
The Higher, More Complicated: The Neural Mechanism of Hierarchical Task Switching on Prefrontal Cortex

{kind=link}
Abstract
:1. Introduction
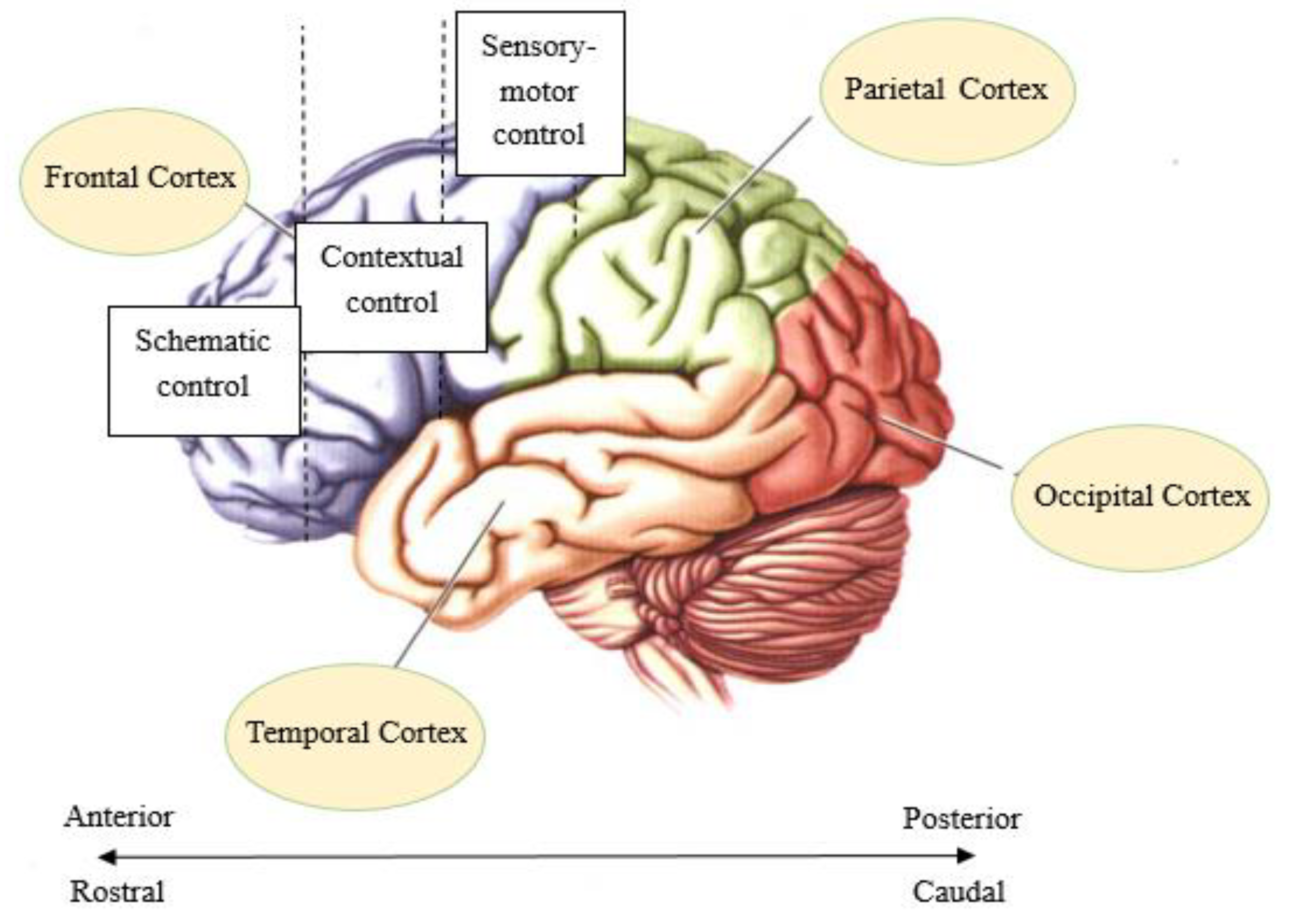
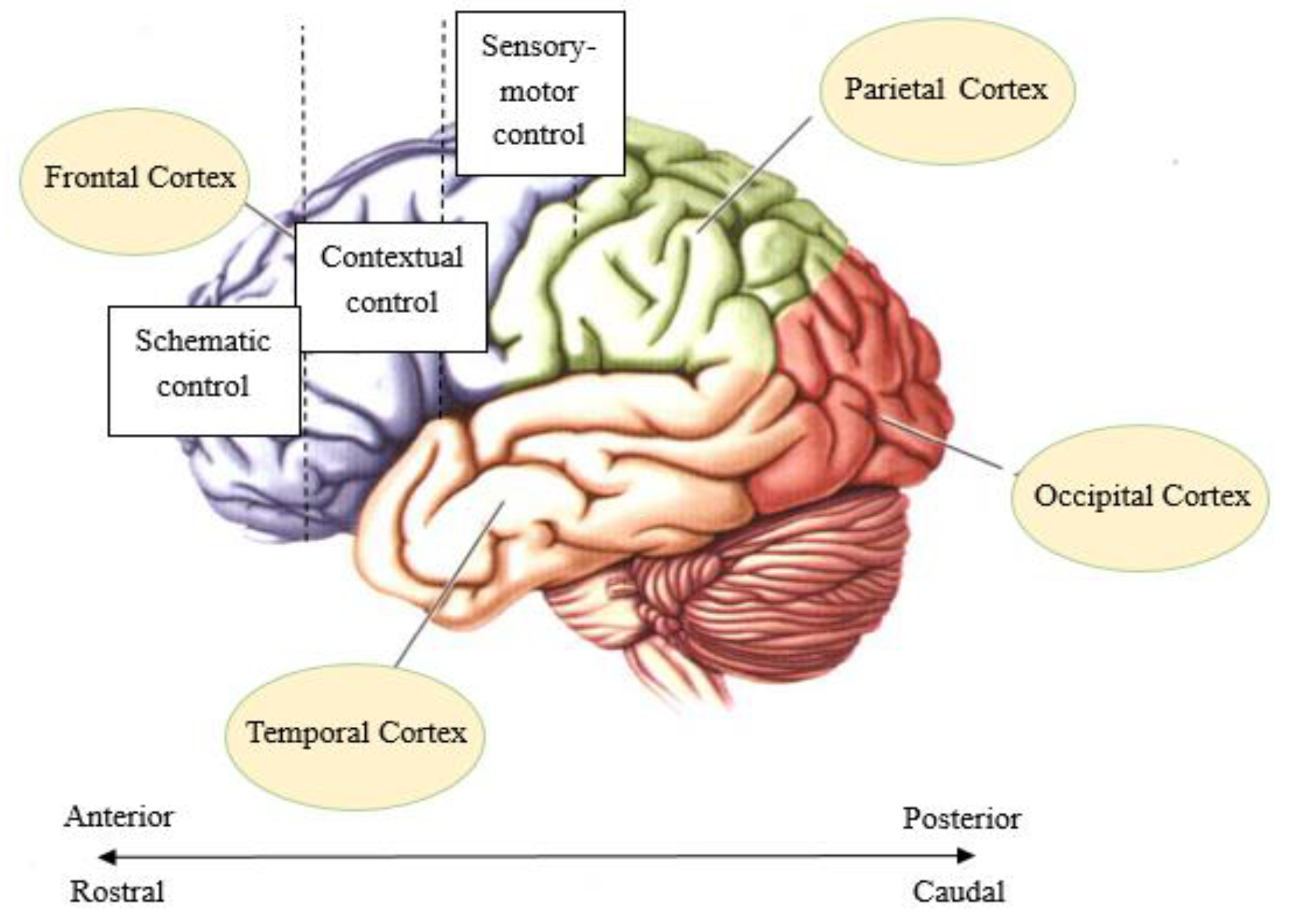
2. Hierarchy of Rule Representation in the Prefrontal Cortex
3. Cognitive Control and Task Switching
3.1. Switching Cost and Its Theoretical Explanations
3.2. Prefrontal Cortex Related to Task Switching
3.3. EEG Markers of Neural Mechanisms of Task Switching
4. Hierarchical Process Mechanisms of Task Switching
4.1. The Generation Forms of Hierarchical Tasks
4.2. Hierarchical Network Processing Models of Task Switching
4.3. Brain Mechanisms of Hierarchical Task Switching
5. Conclusions and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Vandierendonck, A.; Liefooghe, B.; Verbruggen, F. Task switching: Interplay of reconfiguration and interference control. Psychol. Bull. 2010, 136, 601. [Google Scholar] [CrossRef] [PubMed]
- Kleinsorge, T.; Scheil, J. Further evidence for functional differences between guessing versus choosing an upcoming task. Acta Psychol. 2018, 183, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Capizzi, M.; Ambrosini, E.; Arbula, S.; Vallesi, A. Brain oscillatory activity associated with switch and mixing costs during reactive control. Psychophysiology 2020, 57, e13642. [Google Scholar] [CrossRef]
- Liu, H.; Liang, L.; Dunlap, S.; Fan, N.; Chen, B. The effect of domain-general inhibition-related training on language switching: An ERP study. Cognition 2016, 146, 264–276. [Google Scholar] [CrossRef]
- Grange, J.A.; Kowalczyk, A.W.; O’Loughlin, R. The Effect of Episodic Retrieval on Inhibition in Task Switching. J. Exp. Psychol. Hum. Percept. Perform. 2017, 43, 1568. [Google Scholar] [CrossRef]
- Badre, D. Cognitive control, hierarchy, and the rostro–caudal organization of the frontal lobes. Trends Cogn. Sci. 2008, 12, 193–200. [Google Scholar] [CrossRef]
- Bahlmann, J.; Blumenfeld, R.S.; D’Esposito, M. The Rostro-Caudal Axis of Frontal Cortex Is Sensitive to the Domain of Stimulus Information. Cereb. Cortex 2015, 25, 1815–1826. [Google Scholar] [CrossRef] [Green Version]
- Kleinsorge, T.; Heuer, H. Hierarchical switching in a multi-dimensional task space. Psychol. Res. 1999, 62, 300–312. [Google Scholar] [CrossRef]
- Collins, A.G.; Frank, M.J. Cognitive control over learning: Creating, clustering, and generalizing task-set structure. Psychol. Rev. 2013, 120, 190. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.G.; Frank, M.J. Neural signature of hierarchically structured expectations predicts clustering and transfer of rule sets in reinforcement learning. Cognition 2016, 152, 160. [Google Scholar] [CrossRef] [Green Version]
- Werchan, D.M.; Collins, A.G.; Frank, M.J.; Amso, D. Role of Prefrontal Cortex in Learning and Generalizing Hierarchical Rules in 8-Month-Old Infants. J. Neurosci. 2016, 36, 10314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.G.; Cavanagh, J.F.; Frank, M.J. Human EEG uncovers latent generalizable rule structure during learning. J. Neurosci. 2014, 34, 4677–4685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donoso, M.; Collins, A.G.; Koechlin, E. Human cognition. Foundations of human reasoning in the prefrontal cortex. Science 2014, 344, 1481. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.J.; Badre, D. Mechanisms of hierarchical reinforcement learning in corticostriatal circuits 1: Computational analysis. Cereb. Cortex 2011, 22, 509–526. [Google Scholar] [CrossRef] [Green Version]
- Botvinick, M.M. Hierarchical models of behavior and prefrontal function. Trends Cogn. Sci. 2008, 12, 201. [Google Scholar] [CrossRef] [Green Version]
- Badre, D.; D’Esposito, M. Functional Magnetic Resonance Imaging Evidence for a Hierarchical Organization of the Prefrontal Cortex. J. Cognit. Neurosci. 2007, 19, 2082. [Google Scholar] [CrossRef] [Green Version]
- Nee, D.E.; Brown, J.W. Rostral-Caudal Gradients of Abstraction Revealed by Multi-Variate Pattern Analysis of Working Memory. NeuroImage 2012, 63, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Badre, D.; Nee, D.E. Frontal Cortex and the Hierarchical Control of Behavior. Trends Cogn. Sci. 2017, 22, 170–188. [Google Scholar] [CrossRef]
- Evan, N.D.; Andrew, J.; Brown, J.W. Prefrontal Cortex Organization: Dissociating Effects of Temporal Abstraction, Relational Abstraction, and Integration with fMRI. Cereb. Cortex 2013, 24, 2377–2387. [Google Scholar]
- Koechlin, E.; Ody, C.; Kouneiher, F. The Architecture of Cognitive Control in the Human Prefrontal Cortex. Science 2003, 302, 1181–1185. [Google Scholar] [CrossRef] [Green Version]
- Koechlin, E.; Summerfield, C. An information theoretical approach to prefrontal executive function. Trends Cogn. Sci. 2007, 11, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Monsell, S. Task Switching. Trends Cogn. Sci. 2003, 7, 134–140. [Google Scholar] [CrossRef]
- Schneider, D.W. Phasic Alertness and Residual Switch Costs in Task Switching. J. Exp. Psychol. Hum. Percept. Perform. 2016, 43, 317. [Google Scholar] [CrossRef] [PubMed]
- Proskovec, A.; Wiesman, A.; Wilson, T. The strength of alpha and gamma oscillations predicts behavioral switch costs. NeuroImage 2019, 188, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.W.; Foxe, J.J.; Molholm, S. Neuro-oscillatory mechanisms of intersensory selective attention and task switching in school-aged children, adolescents and young adults. Dev. Sci. 2016, 19, 469–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, Y.C.; Egner, T. Cueing Cognitive Flexibility: Item-Specific Learning of Switch Readiness. J. Exp. Psychol. Hum. Percept. Perform. 2017, 43, 1950. [Google Scholar] [CrossRef] [PubMed]
- Seibold, J.; Nolden, S.; Oberem, J.; Fels, J.; Koch, I. Intentional preparation of auditory attention-switches: Explicit cueing and sequential switch-predictability. Q. J. Exp. Psychol. 2018, 71, 1382–1395. [Google Scholar] [CrossRef]
- Kiesel, A.; Steinhauser, M.; Wendt, M.; Falkenstein, M.; Jost, K.; Philipp, A.M.; Koch, I. Control and interference in task switching—A review. Psychol. Bull. 2010, 136, 849–874. [Google Scholar] [CrossRef] [Green Version]
- Meiran, N. Reconfiguration of processing mode prior to task performance. J. Exp. Psychol. Learn. Mem. Cogn. 1996, 22, 1423–1442. [Google Scholar] [CrossRef]
- Rogers, R.D.; Monsell, S. Costs of a predictible switch between simple cognitive tasks. J. Exp. Psychol. Gen. 1995, 124, 207–231. [Google Scholar] [CrossRef]
- Crone, E.A.; Wendelken, C.; Donohue, S.E.; Bunge, A.S. Neural evidence for dissociable components of task-switching. Cereb. Cortex 2006, 16, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Ravizza, S.M.; Carter, C.S. Shifting set about task switching: Behavioral and neural evidence for distinct forms of cognitive flexibility. Neuropsychologia 2008, 46, 2924–2935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braver, T.S.; Reynolds, J.R.; Donaldson, D.I. Neural mechanisms of transient and sustained cognitive control during task switching. Neuron 2003, 39, 713–726. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Cilles, S.E.; Johnson, N.F.; Gold, B.T. Domain general and domain preferential brain regions associated with different types of task switching: A meta-analysis. Hum. Brain Mapp. 2015, 33, 130–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallesi, A.; Visalli, A.; Gracia-Tabuenca, Z.; Tarantino, V.; Pini, L. Fronto-parietal homotopy in resting-state functional connectivity predicts task-switching performance. Brain Struct. Funct. 2022, 227, 655–672. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, K.; Aoki, R.; Takeda, M.; Nakahara, K.; Jimura, K. Cross-Hemispheric Complementary Prefrontal Mechanisms during Task Switching under Perceptual Uncertainty. J. Neurosci. 2021, 41, 2197–2213. [Google Scholar] [CrossRef]
- Sua, B.; Nma, D.; Sha, D.; Sy, C.; Ena, D. Involvement of human left frontoparietal cortices in neural processes associated with task-switching between two sequences of skilled finger movements. Brain Res. 2019, 1722, 146365. [Google Scholar]
- Cooper, P.S.; Karayanidis, F.; McKewen, M.; McLellan-Hall, S.; Wong, A.S.; Skippen, P.; Cavanagh, J.F. Frontal theta predicts specific cognitive control-induced behavioural changes beyond general reaction time slowing. NeuroImage 2019, 189, 130–140. [Google Scholar] [CrossRef]
- Cooper, P.S.; Wong, A.S.; Fulham, W.R.; Thienel, R.; Mansfield, E.; Michie, P.T.; Karayanidis, F. Theta frontoparietal connectivity associated with proactive and reactive cognitive control processes. NeuroImage 2015, 108, 354–363. [Google Scholar] [CrossRef]
- Mansfield, E.L.; Karayanidis, F.; Cohen, M.X. Switch-related and general preparation processes in task-switching: Evidence from multivariate pattern classification of EEG data. J. Neurosci. 2012, 32, 18253–18258. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Wall, H.J.; Hommel, B. Action-effect sharing induces task-set sharing in joint task switching. Cognition 2017, 165, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Wall, H.J.; Hommel, B. The roles of action selection and actor selection in joint task settings. Cognition 2019, 182, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, P.S.; Wong, A.S.; McKewen, M.; Michie, P.T.; Karayanidis, F. Frontoparietal theta oscillations during proactive control are associated with goal-updating and reduced behavioral variability. Biol. Psychol. 2017, 129, 253–264. [Google Scholar] [CrossRef] [PubMed]
- López, M.E.; Pusil, S.; Pereda, E.; Maestú, F.; Barceló, F. Dynamic low frequency EEG phase synchronization patterns during proactive control of task switching. NeuroImage 2019, 186, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Braver, T.S. The variable nature of cognitive control: A dual mechanisms framework. Trends Cogn. Sci. 2012, 16, 106–113. [Google Scholar] [CrossRef] [Green Version]
- McKewen, M.; Cooper, P.S.; Wong, A.S.; Michie, P.T.; Sauseng, P.; Karayanidis, F. Task-switching costs have distinct phase-locked and nonphase-locked EEG power effects. Psychophysiology 2020, 57, e13533. [Google Scholar] [CrossRef]
- Foxe, J.J.; Murphy, J.W.; De Sanctis, P. Throwing out the rules: Anticipatory alpha-band oscillatory attention mechanisms during task-set reconfigurations. Eur. J. Neurosci. 2014, 39, 1960–1972. [Google Scholar] [CrossRef]
- Prada, L.; Barceló, F.; Herrmann, C.S.; Escera, C. EEG delta oscillations index inhibitory control of contextual novelty to both irrelevant distracters and relevant task-switch cues. Psychophysiology 2014, 51, 658–672. [Google Scholar] [CrossRef]
- Peng, W.; Hu, L.; Zhang, Z.; Hu, Y. Causality in the association between P300 and alpha event-related desynchronization. PLoS ONE 2012, 7, e34163. [Google Scholar] [CrossRef] [Green Version]
- Schneider, D.W.; Logan, G.D. “Hierarchical control of cognitive processes: Switching tasks in sequences”: Correction to Schneider and Logan (2006). J. Exp. Psychol. Gen. 2006, 135, 623. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Dai, Y.; Xie, L.; Li, F. Brain responses associated with different hierarchical effects on cues and targets during rule shifting. Biol. Psychol. 2018, 134, 52–63. [Google Scholar] [CrossRef]
- Weaver, S.M.; Arrington, C.M. The Effect of Hierarchical Task Representations on Task Selection in Voluntary Task Switching. J. Exp. Psychol. Learn. Mem. Cogn. 2013, 39, 1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lien, M.C.; Ruthruff, E. Task Switching in a Hierarchical Task Structure: Evidence for the Fragility of the Task Repetition Benefit. J. Exp. Psychol. Learn. Mem. Cogn. 2004, 30, 697–713. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Xie, L.; Cao, B.; Li, J.; Chen, Y.; Li, F. More abstract, more difficult to shift: Behavior and electrophysiological evidence. Behav. Brain Res. 2019, 362, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Doñamayor, N.; Münte, T.F.; Bahlmann, J. Event-related potentials and neural oscillations dissociate levels of cognitive control. Behav. Brain Res. 2017, 320, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A.; Wager, T.D. The Unity and Diversity of Executive Functions and Their Contributions to Complex “Frontal Lobe” Tasks: A Latent Variable Analysis. Cogn. Psychol. 2000, 41, 49–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarr, N.; Brown, J.W. Hierarchical error representation in medial prefrontal cortex. NeuroImage 2016, 124, 238–247. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, C.; Han, J. The Higher, More Complicated: The Neural Mechanism of Hierarchical Task Switching on Prefrontal Cortex. Brain Sci. 2022, 12, 645. https://doi.org/10.3390/brainsci12050645
Zhu C, Han J. The Higher, More Complicated: The Neural Mechanism of Hierarchical Task Switching on Prefrontal Cortex. Brain Sciences. 2022; 12(5):645. https://doi.org/10.3390/brainsci12050645
Chicago/Turabian StyleZhu, Chengdong, and Jiahui Han. 2022. "The Higher, More Complicated: The Neural Mechanism of Hierarchical Task Switching on Prefrontal Cortex" Brain Sciences 12, no. 5: 645. https://doi.org/10.3390/brainsci12050645
APA StyleZhu, C., & Han, J. (2022). The Higher, More Complicated: The Neural Mechanism of Hierarchical Task Switching on Prefrontal Cortex. Brain Sciences, 12(5), 645. https://doi.org/10.3390/brainsci12050645