The effects of intracranial administration of the dopamine agonist apomorphine on penile reflexes and seminal emission in the rat. Brain Res. 1989, 500, 325–332. [Google Scholar] [CrossRef]
- Buijs, R.M.; Geffard, M.; Pool, C.W.; Hoorneman, E.M. The dopaminergic innervation of the supraoptic and para-ventricular nucleus. A light and electron microscopical study. Brain Res. 1984, 323, 65–72. [Google Scholar] [CrossRef]
- Lindvall, O.; Björklund, A.; Skagerberg, G. Selective histochemical demonstration of dopamine terminal systems in rat di- and telecephalon: New evidence for dopaminergic innervation of hypothalamic neurosecretory nuclei. Brain Res. 1984, 306, 19–30. [Google Scholar] [CrossRef]
- Argiolas, A.; Gessa, G.L. Central functions of oxytocin. Neurosci. Biobehav. Rev. 1991, 15, 217–231. [Google Scholar] [CrossRef]
- Melis, M.; Argiolas, A.; Gessa, G. Evidence that apomorphine induces penile erection and yawning by releasing oxytocin in the central nervous system. Eur. J. Pharmacol. 1989, 164, 565–570. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.; Mauri, A.; Gessa, G. Paraventricular nucleus lesion prevents yawning and penile erection induced by apomorphine and oxytocin but not by ACTH in rats. Brain Res. 1987, 421, 349–352. [Google Scholar] [CrossRef]
- Hawthorn, J.; Ang, V.T.; Jenkins, J.S. Effects of lesions in the hypothalamic paraventricular, supraoptic and suprachiasmatic nuclei on vasopressin and oxytocin in rat brain and spinal cord. Brain Res. 1985, 346, 51–57. [Google Scholar] [CrossRef]
- Lang, R.E.; Heil, J.; Ganten, D.; Herman, K.; Rascher, W.; Unger, T. Effects of lesions in the paraventricular nucleus of the hypothalamus on vasopressin and oxytocin contents in brainstem and spinal cord of rat. Brain Res. 1983, 260, 326–329. [Google Scholar] [CrossRef]
- McCleskey, E.W.; Fox, A.P.; Feldman, D.H.; Cruz, L.J.; Olivera, B.M.; Tsien, R.W.; Yoshikami, D. ω-Conotoxin: Direct and persistent blockade of specific types of calcium channels in neurons but not muscle. Proc. Natl. Acad. Sci. USA 1987, 84, 4327–4331. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.; Stancampiano, R.; Gessa, G. ω-Conotoxin prevents apomorphine- and oxytocin-induced penile erection and yawning in male rats. Pharmacol. Biochem. Behav. 1990, 37, 253–257. [Google Scholar] [CrossRef]
- Scott, R.H.; Dolphin, A.C. Activation of a G protein promotes agonist responses to calcium channel ligands. Nature 1987, 330, 760–762. [Google Scholar] [CrossRef] [PubMed]
- Stancampiano, R.; Melis, M.R.; Argiolas, A. Apomorphine- and oxytocin-induced penile erection and yawning in male rats: Effect of pertussis toxin. Brain Res. Bull. 1992, 28, 315–318. [Google Scholar] [CrossRef]
- Ignarro, L.J. Biosynthesis and metabolism of endothelium-derived nitric oxide. Annu. Rev. Pharmacol. Toxicol. 1990, 30, 535–560. [Google Scholar] [CrossRef] [PubMed]
- Snyder, S.H. Nitric oxide: First in a new class of neurotransmitters. Science 1992, 257, 494–496. [Google Scholar] [CrossRef]
- Burnett, A.L.; Lowenstein, C.J.; Bredt, D.S.; Chang, T.S.K.; Snyder, S.H. Nitric oxide: A physiologic mediator of penile erection. Science 1992, 257, 401–403. [Google Scholar] [CrossRef] [PubMed]
- Rajfer, J.; Aronson, W.J.; Bush, P.A.; Dorey, F.J.; Ignarro, L.J. Nitric oxide as a mediator of relaxation of the corpus cavernosum in response to nonadrenergic, noncholinergic neurotransmission. N. Engl. J. Med. 1992, 326, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Vincent, S.; Kimura, H. Histochemical mapping of nitric oxide synthase in the rat brain. Neuroscience 1992, 46, 755–784. [Google Scholar] [CrossRef]
- Melis, M.; Argiolas, A.; Stancampiano, R.; Gessa, G. Effect of apomorphine on oxytocin concentrations in different brain areas and plasma of male rats. Eur. J. Pharmacol. 1990, 182, 101–107. [Google Scholar] [CrossRef]
- Melis, M.R.; Stancampiano, R.; Argiolas, A. Hippocampal oxytocin mediates apomorphine-induced penile erection and yawning. Pharmacol. Biochem. Behav. 1992, 42, 61–66. [Google Scholar] [CrossRef]
- Melis, M.R.; Argiolas, A.; Gessa, G.L. Oxytocin-induced penile erection and yawning: Site of action in the brain. Brain Res. 1986, 398, 259–265. [Google Scholar] [CrossRef]
- Melis, M.R.; Stancampiano, R.; Gessa, G.L.; Argiolas, A. Prevention by morphine of apomorphine- and oxytocin-induced penile erection and yawning: Site of action in the brain. Neuropsychopharmacology 1992, 6, 17–21. [Google Scholar] [PubMed]
- Melis, M.R.; Succu, S.; Mascia, M.S.; Cortis, L.; Argiolas, A. Extracellular dopamine increases in the paraventricular nucleus of male rats during sexual activity. Eur. J. Neurosci. 2003, 17, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Pleim, E.T.; Matochik, J.A.; Barfield, R.J.; Auerbach, S.B. Correlation of dopamine release in the nucleus accumbens with masculine sexual behavior in rats. Brain Res. 1990, 524, 160–163. [Google Scholar] [CrossRef]
- Damsma, G.; Pfaus, J.G.; Wenkstern, D.; Phillips, A.G.; Fibiger, H.C. Sexual behavior increases dopamine transmission in the nucleus accumbens and striatum of male rats: Comparison with novelty and locomotion. Behav. Neurosci. 1992, 106, 181–191. [Google Scholar] [CrossRef]
- Mas, M.; Gonzalez-Mora, J.L.; Louilot, A.; Solè, C.; Guadalupe, T. Increased dopamine release in the nucleus accumbens of copulating male rats as evidenced by in vivo voltammetry. Neurosci. Lett. 1990, 110, 303–308. [Google Scholar] [CrossRef]
- Bitran, D.; Thompson, J.T.; Hull, E.M.; Sachs, B.D. Quinelorane (LY 163502), a D2 dopamine receptor agonist, facilitates seminal emission, but inhibits penile erection in the rat. Pharmacol. Biochem. Behav. 1989, 34, 453–458. [Google Scholar] [CrossRef]
- Pehek, E.A.; Thompson, J.T.; Eaton, R.C.; Bassett, T.J.; Hull, E.M. Apomorphine and haloperidol, but not domperidone, affect penile reflexes in rats. Pharmacol. Biochem. Behav. 1988, 31, 201–208. [Google Scholar] [CrossRef]
- Bazzett, T.J.; Eaton, R.C.; Thompson, J.T.; Markowski, V.P.; Lumley, L.A.; Hull, E.M. Dose dependent D2 effects on genital reflexes after MPOA injections of quinelorane and apomorphine. Life Sci. 1991, 48, 2309–2315. [Google Scholar] [CrossRef]
- Warner, R.K.; Thompson, J.T.; Markowski, V.P.; Loucks, J.A.; Bazzett, T.J.; Eaton, R.C.; Hull, E.M. Microinjection of the dopamine antagonist cis-flupenthixol into the MPOA impairs copulation, penile reflexes and sexual motivation in male rats. Brain Res. 1991, 540, 177–182. [Google Scholar] [CrossRef]
- Eaton, R.C.; Markowski, V.P.; Lumley, L.A.; Thompson, J.T.; Moses, J.; Hull, E.A. D2 receptors in the paraventricular nucleus regulate genital responses and copulation in male rats. Pharmacol. Biochem. Behav. 1991, 39, 177–181. [Google Scholar] [CrossRef][Green Version]
- Baskerville, T.A.; Douglas, A.J. Interactions between dopamine and oxytocin in the control of sexual behaviour. Prog. Brain Res. 2008, 170, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Baskerville, T.A.; Allard, J.; Wayman, C.; Douglas, A.J. Dopamine-oxytocin interaction in penile erection. Eur. J. Neurosci. 2009, 30, 2151–2164. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Melis, M.R.; Argiolas, A. Dopamine, erectile function and sexual behavior: Last discoveries and possible advances. In New Developments in Dopamine Research; Hopkins, G., Ed.; Nova Publishers, Inc.: New York, NY, USA, 2016; pp. 45–84. [Google Scholar]
- Bitner, R.S.; Nikkel, A.L.; Otte, S.; Martino, B.; Barlow, E.H.; Bhatia, P.; Stewart, A.O.; Brioni, J.D.; Decker, M.W.; Moreland, R.B. Dopamine D4 receptor signaling in the rat paraventricular hypothalamic nucleus: Evidence of natural coupling involving immediate early gene induction and mitogen activated protein kinase phosphorylation. Neuropharmacology 2006, 50, 521–531. [Google Scholar] [CrossRef]
- Osinski, M.A.; Uchic, M.E.; Seifert, T.; Shaughnessy, T.K.; Miller, L.N.; Natane, M.; Cox, B.F.; Brioni, J.D.; Moreland, R.B. Dopamine D2, but not D4, receptor agonists are emetogenic in ferrets. Pharmacol. Biochem. Behav. 2005, 81, 211–219. [Google Scholar] [CrossRef]
- Brioni, J.D.; Moreland, R.B. Dopamine D4 receptors and the regulation of penile erection. Drug Discov. Today: Ther. Strategies 2006, 3, 599–604. [Google Scholar] [CrossRef]
- Melis, M.R.; Melis, T.; Cocco, C.; Succu, S.; Sanna, F.; Pillolla, G.; Boi, A.; Ferri, G.-L.; Argiolas, A. Oxytocin injected into the ventral tegmental area induces penile erection and increases extracellular dopamine in the nucleus accumbens and paraventricular nucleus of the hypothalamus of male rats. Eur. J. Neurosci. 2007, 26, 1026–1035. [Google Scholar] [CrossRef]
- Succu, S.; Sanna, F.; Cocco, C.; Melis, T.; Boi, A.; Ferri, G.-L.; Argiolas, A.; Melis, M.R. Oxytocin induces penile erection when injected into the ventral tegmental area of male rats: Role of nitric oxide and cyclic GMP. Eur. J. Neurosci. 2008, 28, 813–821. [Google Scholar] [CrossRef]
- Melis, M.R.; Sanna, F.; Succu, S.; Zarone, P.; Boi, A.; Argiolas, A. The role of oxytocin in the anticipatory and consummatory phases of male rat sexual behaviour. In Handbook of Oxytocin Research: Synthesis, Storage and Release, Actions and Drug Forms; Jastrow, H., Feuerbach, D., Eds.; Nova Publishers Inc.: New York, NY, USA, 2009; pp. 109–125. [Google Scholar]
- Melis, M.R.; Succu, S.; Sanna, F.; Boi, A.; Argiolas, A. Oxytocin injected into the ventral subiculum or the posteromedial cortical nucleus of the amygdala induces penile erection and increases extracellular dopamine levels in the nucleus accumbens of male rats. Eur. J. Neurosci. 2009, 30, 1349–1357. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Cocco, C.; Caboni, E.; Sanna, F.; Boi, A.; Ferri, G.L.; Argiolas, A. Oxytocin induces penile erection when injected into the ventral subiculum: Role of nitric oxide and glutamic acid. Neuropharmacology 2010, 58, 1153–1160. [Google Scholar] [CrossRef]
- Succu, S.; Sanna, F.; Argiolas, A.; Melis, M.R. Oxytocin injected into the hippocampal ventral subiculum induces penile erection in male rats by increasing glutamatergic neurotransmission in the ventral tegmental area. Neuropharmacology 2011, 61, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Melis, M.R.; Argiolas, A. Central control of penile erection: A re-visitation of the role of oxytocin and its interaction with dopamine and glutamic acid in male rats. Neurosci. Biobehav. Rev. 2011, 35, 939–955. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Bratzu, J.; Argiolas, A.; Melis, M.R. Oxytocin induces penile erection and yawning when injected into the bed nucleus of the stria terminalis: Involvement of glutamic acid, dopamine, and nitric oxide. Horm. Behav. 2017, 96, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Bratzu, J.; Bharatiya, R.; Manca, E.; Cocco, C.; Argiolas, A.; Melis, M.R.; Sanna, F. Oxytocin induces penile erection and yawning when injected into the bed nucleus of the stria terminalis: A microdialysis and immunohistochemical study. Behav. Brain Res. 2019, 375, 112147. [Google Scholar] [CrossRef] [PubMed]
- Morari, M.; Marti, M.; Sbrenna, S.; Fuxe, K.; Bianchi, C.; Beani, L. Reciprocal dopamine-glutamate modulation of release in the basal ganglia. Neurochem. Int. 1998, 33, 383–397. [Google Scholar] [CrossRef]
- Meloni, E.G.; Gerety, L.P.; Knoll, A.T.; Cohen, B.M.; Carlezon, W.A. Behavioral and anatomical interactions between dopamine and corticotropin-releasing factor in the rat. J. Neurosci. 2006, 26, 3855–3863. [Google Scholar] [CrossRef]
- Beach, F.A. Characteristics of masculine “sex drive”. In Nebraska Symposium on Motivation; Jones, M.R., Ed.; University of Nebraska Press: Lincoln, NE, USA, 1956; Volume 4, pp. 1–31. [Google Scholar]
- Beach, F.A. (Ed.) Hormonal control of sex-related behavior. In Human Sexuality in Four Perspectives; John Hopkins University Press: Baltimore, MD, USA, 1976; pp. 247–267. [Google Scholar]
- Davidson, J.M. Activation of the male rat’s sexual behavior by intracerebral implantation of androgen. Endocrinology 1966, 79, 783–794. [Google Scholar] [CrossRef]
- McClintock, M.K. Group mating in the domestic rat as a context for sexual selection: Consequences for the analysis of sexual behavior and neuroendocrine responses. Adv. Study Behav. 1984, 14, 1–50. [Google Scholar] [CrossRef]
- Mendelson, S.D.; Gorzalka, B.B. An improved chamber for the observation and analysis of the sexual behavior of the female rat. Physiol. Behav. 1987, 39, 67–71. [Google Scholar] [CrossRef]
- Mendelson, S.D.; Pfaus, J. Level searching: A new assay of sexual motivation in the male rat. Physiol. Behav. 1989, 45, 337–341. [Google Scholar] [CrossRef]
- Pfaus, J.; Phillips, A.G. Differential effects of dopamine receptor antagonists on the sexual behavior of male rats. Psychopharmacology 1989, 98, 363–368. [Google Scholar] [CrossRef]
- Sachs, B.D. Conceptual and neural mechanisms of masculine copulatory behavior. In Sex and Behavior: Status and Prospectus; McGill, T.E., Dewsbury, D.A., Sachs, B.D., Eds.; Plenum: New York, NY, USA, 1978; pp. 267–295. [Google Scholar]
- Bitran, D.; Hull, E. Pharmacological analysis of male rat sexual behavior. Neurosci. Biobehav. Rev. 1987, 11, 365–389. [Google Scholar] [CrossRef]
- Sachs, B.D.; Meisel, R.L. The physiology of male sexual behavior. In The Physiology of Reproduction; Knobil, E., Neil, J., Eds.; Raven Press: New York, NY, USA, 1988; Volume 2, pp. 1393–1485. [Google Scholar]
- Hull, E.M.; Meisel, R.L.; Sachs, B.D. Male sexual behavior. In Hormones, Brain and Behavior; Pfaff, D.W., Arnold, A.P., Etgen, A.M., Fahrbach, S.E., Rubin, R.T., Eds.; Academic Press: New York, NY, USA, 2002; pp. 3–137. [Google Scholar]
- Caggiula, A.R.; Shaw, D.H.; Antelman, S.M.; Edwards, D.J. Interactive effects of brain catecholamines and variations in sexual and non-sexual arousal on copulatory behavior of male rats. Brain Res. 1976, 111, 321–336. [Google Scholar] [CrossRef]
- Caggiula, A.R.; Herndon, J.G.; Scanlon, R.; Greenstone, D.; Bradshaw, W.; Sharp, D. Dissociation of active from immobility components of sexual behavior in female rats by central 6-hydroxydopamine: Implications for CA involvement in sexual behavior and sensorimotor responsiveness. Brain Res. 1979, 172, 505–520. [Google Scholar] [CrossRef]
- Burns-Cusato, M.; Scordalakes, E.M.; Rissman, E.F. Of mice and missing data: What we know (and need to learn) about male sexual behavior. Physiol. Behav. 2004, 83, 217–232. [Google Scholar] [CrossRef]
- Okabe, S.; Kitano, K.; Nagasawa, M.; Mogi, K.; Kikusui, T. Testosterone inhibits facilitating effects of parenting experience on parental behavior and the oxytocin neural system in mice. Physiol. Behav. 2013, 118, 159–164. [Google Scholar] [CrossRef]
- Floody, O.R. Processes regulating the initiation and postejaculatory resumption of copulatory behavior in male hamsters. Behav. Process. 2014, 105, 79–84. [Google Scholar] [CrossRef]
- Gray, G.D.; Dewsbury, D.A. A quantitative description of copulatory behavior in prairie voles (Microtus ochrogaster). Brain Behav. Evol. 1973, 8, 437–452. [Google Scholar] [CrossRef]
- Nitsch, F.; Stueckle, S.; Stahl, D.; Zinner, D. Copulation patterns in captive hamadryas baboons: A quantitative analysis. Primates 2011, 52, 373–383. [Google Scholar] [CrossRef]
- Absil, P.; Das, S.; Balthazart, J. Effects of apomorphine on sexual behavior in male quail. Pharmacol. Biochem. Behav. 1994, 47, 77–88. [Google Scholar] [CrossRef]
- Balthazart, J.; Ball, G. Topography in the preoptic region: Differential regulation of appetitive and consummatory male sexual behaviors. Front. Neuroendocr. 2007, 28, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Masters, W.H.; Johnson, V.E. Human Sexual Response; Little Brown & Co. Inc.: Boston, MA, USA, 1966. [Google Scholar]
- Kaplan, H.S. Disorders of Sexual Desire; Brunner/Mazel Inc.: New York, NY, USA, 1979. [Google Scholar]
- Leiblum, S. Classification and diagnosis of female sexual disorders. Int. J. Imp. Res. 1998, 10, S104–S106. [Google Scholar] [CrossRef]
- Lue, T.F.; Tanagho, E.A. Physiology of erection and pharmacological management of impotence. J. Urol. 1987, 137, 829–836. [Google Scholar] [CrossRef]
- Andersson, K.E. Pharmacology of penile erection. Pharmacol. Rev. 2001, 53, 417–500. [Google Scholar]
- Gunnet, J.W.; Lookingland, K.J.; Moore, K.E. Comparison of the effects of castration and steroid replacement on incertohypothalamic dopaminergic neurons in male and female rats. Neuroendocrinology 1986, 44, 269–275. [Google Scholar] [CrossRef]
- Zemlan, F.P.; Ward, I.L.; Crowley, W.R.; Margules, D.L. Activation of lordotic responding in female rats by suppression of serotonergic activity. Science 1973, 179, 1010–1011. [Google Scholar] [CrossRef]
- Agmo, A. Dopamine and sexual behavior in the male rat: A reevaluation. J. Neural Transm. 1989, 77, 21–37. [Google Scholar] [CrossRef]
- Weiner, M. Pharmacology of central nervous system stimulants. In Drug Abuse: Proceedings of the International Conference; Zarafonetis, C.J.D., Ed.; Lea & Febiger: Philadelphia, PA, USA, 1972; pp. 243–251. [Google Scholar]
- Paglietti, E.; Quarantotti, B.P.; Mereu, G.; Gessa, G. Apomorphine and L-DOPA lower ejaculation threshold in the male rat. Physiol. Behav. 1978, 20, 559–562. [Google Scholar] [CrossRef]
- Clark, J.T.; Smith, E.R. Effects of apomorphine on sexual behavior in young and middle-aged rats. Neurobiol. Aging 1987, 8, 153–157. [Google Scholar] [CrossRef]
- Napoli-Farris, L.; Fratta, W.; Gessa, G.L. Stimulation of dopamine autoreceptors elicits “premature ejaculation” in rats. Pharmacol. Biochem. Behav. 1984, 20, 69–72. [Google Scholar] [CrossRef]
- Pfaus, J.G.; Phillips, A.G. Role of dopamine in anticipatory and consummatory aspects of sexual behavior in the male rat. Behav. Neurosci. 1991, 105, 727–743. [Google Scholar] [CrossRef] [PubMed]
- Falaschi, P.; Frajese, G.; Rocco, A.; Pompei, P.; Gessa, G.L. Role of brain dopamine in the regulation of male sexual behavior. In The Menopause: Clinical, Endocrinological and Pathophysiological Aspects; Fioretti, P., Ed.; Academic Press: New York, NY, USA, 1982; pp. 439–446. [Google Scholar]
- Montejo, A.L.; de Alarcón, R.; Prieto, N.; Acosta, J.M.; Buch, B.; Montejo, L. Management strategies for antipsychotic-related sexual dysfunction: A clinical approach. J. Clin. Med. 2021, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Ahlenius, S.; Engel, J.; Larsson, K.; Svensson, L.; Ahlenius, D.D.S. Effects of pergolide and bromocriptine on male rat sexual behavior. J. Neural Transm. 1982, 54, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Ahlenius, S.; Larsson, K.; Svensson, L. Stimulating effects of lisuride on masculine sexual behavior of rats. Eur. J. Pharmacol. 1980, 64, 47–51. [Google Scholar] [CrossRef]
- Clark, J.T.; Smith, E.R.; Stefanick, M.L.; Arnerić, S.P.; Long, J.P.; Davidson, J.M. Effects of a novel dopamine-receptor agonist RDS-127 (2-N,N-di-n-propylamino-4,7-dimethoxyindane), on hormone levels and sexual behavior in the male rat. Physiol. Behav. 1982, 29, 1–6. [Google Scholar] [CrossRef]
- Clark, J.T.; Stefanick, M.L.; Smith, E.R.; Davidson, J.M. Further studies on alterations in male rat copulatory behavior induced by the dopamine-receptor agonist RDS-127. Pharmacol. Biochem. Behav. 1983, 19, 781–786. [Google Scholar] [CrossRef]
- Dallo, J.; Lekka, N.; Knoll, J. The ejaculatory behavior of sexually sluggish male rats treated with (-) deprenyl, apomorphine, bromocriptine and amphetamine. Pol. J. Pharmacol. Pharm. 1986, 38, 251–255. [Google Scholar]
- Ahlenius, S.; Larsson, K. Lisuride, LY-141865, and 8-OH-DPAT facilitate male rat sexual behavior via a non-dopaminergic mechanism. Psychopharmacology 1984, 83, 330–334. [Google Scholar] [CrossRef]
- Clark, J.T.; Peroutka, S.J.; Ciaranello, R.D.; Smith, E.R.; Davidson, J.M. Central effects of RDS-127: Sexual behavior after intracerebroventricular administration and in vitro receptor binding studies. Behav. Brain Res. 1985, 18, 251–260. [Google Scholar] [CrossRef]
- Da Prada, M.; Bonetti, E.; Keller, H. Induction of mounting behaviour in female and male rats by lisuride. Neurosci. Lett. 1977, 6, 349–353. [Google Scholar] [CrossRef]
- Fernández-Guasti, A.; Escalante, A.; Agamo, A. Inhibitory action of various 5-HT1B receptor agonists on rat masculine sexual behavior. Pharmacol. Biochem. Behav. 1989, 34, 811–816. [Google Scholar] [CrossRef]
- Svensson, L.; Hansen, S. Spinal monoaminergic modulation of masculine copulatory behavior in the rat. Brain Res. 1984, 302, 315–321. [Google Scholar] [CrossRef]
- Stoof, J.C.; Kebabian, J.W. Opposing roles for D-1 and D-2 dopamine receptors in efflux of cyclic AMP from rat neostriatum. Nature 1981, 294, 366–368. [Google Scholar] [CrossRef] [PubMed]
- Stoof, J.; Kebabian, J. Two dopamine receptors: Biochemistry, physiology and pharmacology. Life Sci. 1984, 35, 2281–2296. [Google Scholar] [CrossRef]
- Titus, R.; Kornfeld, E.; Jones, N.; Clemens, J.; Smalstig, E.; Fuller, R.; Hynes, M.; Mason, N.; Wong, D.; Foreman, M. Resolution and absolute configuration of an ergoline-related dopamine agonist, trans-4,4a,5,6,7,8,8a,9-octahydro-5-propyl-1H(or 2H)-pyrazolo[3,4-g]quinoline. J. Med. Chem. 1983, 26, 1112–1116. [Google Scholar] [CrossRef]
- Foreman, M.M.; Hall, J.L. Effects of D2-dopaminergic receptor stimulation on male rat sexual behavior. J. Neural Transm. 1987, 68, 153–170. [Google Scholar] [CrossRef]
- Hyttel, J. The first selective dopamine D1 antagonist. Eur. J. Pharmacol. 1983, 91, 153–154. [Google Scholar] [CrossRef]
- Paglietti, E.; Pellegrini-Quarantotti, B.; Fratta, W.; Gessa, G.L. Failure of (-) sulpiride to inhibit sexual behavior in rats. In Sulpiride and Other Benzamides; Spano, P.F., Ed.; Italian Brain Research Foundation Press: Milan, Italy, 1979; pp. 157–162. [Google Scholar]
- Falaschi, P.; Rocco, A.; De Giorgio, G.; Frajese, G.; Fratta, W.; Gessa, G.L. Brain dopamine and premature ejaculation: Results of treatment with dopamine antagonists. In Apomorphine and Other Dopaminomimetics; Gessa, G.L., Corsini, G.U., Eds.; Basic Pharmacology; Raven Press: New York, USA, 1981; Volume 1, pp. 117–121. [Google Scholar]
- Giuliani, D.; Ferrari, F. Differential behavioral response to dopamine D-sub-2 agonists by sexually naive, sexually active, and sexually inactive male rats. Behav. Neurosci. 1996, 110, 802–808. [Google Scholar] [CrossRef]
- Newman-Tancredi, A.; Heusler, P.; Martel, J.-C.; Ormière, A.-M.; Leduc, N.; Cussac, D. Agonist and antagonist properties of antipsychotics at human dopamine D4.4 receptors: G-protein activation and K+ channel modulation in transfected cells. Int. J. Neuropsychopharmacol. 2008, 11, 293–307. [Google Scholar] [CrossRef]
- Driscoll, P.; Battig, K. Behavioral, emotional and neurochemical profiles of rats selected for extreme differences in active, two-way avoidance performance. In Genetics of the Brain; Lieblich, I., Ed.; Elsevier: Amsterdam, The Netherlands, 1982; pp. 95–123. [Google Scholar]
- Steimer, T.; Driscoll, P. Divergent stress responses and coping styles in psychogenetically selected Roman high-(RHA) and low-(RLA) avoidance rats: Behavioural, neuroendocrine and developmental aspects. Stress 2003, 6, 87–100. [Google Scholar] [CrossRef]
- Steimer, T.; Driscoll, P. Inter-individual vs line/strain differences in psychogenetically selected Roman High-(RHA) and Low-(RLA) Avoidance rats: Neuroendocrine and behavioural aspects. Neurosci. Biobehav. Rev. 2005, 29, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Stead, J.D.H.; Clinton, S.; Neal, C.; Schneider, J.; Jama, A.; Miller, S.; Vazquez, D.M.; Watson, S.J.; Akil, H. Selective breeding for divergence in novelty-seeking traits: Heritability and enrichment in spontaneous anxiety-related behaviors. Behav. Genet. 2006, 36, 697–712. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, B.; Urbá-Holmgren, R.; Trucios, N.; Zermeño, M.; Eguíbar, J. Association of spontaneous and dopaminergic-induced yawning and penile erections in the rat. Pharmacol. Biochem. Behav. 1985, 22, 31–35. [Google Scholar] [CrossRef]
- Urbá-Holmgren, R.; Trucios, N.; Holmgren, B.; Eguibar, J.; Gavito, A.; Cruz, G.; Santos, A. Genotypic dependency of spontaneous yawning frequency in the rat. Behav. Brain Res. 1990, 40, 29–35. [Google Scholar] [CrossRef]
- Sanna, F.; Argiolas, A.; Melis, M.R. Oxytocin-induced yawning: Sites of action in the brain and interaction with mesolimbic/mesocortical and incertohypothalamic dopaminergic neurons in male rats. Horm. Behav. 2012, 62, 505–514. [Google Scholar] [CrossRef]
- Walusinski, O. Historical perspectives. Front. Neurol. Neurosci. 2010, 28, 1–21. [Google Scholar] [CrossRef]
- Walusinski, O. Popular knowledge and beliefs. Front. Neurol. Neurosci. 2010, 28, 22–25. [Google Scholar] [CrossRef]
- Walusinski, O. Fetal yawning. Front. Neurol. Neurosci. 2010, 28, 32–41. [Google Scholar] [CrossRef]
- Corda, M.G.; Lecca, D.; Piras, G.; Di Chiara, G.; Giorgi, O. Biochemical parameters of dopaminergic and GABAergic neurotransmission in the CNS of Roman High-Avoidance and Roman Low-Avoidance rats. Behav. Genet. 1997, 27, 527–536. [Google Scholar] [CrossRef]
- Escorihuela, R.M.; Tobeña, A.; Driscoll, P.; Fernández-Teruel, A. Effects of training, early handling, and perinatal flumazenil on shuttle box acquisition in Roman low-avoidance rats: Toward overcoming a genetic deficit. Neurosci. Biobehav. Rev. 1995, 19, 353–367. [Google Scholar] [CrossRef]
- Gentsch, C.; Lichtsteiner, M.; Driscoll, P.; Feer, H. Differential hormonal and physiological responses to stress in Roman high- and low-avoidance rats. Physiol. Behav. 1982, 28, 259–263. [Google Scholar] [CrossRef]
- Walker, C.-D.; Rivest, R.W.; Meaney, M.J.; Aubert, M.L. Differential activation of the pituitary-adrenocortical axis after stress in the rat: Use of two genetically selected lines (Roman low- and high-avoidance rats) as a model. J. Endocrinol. 1989, 123, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, P.; Escorihuela, R.M.; Fernandez-Teruel, A.; Giorgi, O.; Schwegler, H.; Steimer, T.; Wiersma, A.; Corda, M.G.; Flint, J.; Koolhaas, J.M.; et al. Genetic selection and differential stress responses: The Roman lines/strains of rats. Ann. N. Y. Acad. Sci. 1998, 851, 501–510. [Google Scholar] [CrossRef]
- Escorihuela, R.M.; Fernández-Teruel, A.; Gil, L.; Aguilar, R.; Tobeña, A.; Driscoll, P. Inbred Roman high- and low-avoidance rats: Differences in anxiety, novelty-seeking, and shuttlebox behaviors. Physiol. Behav. 1999, 67, 19–26. [Google Scholar] [CrossRef]
- Fernandez-Teruel, A.; Escorihuela, R.M.; Castellano, B.; González, B.; Tobena, A. Neonatal handling and environmental enrichment effects on emotionality, novelty/reward seeking, and age-related cognitive and hippocampal impairments: Focus on the Roman rat lines. Behav. Genet. 1997, 27, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Siegel, J. Augmenting and reducing of visual evoked potentials in high- and low-sensation seeking humans, cats, and rats. Behav. Genet. 1997, 27, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Zeier, H.; Baettig, K.; Driscoll, P. Acquisition of DRL-20 behavior in male and female, Roman high- and low-avoidance rats. Physiol. Behav. 1978, 20, 791–793. [Google Scholar] [CrossRef]
- Giorgi, O.; Corda, M.G.; Carboni, G.; Frau, V.; Valentini, V.; Di Chiara, G. Effects of cocaine and morphine in rats from two psychogenetically selected lines: A behavioral and brain dialysis study. Behav. Genet. 1997, 27, 537–546. [Google Scholar] [CrossRef]
- Corda, M.G.; Piras, G.; Piludu, M.A.; Giorgi, O. Differential effects of voluntary ethanol consumption on dopamine output in the nucleus accumbens shell of Roman high- and low-avoidance rats: A behavioral and brain microdialysis study. World J. Neurosci. 2014, 4, 279–292. [Google Scholar] [CrossRef][Green Version]
- Giorgi, O.; Piras, G.; Corda, M.G. The psychogenetically selected Roman high- and low-avoidance rat lines: A model to study the individual vulnerability to drug addiction. Neurosci. Biobehav. Rev. 2007, 31, 148–163. [Google Scholar] [CrossRef]
- Fattore, L.; Piras, G.; Corda, M.G.; Giorgi, O. The Roman high- and low-avoidance rat lines differ in the acquisition, maintenance, extinction, and reinstatement of intravenous cocaine self-administration. Neuropsychopharmacology 2009, 34, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- López-Aumatell, R.; Vicens-Costa, E.; Masip, M.G.; Martínez-Membrives, E.; Valdar, W.; Johannesson, M.; Cañete, T.; Blázquez, G.; Driscoll, P.; Flint, J.; et al. Unlearned anxiety predicts learned fear: A comparison among heterogeneous rats and the Roman rat strains. Behav. Brain Res. 2009, 202, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Cardona, D.; Gómez, M.J.; Sánchez-Santed, F.; Tobena, A.; Fernandez-Teruel, A.; Campa, L.; Suñol, C.; Escarabajal, M.D.; Torres, C.; et al. Impulsivity characterization in the Roman high- and low-avoidance rat strains: Behavioral and neurochemical differences. Neuropsychopharmacology 2010, 35, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Llort, L.; Cañete, T.; Guitart-Masip, M.; Fernández-Teruel, A.; Tobeña, A. Differential apomorphine-induced locomotion, stereotypy and yawning patterns in Roman high- and low avoidance rats: Revealing two singular dopaminergic phenotypes. Physiol. Behav. 2005, 86, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Guitart-Masip, M.; Johansson, B.; Fernández-Teruel, A.; Cañete, T.; Tobena, A.; Terenius, L.; Giménez-Llort, L. Divergent anatomical pattern of D1 and D3 binding and DARPP-32 mRNA expression in the Roman rat strains. Implications for drug addiction. Neuroscience 2006, 142, 1231–1243. [Google Scholar] [CrossRef]
- Sanna, F.; Corda, M.G.; Melis, M.R.; Piludu, M.A.; Giorgi, O.; Argiolas, A. Male Roman high and low avoidance rats show different patterns of copulatory behaviour: Comparison with Sprague Dawley rats. Physiol. Behav. 2014, 127, 27–36. [Google Scholar] [CrossRef]
- Cummings, J.A.; Clinton, S.M.; Perry, A.N.; Akil, H.; Becker, J.B. Male rats that differ in novelty exploration demonstrate distinct patterns of sexual behavior. Behav. Neurosci. 2013, 127, 47–58. [Google Scholar] [CrossRef]
- Sachs, B.D.; Barfield, R.J. Functional analysis of masculine copulatory behavior in the rat. Adv. Study Behav. 1976, 7, 91–154. [Google Scholar] [CrossRef]
- Pfaus, J.G.; Everitt, B.J. The psychopharmacology of sexual behavior. In Psychopharmacology: The Fourth Generation of Progress; Knobil, F.E., Kupfer, D.J., Eds.; Raven Press: New York, NY, USA, 1995; pp. 742–758. [Google Scholar]
- Giorgi, O.; Orlandi, M.; Escorihuela, R.M.; Driscoll, P.; Lecca, D.; Corda, M. GABAergic and dopaminergic transmission in the brain of Roman high-avoidance and Roman low-avoidance rats. Brain Res. 1994, 638, 133–138. [Google Scholar] [CrossRef]
- Tournier, B.B.; Steimer, T.; Millet, P.; Moulin-Sallanon, M.; Vallet, P.; Ibañez, V.; Ginovart, N. Innately low D2 receptor availability is associated with high novelty-seeking and enhanced behavioural sensitization to amphetamine. Int. J. Neuropsychopharmacol. 2013, 16, 1819–1834. [Google Scholar] [CrossRef]
- Sanna, F.; Piludu, M.A.; Corda, M.G.; Argiolas, A.; Giorgi, O.; Melis, M.R. Dopamine is involved in the different patterns of copulatory behaviour of Roman high and low avoidance rats: Studies with apomorphine and haloperidol. Pharmacol. Biochem. Behav. 2014, 124, 211–219. [Google Scholar] [CrossRef]
- Sanna, F.; Piludu, M.A.; Corda, M.G.; Melis, M.R.; Giorgi, O.; Argiolas, A. Involvement of dopamine in the differences in sexual behaviour between Roman high and low avoidance rats: An intracerebral microdialysis study. Behav. Brain Res. 2015, 281, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Bratzu, J.; Piludu, M.A.; Corda, M.G.; Melis, M.R.; Giorgi, O.; Argiolas, A. Dopamine, Noradrenaline and Differences in Sexual Behavior between Roman High and Low Avoidance Male Rats: A Microdialysis Study in the Medial Prefrontal Cortex. Front. Behav. Neurosci. 2017, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Hull, E.M.; Muschamp, J.W.; Sato, S. Dopamine and serotonin: Influences on male sexual behavior. Physiol. Behav. 2004, 83, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Pfaus, J.G.; Damsma, G.; Nomikos, G.G.; Wenkstern, D.G.; Blaha, C.D.; Phillips, A.G.; Fibiger, H.C. Sexual behavior enhances central dopamine transmission in the male rat. Brain Res. 1990, 530, 345–348. [Google Scholar] [CrossRef]
- Pfaus, J.G. Dopamine: Helping males copulate for at least 200 million years: Theoretical comment on Kleitz-Nelson et al. Behav. Neurosci. 2010, 124, 877–883. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.R. Central control of penile erection: Role of the paraventricular nucleus of the hypothalamus. Prog. Neurobiol. 2005, 76, 1–21. [Google Scholar] [CrossRef]
- Hull, E.M.; Du, J.; Lorrain, D.S.; Matuszewich, L. Extracellular dopamine in the medial preoptic area: Implications for sexual motivation and hormonal control of copulation. J. Neurosci. 1995, 15, 7465–7471. [Google Scholar] [CrossRef][Green Version]
- Hull, E.M.; Lorrain, D.S.; Du, J.; Matuszewich, L.; A Lumley, L.; Putnam, S.K.; Moses, J. Hormone-neurotransmitter interactions in the control of sexual behavior. Behav. Brain Res. 1999, 105, 105–116. [Google Scholar] [CrossRef]
- Dominguez, J.M.; Muschamp, J.W.; Schmich, J.M.; Hull, E.M. Nitric oxide mediates glutamate-evoked dopamine release in the medial preoptic area. Neuroscience 2004, 125, 203–210. [Google Scholar] [CrossRef]
- Dominguez, J.M.; Gil, M.; Hull, E.M. Preoptic glutamate facilitates male sexual behavior. J. Neurosci. 2006, 26, 1699–1703. [Google Scholar] [CrossRef] [PubMed]
- Kerman, I.A.; Clinton, S.M.; Bedrosian, T.A.; Abraham, A.D.; Rosenthal, D.T.; Akil, H.; Watson, S.J. High novelty-seeking predicts aggression and gene expression differences within defined serotonergic cell groups. Brain Res. 2011, 1419, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Flagel, S.B.; Clark, J.J.; Robinson, T.E.; Mayo, L.; Czuj, A.; Willuhn, I.; Akil, H. A selective role for dopamine in stimulus–reward learning. Nature 2011, 469, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.A.; Gowl, B.A.; Westenbroek, C.; Clinton, S.M.; Akil, H.; Becker, J.B. Effects of a selectively bred novelty-seeking phenotype on the motivation to take cocaine in male and female rats. Biol. Sex Differ. 2011, 2, 3. [Google Scholar] [CrossRef]
- Garcãa-Fuster, M.J.; Perez, J.A.; Clinton, S.M.; Watson, S.J.; Akil, H. Impact of cocaine on adult hippocampal neurogenesis in an animal model of differential propensity to drug abuse. Eur. J. Neurosci. 2010, 31, 79–89. [Google Scholar] [CrossRef]
- Clinton, S.M.; Stead, J.D.; Miller, S.; Watson, S.J.; Akil, H. Developmental underpinnings of differences in rodent novelty-seeking and emotional reactivity. Eur. J. Neurosci. 2011, 34, 994–1005. [Google Scholar] [CrossRef]
- Flagel, S.B.; Robinson, T.E.; Clark, J.J.; Clinton, S.M.; Watson, S.J.; Seeman, P.; Phillips, P.; Akil, H. An animal model of genetic vulnerability to behavioral disinhibition and responsiveness to reward-related cues: Implications for addiction. Neuropsychopharmacology 2010, 35, 388–400. [Google Scholar] [CrossRef]
- García-Fuster, M.; Parks, G.; Clinton, S.; Watson, S.; Akil, H.; Civelli, O. The melanin-concentrating hormone (MCH) system in an animal model of depression-like behavior. Eur. Neuropsychopharmacol. 2012, 22, 607–613. [Google Scholar] [CrossRef]
- Stedenfeld, K.A.; Clinton, S.M.; Kerman, I.A.; Akil, H.; Watson, S.J.; Sved, A.F. Novelty-seeking behavior predicts vulnerability in a rodent model of depression. Physiol. Behav. 2011, 103, 210–216. [Google Scholar] [CrossRef]
- Hooks, M.S.; Jones, G.H.; Smith, A.D.; Neill, D.B.; Justice, J.B., Jr. Response to novelty predicts the locomotor and nucleus accumbens dopamine response to cocaine. Synapse 1991, 9, 121–128. [Google Scholar] [CrossRef]
- Hooks, M.S.; Juncos, J.L.; Justice, J.B., Jr.; Meiergerd, S.M.; Povlock, S.L.; Schenk, J.O.; Kalivas, P.W. Individual locomotor response to novelty predicts selective alterations in D1 and D2 receptors and mRNAs. J. Neurosci. 1994, 14, 6144–6152. [Google Scholar] [CrossRef] [PubMed]
- Hooks, M.S.; Kalivas, P.W. Involvement of dopamine and excitatory amino acid transmission in noveltyinduced motor activity. J. Pharmacol. Exp. Ther. 1994, 269, 976–988. [Google Scholar]
- Piazza, P.V.; Rouge-Pont, F.; Deminiere, J.M.; Kharoubi, M.; Le Moal, M.; Simon, H. Dopaminergic activity is reduced in the prefrontal cortex and increased in the nucleus accumbens of rats predisposed to develop amphetamine self-administration. Brain Res. 1991, 567, 169–174. [Google Scholar] [CrossRef]
- Rouge-Pont, F.; Deroche, V.; Le Moal, M.; Piazza, P.V. Individual differences in stress-induced dopamine release in the nucleus accumbens are influenced by corticosterone. Eur. J. Neurosci. 1998, 10, 3903–3907. [Google Scholar] [CrossRef] [PubMed]
- Mabrouk, O.S.; Han, J.L.; Wong, J.T.; Akil, H.; Kennedy, R.T.; Flagel, S.B. The in vivo neurochemical profile of selectively bred high-responder and low-responder rats reveals baseline, cocaine-evoked, and novelty-evoked differences in monoaminergic systems. ACS Chem. Neurosci. 2018, 9, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Eguibar, J.R.; Romero-Carbente, J.C.; Moyaho, A. Behavioral differences between selectively bred rats: D1 versus D2 receptors in yawning and grooming. Pharmacol. Biochem. Behav. 2003, 74, 827–832. [Google Scholar] [CrossRef]
- Eguibar, J.R.; Cortes, C.; Isidro, O.; Ugarte, A. Central administration of oxytocin differentially increases yawning, penile erections and scratching in high- (HY) and low-yawning (LY) sublines of Sprague–Dawley rats. Pharmacol. Biochem. Behav. 2015, 134, 6–11. [Google Scholar] [CrossRef]
- Eguibar, J.R.; Cortes, C.; Toriz, C.G.; Romero-Carbente, J.C.; Gonzales-Flores, O.; Fernández-Guasti, A. Differential organization of male copulatory patterns in high- and low-yawning-frequency sublines versus outbred Sprague–Dawley rats. Physiol. Behav. 2016, 153, 84–90. [Google Scholar] [CrossRef]
- Portillo, W.; Camacho, F.; Eguibar, J.R.; Paredes, R.G. Behavioral characterization of non-copulating male rats with high spontaneous yawning frequency rate. Behav. Brain Res. 2010, 214, 225–230. [Google Scholar] [CrossRef]
- Portillo, W.; Paredes, R.G. Sexual and olfactory preference in noncopulating male rats. Physiol. Behav. 2003, 80, 155–162. [Google Scholar] [CrossRef]
- Portillo, W.; Paredes, R.G. Sexual incentive motivation, olfactory preference, and activation of the vomeronasal projection pathway by sexually relevant cues in non-copulating and naive male rats. Horm. Behav. 2004, 46, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Portillo, W.; Antonio-Cabrera, E.; Camacho, F.J.; Diaz, N.F.; Paredes, R.G. Behavioral characterization of non-copulating male mice. Horm. Behav. 2013, 64, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Giros, B.; Jaber, M.; Jones, S.R.; Wightman, R.M.; Caron, M.G. Hyperlocomotion and indifference to cocaine and amphetamine in mice lacking the dopamine transporter. Nature 1996, 379, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, F.; Racagni, G.; Colombo, E.; Riva, M.A. BDNF gene expression is reduced in the frontal cortex of dopamine transporter knockout mice. Mol. Psychiatry 2003, 8, 898–899. [Google Scholar] [CrossRef]
- Cyr, M.; Beaulieu, J.-M.; Laakso, A.; Sotnikova, T.D.; Yao, W.-D.; Bohn, L.M.; Gainetdinov, R.R.; Caron, M.G. Sustained elevation of extracellular dopamine causes motor dysfunction and selective degeneration of striatal GABAergic neurons. Proc. Natl. Acad. Sci. USA 2003, 100, 11035–11040. [Google Scholar] [CrossRef]
- Yao, W.D.; Gainetdinov, R.R.; Arbuckle, M.I.; Sotnikova, T.D.; Cyr, M.; Beaulieu, J.M.; Torres, G.E.; Grant, S.G.N.; Grant, M.G. Identification of PSD-95 as a regulator of dopamine-mediated synaptic and behavioral plasticity. Neuron 2004, 41, 625–638. [Google Scholar] [CrossRef]
- Efimova, E.V.; Gainetdinov, R.R.; Budygin, E.A.; Sotnikova, T.D. Dopamine transporter mutant animals: A translational perspective. J. Neurogenet. 2016, 30, 5–15. [Google Scholar] [CrossRef]
- Leo, D.; Sukhanov, I.; Zoratto, F.; Illiano, P.; Caffino, L.; Sanna, F.; Messa, G.; Emanuele, M.; Esposito, A.; Dorofeikova, M.; et al. Pronounced hyperactivity, cognitive dysfunctions, and BDNF dysregulation in dopamine transporter knock-out rats. J. Neurosci. 2018, 38, 1959–1972. [Google Scholar] [CrossRef]
- Drago, J.; Gerfen, C.R.; Lachowicz, J.E.; Steiner, H.; Hollon, T.R.; Love, P.E.; Ooi, G.T.; Grinberg, A.; Lee, E.J.; Huang, S.P.; et al. Altered striatal function in a mutant mouse lacking D1A dopamine receptors. Proc. Natl. Acad. Sci. USA 1994, 91, 12564–12568. [Google Scholar] [CrossRef]
- Baik, J.-H.; Picetti, R.; Saiardi, A.; Thiriet, G.; Dierich, A.; Depaulis, A.; Le Meur, M.; Borrelli, E. Parkinsonian-like locomotor impairment in mice lacking dopamine D2 receptors. Nature 1995, 377, 424–428. [Google Scholar] [CrossRef]
- Accili, D.; Fishburn, C.S.; Drago, J.; Steiner, H.; Lachowicz, J.E.; Park, B.H.; Gauda, E.B.; Lee, E.J.; Cool, M.H.; Sibley, D.R.; et al. A targeted mutation of the D3 dopamine receptor gene is associated with hyperactivity in mice. Proc. Natl. Acad. Sci. USA 1996, 93, 1945–1949. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.; Phillips, T.J.; Bunzow, J.R.; Falzone, T.; Dziewczapolski, G.; Zhang, G.; Fang, Y.; Larson, J.L.; McDougall, J.A.; Chester, J.A.; et al. Mice lacking dopamine D4 receptors are supersensitive to ethanol, cocaine, and methamphetamine. Cell 1997, 90, 991–1001. [Google Scholar] [CrossRef][Green Version]
- Bortolato, M.; Floris, G.; Shih, J.C. From aggression to autism: New perspectives on the behavioral sequelae of monoamine oxidase deficiency. J. Neural Transm. 2018, 125, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Bratzu, J.; Serra, M.P.; Leo, D.; Quartu, M.; Boi, M.; Espinoza, S.; Gainetdinov, R.R.; Melis, M.R.; Argiolas, A. Altered sexual behavior in Dopamine Transporter (DAT) Knockout male rats: A Behavioral, Neurochemical and Intracerebral Microdialysis Study. Front. Behav. Neurosci. 2020, 14, 58. [Google Scholar] [CrossRef] [PubMed]
- Adinolfi, A.; Carbone, C.; Leo, D.; Gainetdinov, R.R.; Laviola, G.; Adriani, W. Novelty-related behavior of young and adult dopamine transporter knockout rats: Implication for cognitive and emotional phenotypic patterns. Genes Brain Behav. 2018, 17, e12463. [Google Scholar] [CrossRef]
- Adinolfi, A.; Zelli, S.; Leo, D.; Carbone, C.; Mus, L.; Illiano, P.; Alleva, E.; Gainetdinov, R.; Adriani, W. Behavioral characterization of DAT-KO rats and evidence of asocial-like phenotypes in DAT-HET rats: The potential involvement of norepinephrine system. Behav. Brain Res. 2019, 359, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Cinque, S.; Zoratto, F.; Poleggi, A.; Leo, D.; Cerniglia, L.; Cimino, S.; Tambelli, R.; Alleva, E.; Gainetdinov, R.R.; Laviola, G.; et al. Behavioral phenotyping of dopamine transporter knockout rats: Compulsive traits, motor stereotypies, and anhedonia. Front. Psychiatry 2018, 9, 43. [Google Scholar] [CrossRef]
- Apryatin, S.A.; Shipelin, V.A.; Trusov, N.V.; Mzhelskaya, K.V.; Evstratova, V.S.; Kirbaeva, N.V.; Soto, J.S.; Fesenko, Z.F.; Gainetdinov, R.R.; Gmoshinski, I.V. Comparative analysis of the influence of a high-fat/high-carbohydrate diet on the level of anxiety and neuromotor and cognitive functions in Wistar and DAT-KO rats. Physiol. Rep. 2019, 7, e13987. [Google Scholar] [CrossRef]
- Mariano, S.; Pardo, M.; Buccheri, C.; Illiano, P.; Adinolfi, A.; Russo, S.L.M.L.; Alleva, E.; Carbone, C.; Adriani, W. Own or dam’s genotype? Classical colony breeding may bias spontaneous and stress-challenged activity in DAT-mutant rats. Dev. Psychobiol. 2020, 62, 505–518. [Google Scholar] [CrossRef]
- Fielding, S.; Lal, H. Behavioral actions of neuroleptics. In Handbook of Psychopharmacology; Iversen, L.L., Iversen, S.D., Snyder, S.H., Eds.; Plenum Press: New York, NY, USA, 1978; Volume 10, pp. 91–128. [Google Scholar]
- Everitt, B. Sexual motivation: A neural and behavioural analysis of the mechanisms underlying appetitive and copulatory responses of male rats. Neurosci. Biobehav. Rev. 1990, 14, 217–232. [Google Scholar] [CrossRef]
- Everitt, B.J.; Cador, M.; Robbins, T.W. Interactions between the amygdala and ventral striatum in stimulus-reward associations: Studies using a second-order schedule of sexual reinforcement. Neuroscience 1989, 30, 63–75. [Google Scholar] [CrossRef]
- Ahlenius, S.; Carlsson, A.; Hillegaart, V.; Hjorth, S.; Larsson, K. Region-selective activation of brain monoamine synthesis by sexual activity in the male rat. Eur. J. Pharmacol. 1987, 144, 77–82. [Google Scholar] [CrossRef]
- Ahlenius, S.; Hillegaart, V.; Hjorth, S.; Larsson, K. Effects of sexual interactions on the in vivo rate of monoamine synthesis in forebrain regions of the male rat. Behav. Brain Res. 1991, 46, 117–122. [Google Scholar] [CrossRef]
- Ahlenius, S.; Hillegaart, V.; Larsson, K. Motivation and performance: Region-selective changes in forebrain monoamine synthesis due to sexual activity in rats. In Behavioral Biology: Neuroendocrine Axes; Archer, T., Hansen, S., Eds.; Lawrance Erlbaum Associates: Hillsdale, MI, USA, 1991; pp. 93–103. [Google Scholar]
- Mas, M.; Rodriguez del Castillo, A.; Guerra, M.; Davidson, J.M.; Battaner, E. Neurochemical correlates of male sexual behavior. Physiol. Behav. 1987, 41, 341–345. [Google Scholar] [CrossRef]
- Blackburn, J.R.; Pfaus, J.G.; Phillips, A.G. Dopamine functions in appetitive and defensive behaviours. Prog. Neurobiol. 1992, 39, 247–279. [Google Scholar] [CrossRef]
- Everitt, B.J.; Robbins, T. Neural systems of reinforcement for drug addiction: From actions to habits to compulsion. Nat. Neurosci. 2005, 8, 1481–1489. [Google Scholar] [CrossRef]
- Sachs, B.D. Erection evoked in male rats by airborne scent from estrous females. Physiol. Behav. 1997, 62, 921–924. [Google Scholar] [CrossRef]
- Heimer, L.; Larsson, K. Impairment of mating behavior in male rats following lesions in the preoptic-anterior hypothalamic continuum. Brain Res. 1967, 3, 248–263. [Google Scholar] [CrossRef]
- Fibiger, H.C.; Phillips, A.G. Mesocorticolimbic dopamine systems and reward. Ann. N. Y. Acad. Sci. 1988, 537, 206–215. [Google Scholar] [CrossRef]
- Wise, R.A.; Rompre, P.-P. Brain dopamine and reward. Annu. Rev. Psychol. 1989, 40, 191–225. [Google Scholar] [CrossRef]
- Goto, Y.; Grace, A.A. Dopaminergic modulation of limbic and cortical drive of nucleus accumbens in goal-directed behavior. Nat. Neurosci. 2005, 8, 805–812. [Google Scholar] [CrossRef]
- Melis, M.R.; Sanna, F.; Argiolas, A. Rats selectively bred for showing divergent behavioral traits in response to stress or novelty or spontaneous yawning with a divergent frequency show similar changes in sexual behavior: The role of dopamine. Rev. Neurosci. 2019, 30, 427–454. [Google Scholar] [CrossRef] [PubMed]
- Viggiano, D.; Ruocco, L.A.; Sadile, A.G. Dopamine phenotype and behaviour in animal models: In relation to attention deficit hyperactivity disorder. Neurosci. Biobehav. Rev. 2003, 27, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Ågmo, A.; Laan, E. Sexual incentive motivation, sexual behavior, and general arousal: Do rats and humans tell the same story? Neurosci. Biobehav. Rev. 2022, 135, 104595. [Google Scholar] [CrossRef] [PubMed]
- Rettemberger, M.; Kelin, V.; Briken, P. The relationship between hypersexual behavior, sexual excitation, sexual inhibition, and personality traits. Arch. Sex. Behav. 2016, 45, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Derefinko, K.J.; Peters, J.R.; Eisenlohr-Moul, T.A.; Walsh, E.C.; Adams, Z.W.; Lunam, D.R. Relations between trait impulsivity, behavioral impulsivity, physiological arousal, and risky sexual behavior among young men. Arch. Sex. Behav. 2014, 43, 1149–1158. [Google Scholar] [CrossRef]
- Skakoon-Sparling, S.; Cramer, K.M.; Shuper, P.A. The impact of sexual arousal on sexual risk-taking and decision-making in men and women. Arch. Sex. Behav. 2016, 45, 33–42. [Google Scholar] [CrossRef]
- Solla, P.; Bortolato, M.; Cannas, A.; Mulas, C.S.; Marrosu, F. Paraphilias and paraphilic disorders in Parkinson’s disease: A systematic review of the literature. Mov. Disord. 2015, 30, 604–613. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Dopaminergic System | Localization of Neuronal Cell Bodies | Localization of Nerve Endings | Functions |
|---|---|---|---|
| nigrostriatal | substantia nigra (A9 group) | striatum | motor and posture control |
| mesolimbic | ventral tegmental area (A10 group) | nucleus accumbens | motivation, arousal, reward |
| mesocortical | ventral tegmental area | medial prefrontal cortex | motivation, attention, decision making |
| incertohypthalamic | hypothalamus (dopaminergic A13–A14 groups) | hypothalamic nuclei (PVN) and medial preoptic area | different functions (feeding, erectile function, sexual behavior) |
| tuberoinfundibular | hypothalamus-arcuate nucleus | median eminence | inhibition of prolactin release by pituitary lactotropes |
| Dopamine Receptor Families | Transduction Mechanisms | Receptor Subtypes | Role in Penile Erection | Role in Copulatory Behavior |
|---|---|---|---|---|
| D1 | increased activity of adenylate cyclase-cAMP signaling pathway, increased PIP2 turnover, increased Ca2+ mobilization | D1 | none (but see [48]) | none (but see [49,50]) |
| D5 | not available | not available | ||
| D2 | decreased activity of adenylate cyclase-cAMP signaling pathway, increased K+ channels activation, increased voltage-gated Ca2+ channels activation | D2s–D2l | facilitatory | facilitatory |
| D3 | none (but see [51]) | facilitatory * | ||
| D4 | facilitatory (but see [51,52]) | facilitatory |
| Binding Affinities at | References | |||
|---|---|---|---|---|
| D2 Receptors | D3 Receptors | D4 Receptors | ||
| Ki Values (nM) | ||||
| D2-like receptor agonists | ||||
| apomorphine | 32 | 26 | 2.6 | [55] |
| quinpirole | 1.8 | 0.96 | 3.0 | [56] |
| pramipexole | 3.9 | 0.5 | 5.1 | [56] |
| PD 128,907 | 931 | 9.7 | 2430 | [51] |
| PNU-95,666E | 144 | 493 | >10,000 | [51] |
| PD 168,077 | 3740 | 2810 | 8.7 | [57] |
| ABT-724 | >10,000 | NR | 63.6 | [58] |
| PIP-3EA | 990 | 3900 | 2.8 | [59] |
| FAUC 3019 | 33 | 82 | 0.4 | [60] |
| FAUC 389 | 210 | 390 | 1.5 | [61] |
| A-412997 | 2848 | 2095 | 7.9 | [62] |
| CP 226269 | 1760 | NR | 6.0 | [63] |
| D2-like receptor antagonists | ||||
| L-741,626 | 2.4 | 100 | 220 | [64] |
| SB277011A | 1000 | 10 | not available | [65] |
| FAUC 365 | 3600 | 0.5 | 340 | [66] |
| L-745,870 | 960 | 2300 | 0.43 | [64] |
| haloperidol | 6.3 | 6.1 | 10 | [67] |
| raclopride | 1.0 | 1.3 | 5070 | [68] |
| Dopamine Receptor Agonist | Dopamine Receptor Involved | Effect on Penile Erection | Effect on Copulatory Behavior | References |
|---|---|---|---|---|
| Apomorphine | D1/D2/D3/D4/D5 | facilitatory | facilitatory | [46,47,55,69,70] |
| SKF 38393 | D1 | none | not available | [46,71,72,73] |
| Quinpirole | D2/D3/D4 | facilitatory | facilitatory | [51,69] |
| Pramipexole | D3/D2 | facilitatory | facilitatory | [51,74,75,76,77] |
| PD 128,907 | D3/D2 | facilitatory | not available | [51,52] |
| PNU-95666E | D2 | facilitatory | not available | [51] |
| PD 168,077 | D4 | facilitatory | facilitatory | [55,74,75,78,79,80,81] |
| ABT-724 | D4 | facilitatory | facilitatory | [58,70] |
| PIP3EA | D4 | facilitatory | not available | [59,79] |
| FAUC 3019 | D4 | facilitatory | not available | [60] |
| FAUC 389 | D4 | facilitatory | not available | [61] |
| A-412997 | D4 | facilitatory | not available | [62] |
| CP 226269 | D4 | facilitatory | not available | [55] |
| Pre-Treatment | Treatment | |||
|---|---|---|---|---|
| Saline | Apomorphine | Pramipexole | PD 168,077 | |
| Effect on Penile Erection | ||||
| Saline | == | ↑ | ↑ | ↑ |
| L741,626 | == | ↓ | ↓ | ↑ |
| SB277011A | == | ↑ | ↑ | ↑ |
| FAUC365 | == | ↑ | ↑ | ↑ |
| L745,870 | == | ↑ | ↑ | ↓ |
| Raclopride | == | ↓ | ↓ | ↑ |
| Pre-Treatment | Treatment | |||||
|---|---|---|---|---|---|---|
| Apomorphine | PD 168,077 | ABT 724 | ||||
| Effect on | ||||||
| Penile Erection | NO Production | Penile Erection | NO Production | Penile Erection | NO Production | |
| Saline | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ |
| L741,626 | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ |
| L745,870 | ↑ | ↑ | ↓ | ↓ | ↓ | ↓ |
| Oxy-Ant | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ |
| Oxy-Ant I.C.V. | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ |
| L-NAME | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ |
| ω-Conotoxin | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ |
| Pretreatment | Treatment | Penile Erection | Dopamine Release in the NAc |
|---|---|---|---|
| Vehicle | Oxytocin into the VTA, VS, or Am | ↑ | ↑ |
| Oxytocin antagonist into the VTA | Oxytocin into the VTA, VS, or Am | ↓ | ↓ |
| Dizolcipine into the VTA | Oxytocin into the VS and Am | ↓ | ↓ |
| Haloperidol into the NAc | Oxytocin into the VTA | ↓ | ↑ |
| Vehicle | Apomorphine into the PVN | ↑ | ↑ |
| Vehicle | PD 168,077 into the PVN | ↑ | ↑ |
| Oxytocin antagonist into the VTA | Apomorphine into the PVN | ↓ | ↓ |
| Oxytocin antagonist into the VTA | PD 168,077 into the PVN | ↓ | ↓ |
| Treatments | ML | IL | EL | MF | IF | EF | PEI |
|---|---|---|---|---|---|---|---|
| PD 168,077 | == | == | == | ↓ | == | ↑ | ↓ |
| ABT-724 | == | == | == | ↓ | == | ↑ | ↓ |
| Apomorphine | == * | == * | ↓ | ↓ | ↓ | ↑ | == |
| Treatments | ML | IL | EL | MF | IF | EF | PEI |
|---|---|---|---|---|---|---|---|
| L-745,870 | == | ↑ | ↑ | ↑ | == | ↓ | ↑ |
| Haloperidol | ↑ | ↑ | ↑ | == | ↓ | ↓ | ↑ |
| Pretreatment | Treatment | ML | IL | EL | MF | IF | EF | PEI |
|---|---|---|---|---|---|---|---|---|
| Vehicle | PD 168,077 | == | == | == | ↓ | == | ↑ | ↓ |
| L-745,870 | Vehicle | == | ↑ | ↑ | ↑ | == | ↓ | ↑ |
| L-745,870 | PD 168,077 | == | == | ↑ | == | == | == | == |
| Vehicle | ABT 724 | == | == | == | ↓ | == | ↑ | ↓ |
| L-745,870 | ABT 724 | == | == | ↑ | == | == | == | == |
| Vehicle | Apomorphine | == * | == * | ↓ | ↓ | ↓ | ↑ | == |
| L-745,870 | Apomorphine | == | ↑ | ↓ | ↓ | ↓ | ↑ | == |
| Psychogenetically Selected Rat Sub-Lines | ||||||||
|---|---|---|---|---|---|---|---|---|
| Copulatory Parameters | RHA | RLA | NEHR | NELR | ||||
| Naive | Experienced | Naive | Experienced | Naive | Experienced | Naive | Experienced | |
| NCPE | 2.0 | 2.4 * | 1.1 # | 1.3 # | na | na | na | na |
| ML | 290 | 25 * | 600 # | 400 * # | 180 | 20 * | 410 # | 200 * # |
| IL | 320 | 30 * | 620 # | 350 * # | 190 | 20 * | 520 # | 200 * |
| EL | 1200 | 500 * | 1280 | 980 * # | 830 | 570 * | 750 | 580 * |
| MF | 12 | 6 * | 7 # | 6 | 18 | 11 * | 10 # | 9 |
| IF | 20 | 20 | 10 | 10 | 15 | 15 | 10 | 9 |
| PEI | 360 | 300 * | 480 | 450 # | 280 | 330 | 330 | 320 |
| EF | 1.3 | 3.2 * | 1 | 1.7 * # | na | na | na | na |
| III | 62 | 30 * | 120 # | 76 * # | 58 | 40 * | 85 # | 60 * # |
| CE | 0.60 | 0.78 * | 0.57 | 0.68 * # | 0.60 | 0.65 | 0.55 | 0.57 # |
| Rat Sub-Lines | Sexual Condition | Nucleus Accumbens Extracellular Dopamine (nM) | Medial Prefrontal Cortex Extracellular Dopamine (nM) |
|---|---|---|---|
| RHA | Naive | 1.9 | 0.8 |
| Experienced | 2.0 | 1.2 | |
| RLA | Naive | 1.9 | 0.7 |
| Experienced | 1.8 | 1.1 |
| Rat Sub-Lines | Sexual Condition | Area under the Curve of Extracellular Dopamine Values in the Nucleus Accumbens Dialysate | Area under the Curve of Extracellular Dopamine Values in the Medial Prefrontal Cortex dialysate |
|---|---|---|---|
| RHA | Naive | 281 | 210 |
| Experienced | 256 | 315 * | |
| RLA | Naive | 95 # | 154 # |
| Experienced | 164 * | 252 * |
| Psychogenetically Selected Rat Sub-Lines | ||||||
|---|---|---|---|---|---|---|
| Copulatory Parameters | LY | HY | SD | |||
| Naive | Experienced | Naive | Experienced | Naive | Experienced | |
| ML | 683 | 93 * | 694 | 140 * | 400 | 25 * |
| IL | 2590 | 187 * | 1499 | 226 * | 500 | 30 * |
| EL | 2655 | 345 * | 2700 | 520 * | 1200 | 520 * |
| MF | na | 17 | na | 13 | 15 | 6 * |
| IF | na | 10 | na | 8 | 21 | 10 * |
| PEI | na | 300 * | na | 380 * | 470 | 360 |
| EF | na | na | na | na | 1 | 2.7 * |
| III | na | 70 | na | 120 # | 60 | 30 * |
| CE | na | 0.59 | na | 0.45 # | 0.60 | 0.72 * |
| Rat Sub-Lines | ||||||
|---|---|---|---|---|---|---|
| Copulatory Parameters | DAT-KO | HET | WT | |||
| Naive | Experienced | Naive | Experienced | Naive | Experienced | |
| NCPE | 0.5 # | 0.5 # | 1.8 | 3 * # | 1 | 1.2 |
| ML | 180 | 20 * | 200 | 20 * | 560 | 50 * |
| IL | 210 | 17 * | 360 | 26 * | 580 | 40 * |
| EL | 980 | 445 * | 1320 | 920 | 1420 | 1020 * |
| MF | 10 | 7 * | 20 | 20 | 14 | 13 |
| IF | 10 | 9 | 11 | 12 | 10 | 14 |
| PEI | 250 # | 260 # | 480 | 280 * | 470 | 360 |
| EF | 2.2 | 3.5 * | 1.3 | 2.7 * | 1.1 | 2.4 * |
| III | 350 # | 30 * | 360 | 40 * | 700 | 50 * |
| CE | 0.50 # | 0.56 * # | 0.46 | 0.48 | 0.41 | 0.52 * |
| Rat Sub-Lines | Sexual Condition | Extracellular Dopamine (nM) |
|---|---|---|
| WT | Naive | 1.8 |
| Experienced | 2.0 | |
| HET | Naive | 4.5 * |
| Experienced | 5.3 * | |
| DAT-KO | Naive | 17 * # |
| Experienced | 19 * # |
| Rat Sub-Lines | Area under the Curve of Extracellular Dopamine Values |
|---|---|
| WT | 300 |
| HET | 680 * |
| DAT-KO | 2200 * # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melis, M.R.; Sanna, F.; Argiolas, A. Dopamine, Erectile Function and Male Sexual Behavior from the Past to the Present: A Review. Brain Sci. 2022, 12, 826. https://doi.org/10.3390/brainsci12070826
Melis MR, Sanna F, Argiolas A. Dopamine, Erectile Function and Male Sexual Behavior from the Past to the Present: A Review. Brain Sciences. 2022; 12(7):826. https://doi.org/10.3390/brainsci12070826
Chicago/Turabian StyleMelis, Maria Rosaria, Fabrizio Sanna, and Antonio Argiolas. 2022. "Dopamine, Erectile Function and Male Sexual Behavior from the Past to the Present: A Review" Brain Sciences 12, no. 7: 826. https://doi.org/10.3390/brainsci12070826
APA StyleMelis, M. R., Sanna, F., & Argiolas, A. (2022). Dopamine, Erectile Function and Male Sexual Behavior from the Past to the Present: A Review. Brain Sciences, 12(7), 826. https://doi.org/10.3390/brainsci12070826