
Behavioral and Immunohistochemical Evidence for Suppressive Effects of Goshajinkigan on Salicylate-Induced Tinnitus in Rats

 ,
,  ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Animals
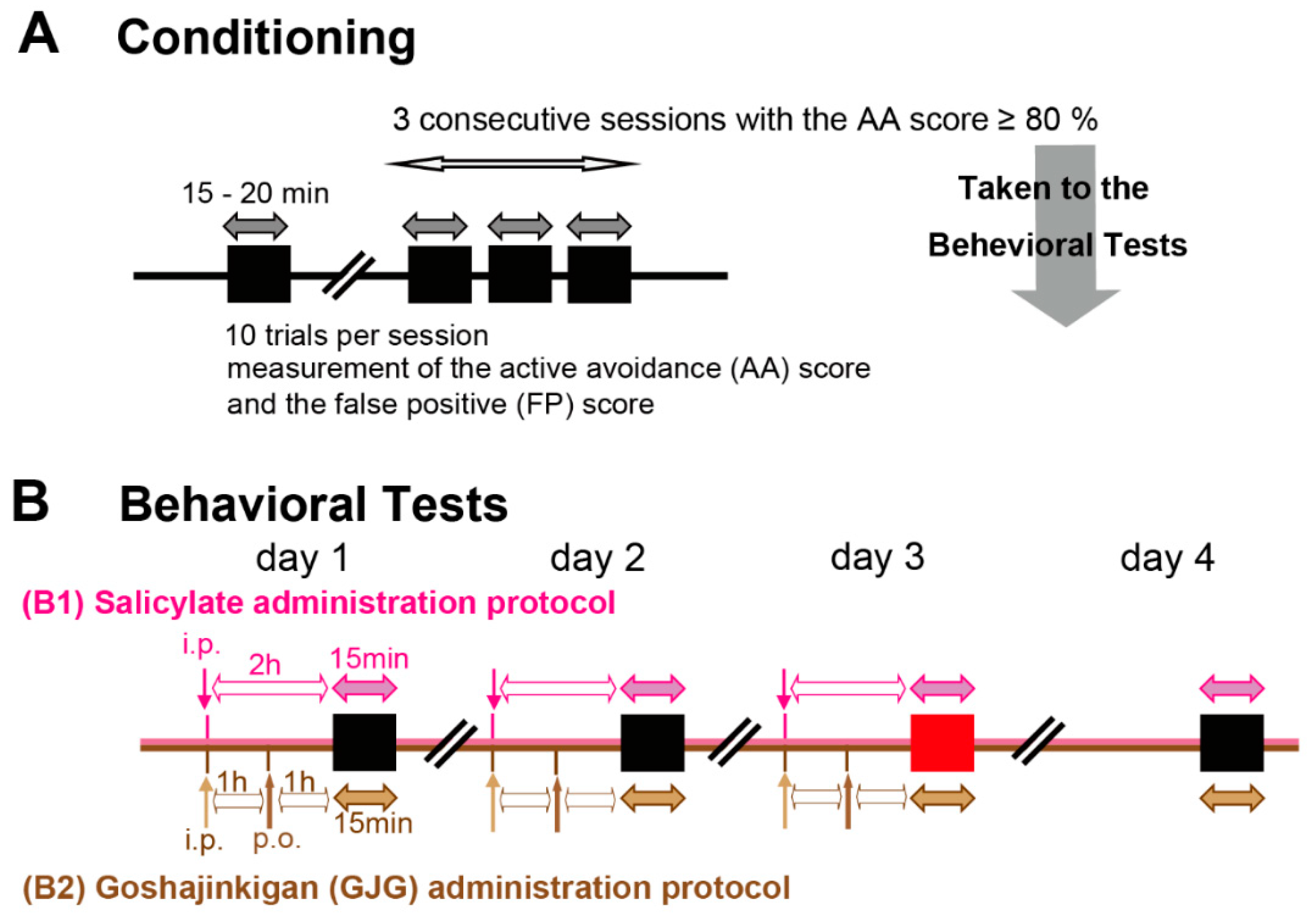
2.2. Behavioral Assessment of Tinnitus
2.3. Immunohistochemical Analysis of c-Fos Expression in the Auditory Pathway
2.4. Observation and Data Analysis of Immunohistochemical Stainings
2.5. Statical Analysis
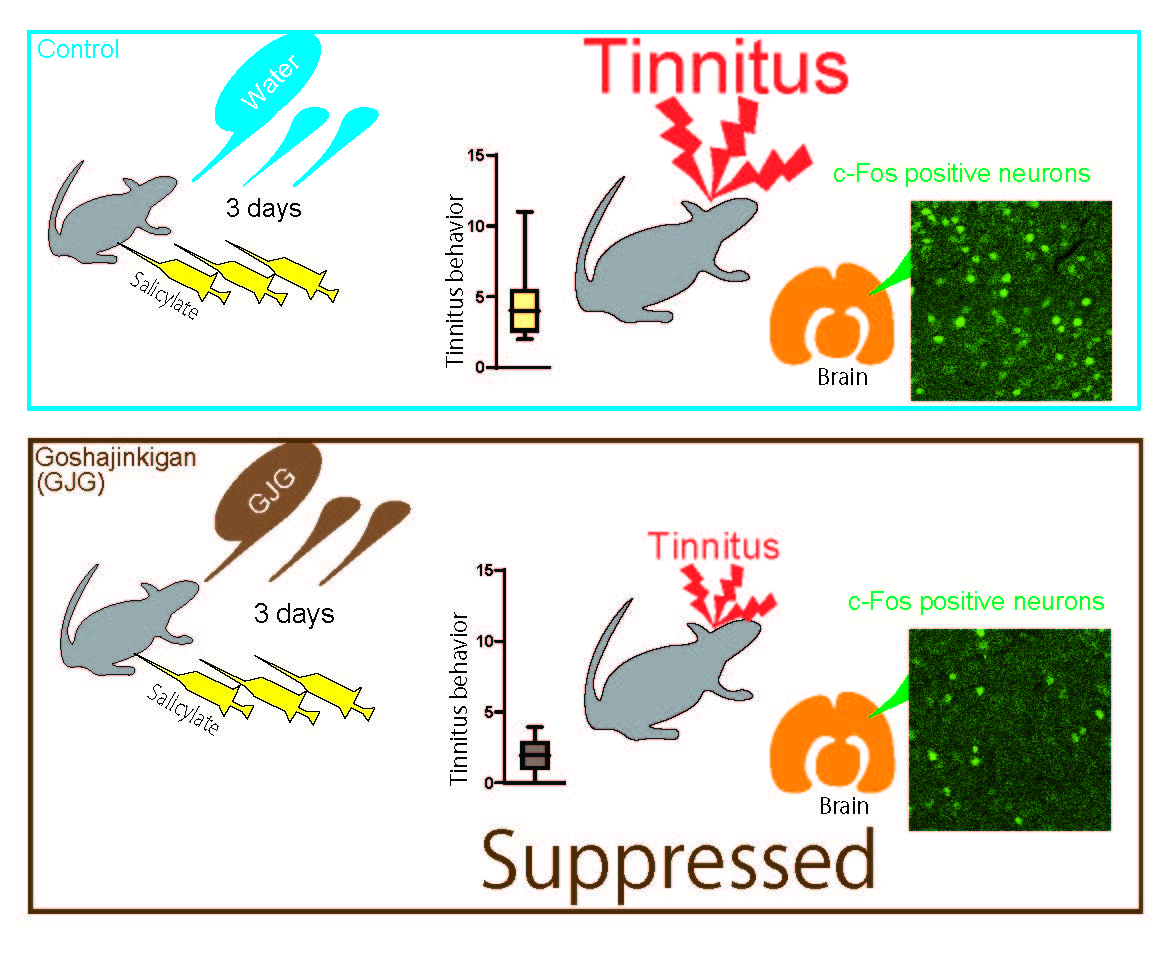
3. Results
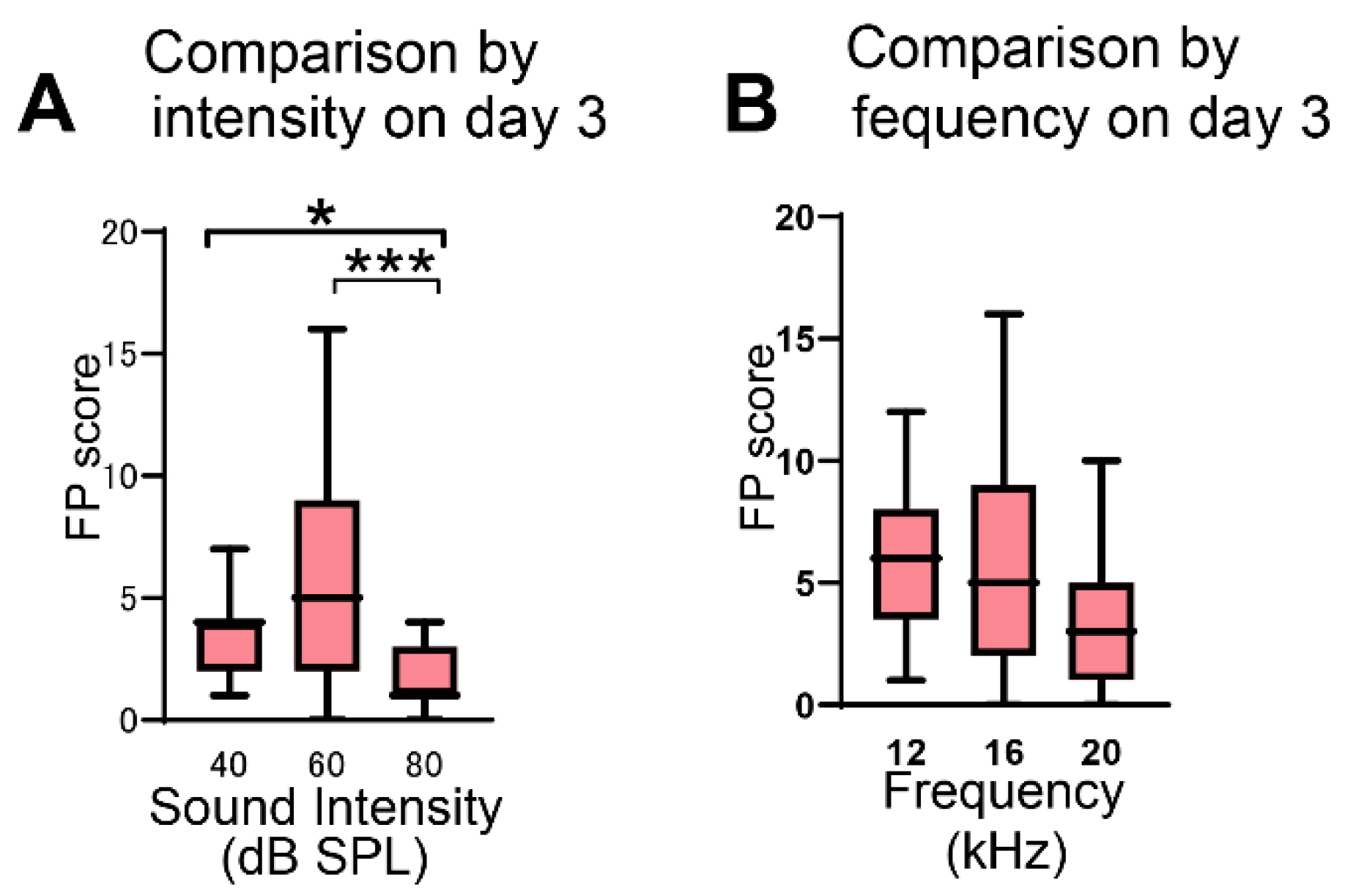
3.1. Effect of Sound Intensity and Frequency on FP
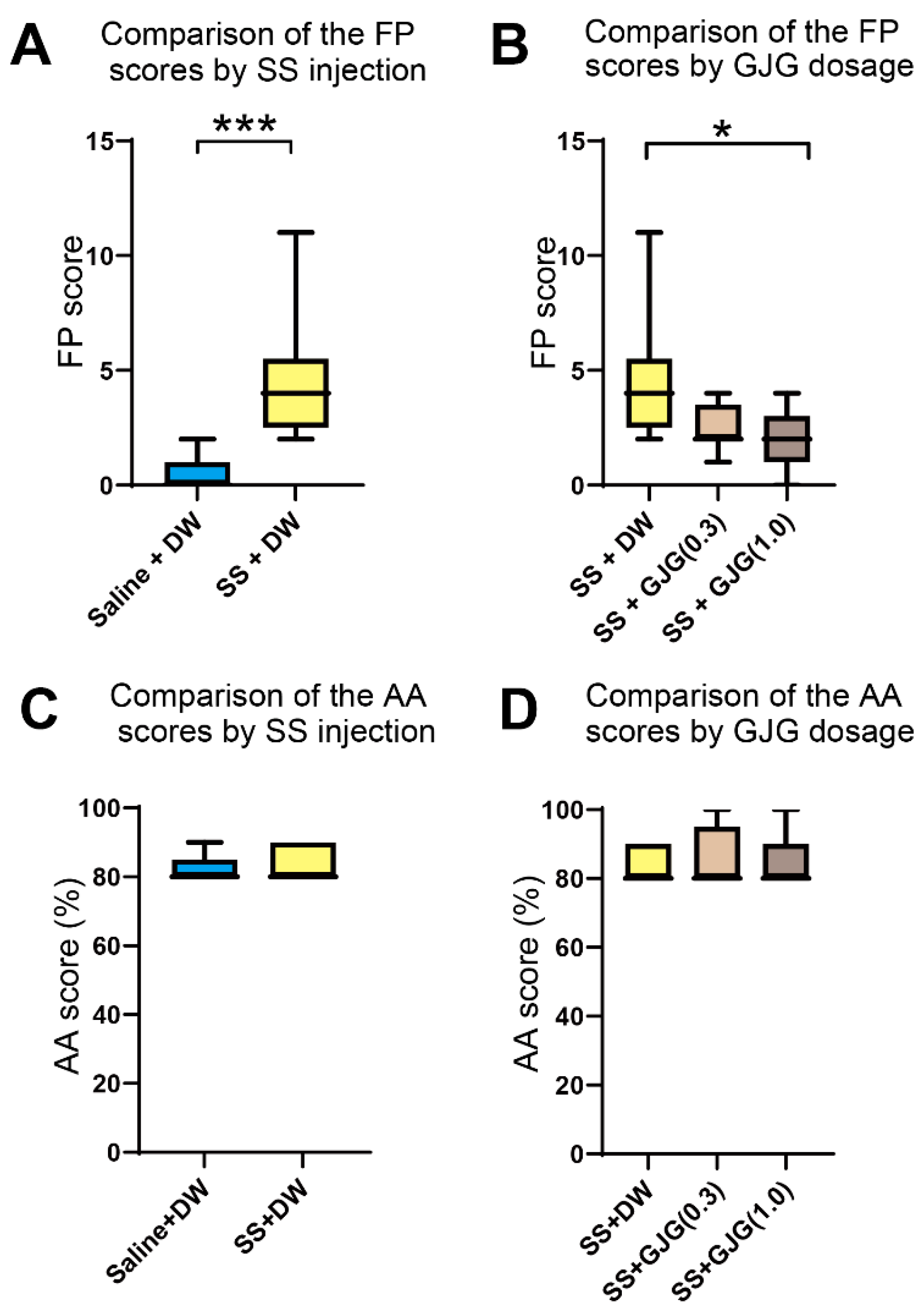
3.2. Effect of GJG on FP scores
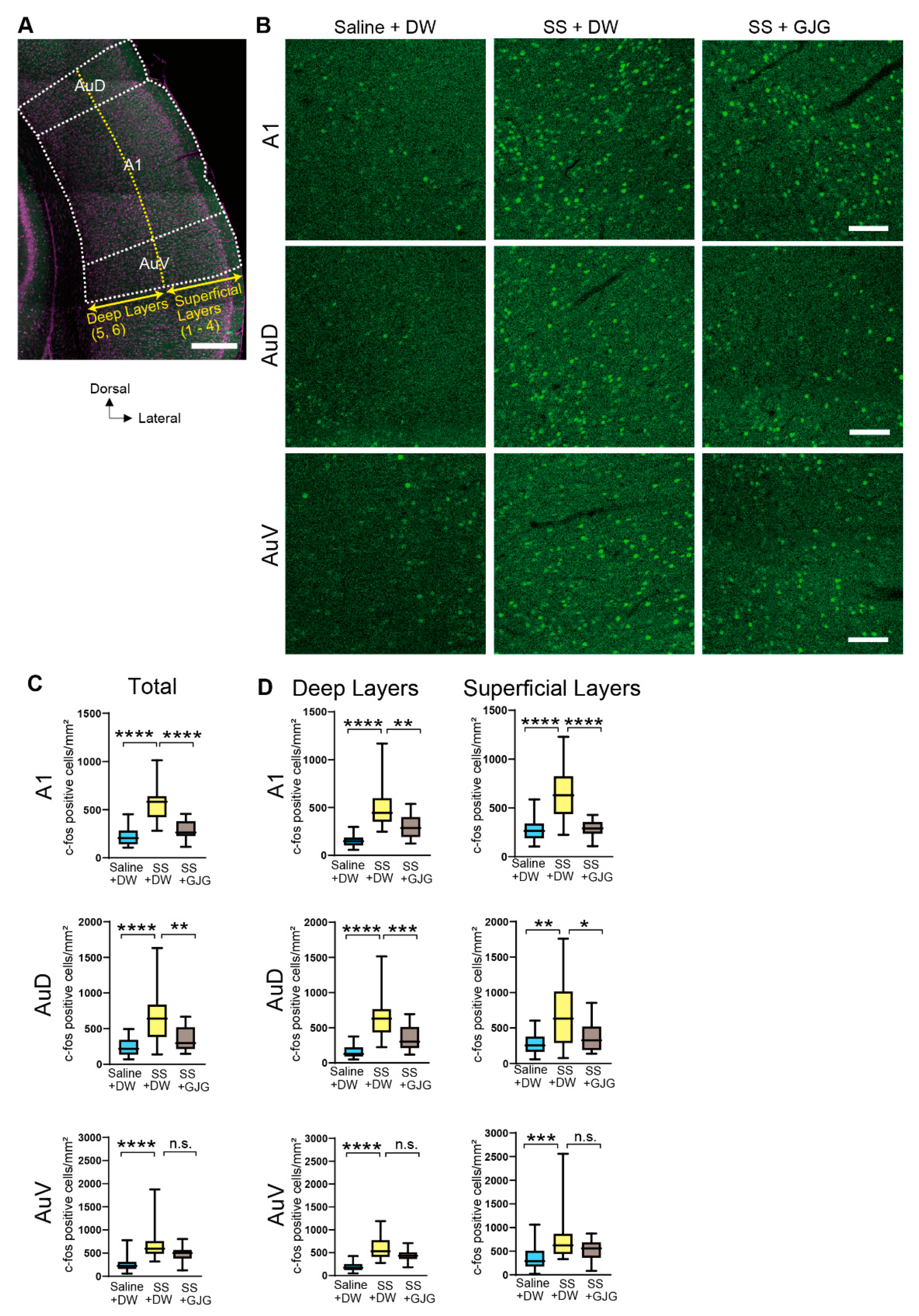
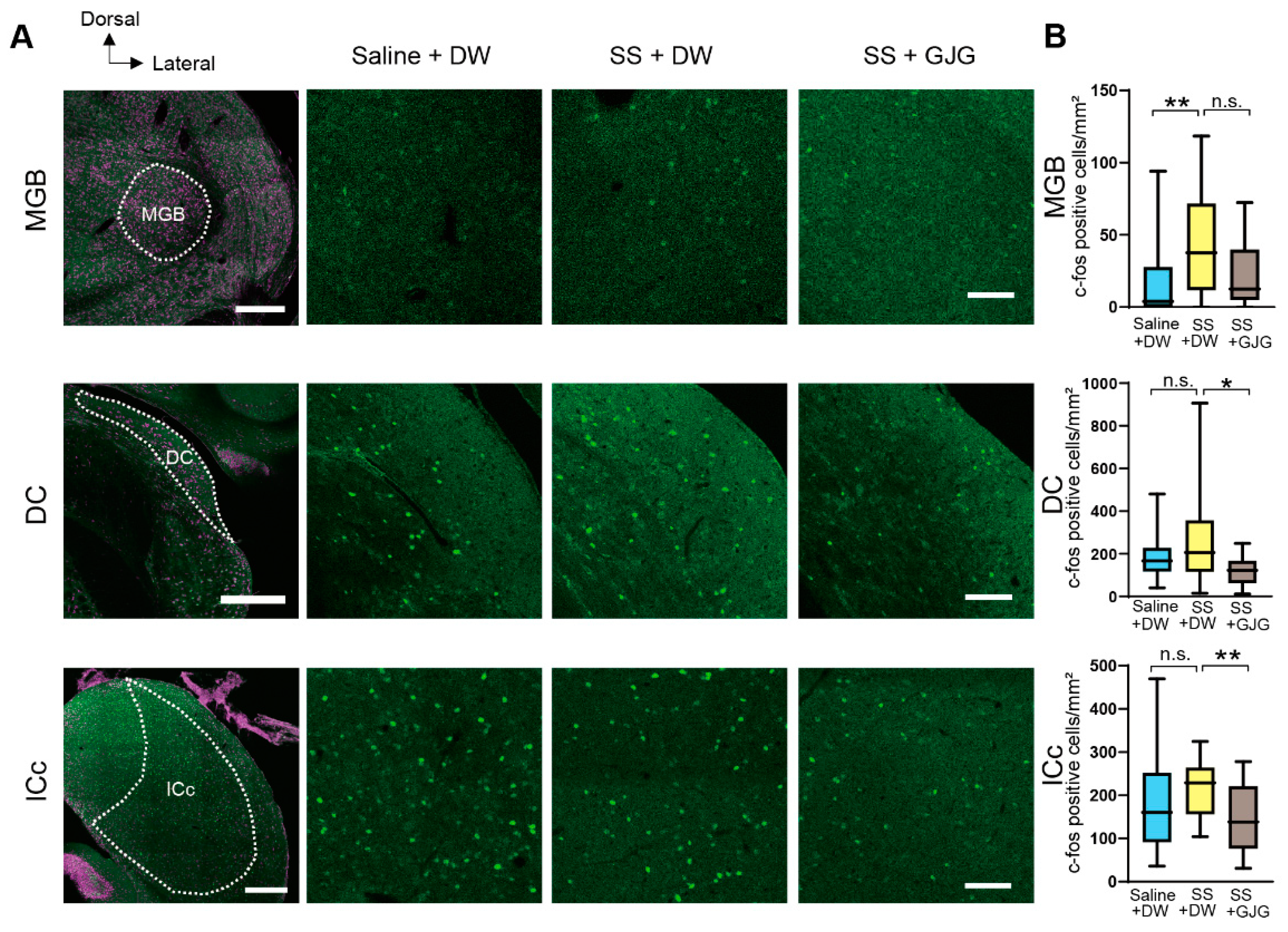
3.3. Effect of GJG on c-Fos Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCormack, A.; Edmondson-Jones, M.; Somerset, S.; Hall, D. A systematic review of the reporting of tinnitus prevalence and severity. Hear. Res. 2016, 337, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Nagata, C.; Nakamura, K.; Kawachi, T.; Takatsuka, N.; Oba, S.; Shimizu, H. Prevalence of tinnitus in community-dwelling Japanese adults. J. Epidemiol. 2011, 21, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Eggermont, J.J.; Roberts, L.E. The neuroscience of tinnitus. Trends Neurosci. 2004, 27, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Eggermont, J.J. Hearing loss, hyperacusis, or tinnitus: What is modeled in animal research? Hear. Res. 2013, 295, 140–149. [Google Scholar] [CrossRef]
- Eggermont, J.J. The auditory cortex and tinnitus—A review of animal and human studies. Eur. J. Neurosci. 2015, 41, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.G. Behavioral measures of tinnitus in laboratory animals. Prog. Brain Res. 2007, 166, 147–156. [Google Scholar] [CrossRef]
- Jastreboff, P.J.; Brennan, J.F.; Coleman, J.K.; Sasaki, C.T. Phantom auditory sensation in rats: An animal model for tinnitus. Behav. Neurosci. 1988, 102, 811–822. [Google Scholar] [CrossRef]
- Jastreboff, P.J.; Brennan, J.F.; Sasaki, C.T. An animal model for tinnitus. Laryngoscope 1988, 98, 280–286. [Google Scholar] [CrossRef]
- Bauer, C.A.; Brozoski, T.J.; Rojas, R.; Boley, J.; Wyder, M. Behavioral model of chronic tinnitus in rats. Otolaryngol. Head Neck Surg. 1999, 121, 457–462. [Google Scholar] [CrossRef]
- Bauer, C.A.; Brozoski, T.J. Assessing tinnitus and prospective tinnitus therapeutics using a psychophysical animal model. J. Assoc. Res. Otolaryngol. 2001, 2, 54–64. [Google Scholar] [CrossRef][Green Version]
- Guitton, M.J.; Caston, J.; Ruel, J.; Johnson, R.M.; Pujol, R.; Puel, J.L. Salicylate induces tinnitus through activation of cochlear NMDA receptors. J. Neurosci. 2003, 23, 3944–3952. [Google Scholar] [CrossRef] [PubMed]
- Uno, T.; Ohsawa, I.; Tokudome, M.; Sato, Y. Effects of Goshajinkigan on insulin resistance in patients with type 2 diabetes. Diabetes Res. Clin. Pract. 2005, 69, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Tawata, M.; Kurihara, A.; Nitta, K.; Iwase, E.; Gan, N.; Onaya, T. The effects of goshajinkigan, a herbal medicine, on subjective symptoms and vibratory threshold in patients with diabetic neuropathy. Diabetes Res. Clin. Pract. 1994, 26, 121–128. [Google Scholar] [CrossRef]
- Usuki, Y.; Usuki, S.; Hommura, S. Successful treatment of a senile diabetic woman with cataract with goshajinkigan. Am. J. Chin. Med. 1991, 19, 259–263. [Google Scholar] [CrossRef]
- Ohnishi, S.; Sawaki, S.; Tsuchiya, K.; Hiiragi, K.; Taniuchi, A.; Furukawa, M.; Wada, H.; Tanaka, Y.; Ito, M.; Bono, K.; et al. Effect of TJ-107 (Tsumura Goshajinkigan) on tinnitus in a multicenter clinical trial. Jibiinkoukatenbo 1994, 37, 371–379. (In Japanese) [Google Scholar]
- Kizawa, K.; Kitahara, T.; Horii, A.; Maekawa, C.; Kuramatsu, T.; Kawashima, T.; Nishiike, S.; Doi, K.; Inohara, H. Behavioral assessment and identification of a molecular marker in a salicylate-induced tinnitus in rats. Neuroscience 2010, 165, 1323–1332. [Google Scholar] [CrossRef]
- Wallhäusser-Franke, E. Salicylate evokes c-fos expression in the brain stem: Implications for tinnitus. Neuroreport 1997, 8, 725–728. [Google Scholar] [CrossRef]
- Wallhäusser-Franke, E.; Mahlke, C.; Oliva, R.; Braun, S.; Wenz, G.; Langner, G. Expression of c-fos in auditory and non-auditory brain regions of the gerbil after manipulations that induce tinnitus. Exp. Brain Res. 2003, 153, 649–654. [Google Scholar] [CrossRef]
- Wu, J.L.; Chiu, T.W.; Poon, P.W. Differential changes in Fos-immunoreactivity at the auditory brainstem after chronic injections of salicylate in rats. Hear. Res. 2003, 176, 80–93. [Google Scholar] [CrossRef]
- Mahlke, C.; Wallhäusser-Franke, E. Evidence for tinnitus-related plasticity in the auditory and limbic system, demonstrated by arg3.1 and c-fos immunocytochemistry. Hear. Res. 2004, 195, 17–34. [Google Scholar] [CrossRef]
- Santos, P.; da Silva, L.E.; Leão, R.M. Specific immediate early gene expression induced by high doses of salicylate in the cochlear nucleus and inferior colliculus of the rat. Braz. J. Otorhinolaryngol. 2017, 83, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Kono, T.; Suzuki, Y.; Miyagi, C.; Omiya, Y.; Miyano, K.; Kase, Y.; Uezono, Y. Goshajinkigan, a traditional Japanese medicine, prevents oxaliplatin-induced acute peripheral neuropathy by suppressing functional alteration of TRP channels in rat. J. Pharm. Sci. 2014, 125, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Goto, K.; Ishige, A.; Komatsu, Y.; Kamei, J. Antinociceptive effect of Gosha-jinki-gan, a Kampo medicine, in streptozotocin-induced diabetic mice. Jpn J. Pharmacol. 1999, 79, 169–175. [Google Scholar] [CrossRef]
- Ushio, S.; Egashira, N.; Sada, H.; Kawashiri, T.; Shirahama, M.; Masuguchi, K.; Oishi, R. Goshajinkigan reduces oxaliplatin-induced peripheral neuropathy without affecting anti-tumour efficacy in rodents. Eur. J. Cancer 2012, 48, 1407–1413. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: Burlington, VT, USA, 2007. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Jastreboff, P.J.; Brennan, J.F. Evaluating the loudness of phantom auditory perception (tinnitus) in rats. Audiology 1994, 33, 202–217. [Google Scholar] [CrossRef]
- Turner, J.G.; Brozoski, T.J.; Bauer, C.A.; Parrish, J.L.; Myers, K.; Hughes, L.F.; Caspary, D.M. Gap detection deficits in rats with tinnitus: A potential novel screening tool. Behav. Neurosci. 2006, 120, 188–195. [Google Scholar] [CrossRef]
- Zheng, Y.; Seung Lee, H.; Smith, P.F.; Darlington, C.L. Neuronal nitric oxide synthase expression in the cochlear nucleus in a salicylate model of tinnitus. Brain Res. 2006, 1123, 201–206. [Google Scholar] [CrossRef]
- Yang, G.; Lobarinas, E.; Zhang, L.; Turner, J.; Stolzberg, D.; Salvi, R.; Sun, W. Salicylate induced tinnitus: Behavioral measures and neural activity in auditory cortex of awake rats. Hear. Res. 2007, 226, 244–253. [Google Scholar] [CrossRef]
- Ralli, M.; Lobarinas, E.; Fetoni, A.R.; Stolzberg, D.; Paludetti, G.; Salvi, R. Comparison of salicylate- and quinine-induced tinnitus in rats: Development, time course, and evaluation of audiologic correlates. Otol. Neurotol. 2010, 31, 823–831. [Google Scholar] [CrossRef]
- Hu, S.S.; Mei, L.; Chen, J.Y.; Huang, Z.W.; Wu, H. Expression of immediate-early genes in the inferior colliculus and auditory cortex in salicylate-induced tinnitus in rat. Eur. J. Histochem. 2014, 58, 2294. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Yin, M.; Wu, C.; Ji, Y.; Zhou, Y. Neuroglial activation in the auditory cortex and medial geniculate body of salicylate-induced tinnitus rats. Am. J. Transl. Res. 2020, 12, 6043–6059. [Google Scholar]
- Didier, A.; Miller, J.M.; Nuttall, A.L. The vascular component of sodium salicylate ototoxicity in the guinea pig. Hear. Res. 1993, 69, 199–206. [Google Scholar] [CrossRef]
- Eggermont, J.J.; Kenmochi, M. Salicylate and quinine selectively increase spontaneous firing rates in secondary auditory cortex. Hear. Res. 1998, 117, 149–160. [Google Scholar] [CrossRef]
- Sun, W.; Lu, J.; Stolzberg, D.; Gray, L.; Deng, A.; Lobarinas, E.; Salvi, R.J. Salicylate increases the gain of the central auditory system. Neuroscience 2009, 159, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, P.; Cao, Y.; Qin, L.; Sato, Y. Salicylate induced neural changes in the primary auditory cortex of awake cats. Neuroscience 2011, 172, 232–245. [Google Scholar] [CrossRef] [PubMed]
- Lanaia, V.; Tziridis, K.; Schulze, H. Salicylate-Induced Changes in Hearing Thresholds in Mongolian Gerbils Are Correlated with Tinnitus Frequency but Not with Tinnitus Strength. Front. Behav. Neurosci. 2021, 15, 698516. [Google Scholar] [CrossRef]
- Chen, Y.C.; Li, X.; Liu, L.; Wang, J.; Lu, C.Q.; Yang, M.; Jiao, Y.; Zang, F.C.; Radziwon, K.; Chen, G.D.; et al. Tinnitus and hyperacusis involve hyperactivity and enhanced connectivity in auditory-limbic-arousal-cerebellar network. Elife 2015, 4, e06576. [Google Scholar] [CrossRef]
- Salvi, R.; Radziwon, K.; Manohar, S.; Auerbach, B.; Ding, D.; Liu, C.; Chen, Y.C.; Chen, G.D. Review: Neural Mechanisms of Tinnitus and Hyperacusis in Acute Drug-Induced Ototoxicity. Am. J. Audiol. 2021, 30, 901–915. [Google Scholar] [CrossRef]
- Kakehata, S.; Santos-Sacchi, J. Effects of salicylate and lanthanides on outer hair cell motility and associated gating charge. J. Neurosci. 1996, 16, 4881–4889. [Google Scholar] [CrossRef]
- Peng, B.G.; Chen, S.; Lin, X. Aspirin selectively augmented N-methyl-D-aspartate types of glutamate responses in cultured spiral ganglion neurons of mice. Neurosci. Lett. 2003, 343, 21–24. [Google Scholar] [CrossRef]
- Ruel, J.; Chabbert, C.; Nouvian, R.; Bendris, R.; Eybalin, M.; Leger, C.L.; Bourien, J.; Mersel, M.; Puel, J.L. Salicylate enables cochlear arachidonic-acid-sensitive NMDA receptor responses. J. Neurosci. 2008, 28, 7313–7323. [Google Scholar] [CrossRef] [PubMed]
- Deer, B.C.; Hunter-Duvar, I. Salicylate ototoxicity in the chinchilla: A behavioral and electron microscope study. J. Otolaryngol. 1982, 11, 260–264. [Google Scholar] [PubMed]
- Jastreboff, P.J.; Hansen, R.; Sasaki, P.G.; Sasaki, C.T. Differential uptake of salicylate in serum, cerebrospinal fluid, and perilymph. Arch. Otolaryngol. Head Neck Surg. 1986, 112, 1050–1053. [Google Scholar] [CrossRef]
- Boettcher, F.A.; Bancroft, B.R.; Salvi, R.J. Concentration of salicylate in serum and perilymph of the chinchilla. Arch. Otolaryngol. Head Neck Surg. 1990, 116, 681–684. [Google Scholar] [CrossRef]
- Rezapour, M.; Moossavi, A. Tinnitus induction in animals and its impact on auditory system structure. Aud. Vestib. Res. 2019, 28, 204–216. [Google Scholar] [CrossRef]
- Stolzberg, D.; Salvi, R.J.; Allman, B.L. Salicylate toxicity model of tinnitus. Front. Syst. Neurosci. 2012, 6, 28. [Google Scholar] [CrossRef]
- Basta, D.; Ernst, A. Effects of salicylate on spontaneous activity in inferior colliculus brain slices. Neurosci. Res. 2004, 50, 237–243. [Google Scholar] [CrossRef]
- Basta, D.; Goetze, R.; Ernst, A. Effects of salicylate application on the spontaneous activity in brain slices of the mouse cochlear nucleus, medial geniculate body and primary auditory cortex. Hear. Res. 2008, 240, 42–51. [Google Scholar] [CrossRef]
- Lu, Y.G.; Tang, Z.Q.; Ye, Z.Y.; Wang, H.T.; Huang, Y.N.; Zhou, K.Q.; Zhang, M.; Xu, T.L.; Chen, L. Salicylate, an aspirin metabolite, specifically inhibits the current mediated by glycine receptors containing alpha1-subunits. Br. J. Pharmacol. 2009, 157, 1514–1522. [Google Scholar] [CrossRef]
- Su, Y.Y.; Luo, B.; Wang, H.T.; Chen, L. Differential effects of sodium salicylate on current-evoked firing of pyramidal neurons and fast-spiking interneurons in slices of rat auditory cortex. Hear. Res. 2009, 253, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.T.; Luo, B.; Zhou, K.Q.; Xu, T.L.; Chen, L. Sodium salicylate reduces inhibitory postsynaptic currents in neurons of rat auditory cortex. Hear. Res. 2006, 215, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Domarecka, E.; Olze, H.; Szczepek, A.J. Auditory Brainstem Responses (ABR) of Rats during Experimentally Induced Tinnitus: Literature Review. Brain Sci. 2020, 10, 901. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Kono, T.; Suzuki, Y.; Mizuno, K.; Miyagi, C.; Omiya, Y.; Sekine, H.; Mizuhara, Y.; Miyano, K.; Kase, Y.; Uezono, Y. Preventive effect of oral goshajinkigan on chronic oxaliplatin-induced hypoesthesia in rats. Sci. Rep. 2015, 5, 16078. [Google Scholar] [CrossRef]
- Jiang, S.; Baba, K.; Okuno, T.; Kinoshita, M.; Choong, C.J.; Hayakawa, H.; Sakiyama, H.; Ikenaka, M.; Nagano, S.; Sasaki, T.; et al. Go-sha-jinki-Gan Alleviates Inflammation in Neurological Disorders via p38-TNF Signaling in the Central Nervous System. Neurotherapeutics 2021, 18, 460–473. [Google Scholar] [CrossRef]
- Krizman, J.; Rotondo, E.K.; Nicol, T.; Kraus, N.; Bieszczad, K.M. Sex differences in auditory processing vary across estrous cycle. Sci. Rep. 2021, 11, 22898. [Google Scholar] [CrossRef]
- Becker, J.B.; Prendergast, B.J.; Liang, J.W. Female rats are not more variable than male rats: A meta-analysis of neuroscience studies. Biol. Sex Differ. 2016, 7, 34. [Google Scholar] [CrossRef]
- Depner, M.; Tziridis, K.; Hess, A.; Schulze, H. Sensory cortex lesion triggers compensatory neuronal plasticity. BMC Neurosci. 2014, 15, 57. [Google Scholar] [CrossRef]
- Tziridis, K.; Ahlf, S.; Jeschke, M.; Happel, M.F.; Ohl, F.W.; Schulze, H. Noise Trauma Induced Neural Plasticity Throughout the Auditory System of Mongolian Gerbils: Differences between Tinnitus Developing and Non-Developing Animals. Front. Neurol. 2015, 6, 22. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| With SS | Without SS | |||
|---|---|---|---|---|
| Intensity (SPL) | Frequency (Hz) | FP on Day 3 Median (IQR) | FP on Day 4 Median (IQR) | p Value |
| 40 dB | 16 k | 4.0 (2.0–4.0) | 1.0 (0.0–3.0) | ** p = 0.0013 |
| 60 dB | 16 k | 5.0 (2.0–9.0) | 2.0 (1.0–4.0) | **** p < 0.0001 |
| 80 dB | 16 k | 1.0 (1.0–3.0) | 0.0 (0.0–1.0) | * p = 0.0313 |
| 60 dB | 12 k | 6.0 (3.5–8.0) | 1.0 (0.5–5.0) | ** p = 0.0065 |
| 60 dB | 20 k | 3.0 (1.0–5.0) | 1.0 (0.0–2.0) | ** p = 0.0076 |
| Area | Group | c-Fos Expressing Cells Median (IQR) | p Value |
|---|---|---|---|
| Saline + DW | 205 (143–286) | **** p < 0.0001 **** p < 0.0001 | |
| A1 total | SS + DW | 583 (442–641) | |
| SS + GJG | 264 (227–381) | ||
| Saline + DW | 218 (134–343) | **** p < 0.0001 ** p = 0.004 | |
| AuD total | SS + DW | 641 (382–837) | |
| SS + GJG | 297 (214–518) | ||
| Saline + DW | 221 (157–308) | **** p < 0.0001 p = 0.059 | |
| AuV total | SS + DW | 595 (484–760) | |
| SS + GJG | 499 (382–567) | ||
| Saline + DW | 264 (187–339) | **** p < 0.0001 **** p < 0.0001 | |
| A1 superficial | SS + DW | 630 (436–825) | |
| SS + GJG | 290 (235–355) | ||
| Saline + DW | 255 (164–378) | ** p = 0.002 * p = 0.021 | |
| AuD superficial | SS + DW | 632 (288–1013) | |
| SS + GJG | 328 (191–521) | ||
| Saline + DW | 290 (176–509) | *** p = 0.0007 p = 0.214 | |
| AuV superficial | SS + DW | 624 (439–869) | |
| SS + GJG | 561 (362–686) | ||
| Saline + DW | 149 (104–186) | **** p < 0.0001 ** p = 0.002 | |
| A1 deep | SS + DW | 445 (352–599) | |
| SS + GJG | 286 (192–401) | ||
| Saline + DW | 127 (92–221) | **** p < 0.0001 *** p = 0.0008 | |
| AuD deep | SS + DW | 630 (431–764) | |
| SS + GJG | 301 (212–511) | ||
| Saline + DW | 171 (122–253) | **** p < 0.0001 p = 0.069 | |
| AuV deep | SS + DW | 531 (408–777) | |
| SS + GJG | 434 (364–509) |
| Area | Group | c-Fos Expressing Cells Median (IQR) | p Value |
|---|---|---|---|
| Saline + DW | 4 (0–28) | ** p = 0.008 p = 0.097 | |
| MGB | SS + DW | 38 (12–72) | |
| SS + GJG | 13 (5–40) | ||
| Saline + DW | 167 (117–229) | p = 0.406 * p = 0.016 | |
| DC | SS + DW | 206 (116–357) | |
| SS + GJG | 123 (62–168) | ||
| Saline + DW | 161 (91–253) | p = 0.126 ** p = 0.010 | |
| ICc | SS + DW | 229 (156–264) | |
| SS + GJG | 138 (76–221) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitano, K.; Yamashita, A.; Sugimura, T.; Okayasu, T.; Sakagami, M.; Osaki, D.; Kitahara, T.; Saito, Y. Behavioral and Immunohistochemical Evidence for Suppressive Effects of Goshajinkigan on Salicylate-Induced Tinnitus in Rats. Brain Sci. 2022, 12, 587. https://doi.org/10.3390/brainsci12050587
Kitano K, Yamashita A, Sugimura T, Okayasu T, Sakagami M, Osaki D, Kitahara T, Saito Y. Behavioral and Immunohistochemical Evidence for Suppressive Effects of Goshajinkigan on Salicylate-Induced Tinnitus in Rats. Brain Sciences. 2022; 12(5):587. https://doi.org/10.3390/brainsci12050587
Chicago/Turabian StyleKitano, Koichi, Akinori Yamashita, Taketoshi Sugimura, Tadao Okayasu, Masaharu Sakagami, Daisuke Osaki, Tadashi Kitahara, and Yasuhiko Saito. 2022. "Behavioral and Immunohistochemical Evidence for Suppressive Effects of Goshajinkigan on Salicylate-Induced Tinnitus in Rats" Brain Sciences 12, no. 5: 587. https://doi.org/10.3390/brainsci12050587
APA StyleKitano, K., Yamashita, A., Sugimura, T., Okayasu, T., Sakagami, M., Osaki, D., Kitahara, T., & Saito, Y. (2022). Behavioral and Immunohistochemical Evidence for Suppressive Effects of Goshajinkigan on Salicylate-Induced Tinnitus in Rats. Brain Sciences, 12(5), 587. https://doi.org/10.3390/brainsci12050587