
Focused Ultrasound Stimulation as a Neuromodulatory Tool for Parkinson’s Disease: A Scoping Review

 ,
,  , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Study Selection and Reliability
2.3. Data Extraction
2.4. Outcomes
2.5. Synthesis of Results
3. Results
3.1. Characteristics of Inlcuded Studies
3.2. Outcome Measures
3.3. Trends and Findings from In Vitro Preclinical Studies
3.4. Trends and Findings from In Vivo Preclinical Studies
4. Discussion
4.1. Summary of Findings
4.2. Implications of Findings: Association and Causality
4.3. Implication on the Direction of Future Research
4.4. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vila, M.; Jackson-Lewis, V.; Vukosavic, S.; Djaldetti, R.; Liberatore, G.; Offen, D.; Korsmeyer, S.J.; Przedborski, S. Bax ablation prevents dopaminergic neurodegeneration in the 1-methyl- 4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2001, 98, 2837–2842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetz, C.G.; Tilley, B.C.; Shaftman, S.R.; Stebbins, G.T.; Fahn, S.; Martinez-Martin, P.; Poewe, W.; Sampaio, C.; Stern, M.B.; Dodel, R.; et al. Movement Disorder Society-sponsored revision of the Unified Parkinson’s Disease Rating Scale (MDS-UPDRS): Scale presentation and clinimetric testing results. Mov. Disord. 2008, 23, 2129–2170. [Google Scholar] [CrossRef] [PubMed]
- Kadastik-Eerme, L.; Rosenthal, M.; Paju, T.; Muldmaa, M.; Taba, P. Health-related quality of life in Parkinson’s disease: A cross-sectional study focusing on non-motor symptoms. Health Qual. Life Outcomes 2015, 13, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carod-Artal, F.J.; Vargas, A.P.; Martinez-Martin, P. Determinants of quality of life in Brazilian patients with Parkinson’s disease. Mov. Disord. 2007, 22, 1408–1415. [Google Scholar] [CrossRef]
- Hindle, J.V. Ageing, neurodegeneration and Parkinson’s disease. Age Ageing 2010, 39, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, L.; Jette, N.; Frolkis, A.; Steeves, T.; Pringsheim, T. The Incidence of Parkinson’s Disease: A Systematic Review and Meta-Analysis. Neuroepidemiology 2016, 46, 292–300. [Google Scholar] [CrossRef]
- Pringsheim, T.; Jette, N.; Frolkis, A.; Steeves, T.D. The prevalence of Parkinson’s disease: A systematic review and meta-analysis. Mov. Disord. 2014, 29, 1583–1590. [Google Scholar] [CrossRef]
- Yang, W.; Hamilton, J.L.; Kopil, C.; Beck, J.C.; Tanner, C.M.; Albin, R.L.; Ray Dorsey, E.; Dahodwala, N.; Cintina, I.; Hogan, P.; et al. Current and projected future economic burden of Parkinson’s disease in the U.S. NPJ Parkinson’s Dis. 2020, 6, 15. [Google Scholar] [CrossRef]
- Lozano, A.M.; Lipsman, N. Probing and regulating dysfunctional circuits using deep brain stimulation. Neuron 2013, 77, 406–424. [Google Scholar] [CrossRef] [Green Version]
- Lozano, A.M.; Lipsman, N.; Bergman, H.; Brown, P.; Chabardes, S.; Chang, J.W.; Matthews, K.; McIntyre, C.C.; Schlaepfer, T.E.; Schulder, M.; et al. Deep brain stimulation: Current challenges and future directions. Nat. Rev. Neurol. 2019, 15, 148–160. [Google Scholar] [CrossRef]
- McIntyre, C.C.; Anderson, R.W. Deep brain stimulation mechanisms: The control of network activity via neurochemistry modulation. J. Neurochem. 2016, 139 (Suppl. S1), 338–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.L.; Shao, B.; Chen, J.; Zhou, Y.; Lin, S.Y.; Wang, W.W. Effects of neurostimulation for advanced Parkinson’s disease patients on motor symptoms: A multiple-treatments meta-analysas of randomized controlled trials. Sci. Rep. 2016, 6, 25285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St George, R.J.; Nutt, J.G.; Burchiel, K.J.; Horak, F.B. A meta-regression of the long-term effects of deep brain stimulation on balance and gait in PD. Neurology 2010, 75, 1292–1299. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.; Gill, S.; Varma, T.; Jenkinson, C.; Quinn, N.; Mitchell, R.; Scott, R.; Ives, N.; Rick, C.; Daniels, J.; et al. Deep brain stimulation plus best medical therapy versus best medical therapy alone for advanced Parkinson’s disease (PD SURG trial): A randomised, open-label trial. Lancet Neurol. 2010, 9, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Weaver, F.M.; Follett, K.; Stern, M.; Hur, K.; Harris, C.; Marks, W.J.; Rothlind, J.; Sagher, O.; Reda, D.; Moy, C.S.; et al. Bilateral deep brain stimulation vs best medical therapy for patients with advanced Parkinson disease: A randomized controlled trial. JAMA 2009, 301, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zrinzo, L.; Foltynie, T.; Limousin, P.; Hariz, M.I. Reducing hemorrhagic complications in functional neurosurgery: A large case series and systematic literature review. J. Neurosurg. 2012, 116, 84–94. [Google Scholar] [CrossRef]
- Xiaowu, H.; Xiufeng, J.; Xiaoping, Z.; Bin, H.; Laixing, W.; Yiqun, C.; Jinchuan, L.; Aiguo, J.; Jianmin, L. Risks of intracranial hemorrhage in patients with Parkinson’s disease receiving deep brain stimulation and ablation. Parkinsonism Relat. Disord. 2010, 16, 96–100. [Google Scholar] [CrossRef]
- Jitkritsadakul, O.; Bhidayasiri, R.; Kalia, S.K.; Hodaie, M.; Lozano, A.M.; Fasano, A. Systematic review of hardware-related complications of Deep Brain Stimulation: Do new indications pose an increased risk? Brain Stimul. 2017, 10, 967–976. [Google Scholar] [CrossRef]
- Friehs, G.M.; Park, M.C.; Goldman, M.A.; Zerris, V.A.; Norén, G.; Sampath, P. Stereotactic radiosurgery for functional disorders. Neurosurg. Focus 2007, 23, E3. [Google Scholar] [CrossRef]
- Eisenberg, H.M.; Krishna, V.; Elias, W.J.; Cosgrove, G.R.; Gandhi, D.; Aldrich, C.E.; Fishman, P.S. MR-guided focused ultrasound pallidotomy for Parkinson’s disease: Safety and feasibility. J. Neurosurg. 2020, 135, 792–798. [Google Scholar] [CrossRef]
- Martínez-Fernández, R.; Máñez-Miró, J.U.; Rodríguez-Rojas, R.; Del Álamo, M.; Shah, B.B.; Hernández-Fernández, F.; Pineda-Pardo, J.A.; Monje, M.H.G.; Fernández-Rodríguez, B.; Sperling, S.A.; et al. Randomized Trial of Focused Ultrasound Subthalamotomy for Parkinson’s Disease. N. Engl. J. Med. 2020, 383, 2501–2513. [Google Scholar] [CrossRef] [PubMed]
- Jung, N.Y.; Park, C.K.; Kim, M.; Lee, P.H.; Sohn, Y.H.; Chang, J.W. The efficacy and limits of magnetic resonance-guided focused ultrasound pallidotomy for Parkinson’s disease: A Phase I clinical trial. J. Neurosurg. 2018, 130, 1853–1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ClinicalTrials.gov. Safety and Initial Effectiveness of Transcranial MR Guided Focused Ultrasound for the Treatment of Parkinson’s Disease (TDPD). Available online: https://clinicaltrials.gov/ct2/show/NCT04002596 (accessed on 31 May 2021).
- ClinicalTrials.gov. Parkinson’s Disease (PD) Treated with Focused Ultrasound Subthalamotomy at an Early Stage (EarlyFocus). Available online: https://clinicaltrials.gov/ct2/show/NCT04692116 (accessed on 31 May 2021).
- Cho, S.S.; Strafella, A.P. rTMS of the left dorsolateral prefrontal cortex modulates dopamine release in the ipsilateral anterior cingulate cortex and orbitofrontal cortex. PLoS ONE 2009, 4, e6725. [Google Scholar] [CrossRef] [Green Version]
- Brys, M.; Fox, M.D.; Agarwal, S.; Biagioni, M.; Dacpano, G.; Kumar, P.; Pirraglia, E.; Chen, R.; Wu, A.; Fernandez, H.; et al. Multifocal repetitive TMS for motor and mood symptoms of Parkinson disease: A randomized trial. Neurology 2016, 87, 1907–1915. [Google Scholar] [CrossRef] [Green Version]
- Ba, M.; Kong, M.; Guan, L.; Yi, M.; Zhang, H. Repetitive transcranial magnetic stimulation (rTMS) improves behavioral and biochemical deficits in levodopa-induced dyskinetic rats model. Oncotarget 2016, 7, 58802–58812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefaucheur, J.P.; Antal, A.; Ayache, S.S.; Benninger, D.H.; Brunelin, J.; Cogiamanian, F.; Cotelli, M.; De Ridder, D.; Ferrucci, R.; Langguth, B.; et al. Evidence-based guidelines on the therapeutic use of transcranial direct current stimulation (tDCS). Clin. Neurophysiol. 2017, 128, 56–92. [Google Scholar] [CrossRef]
- Fini, M.; Tyler, W.J. Transcranial focused ultrasound: A new tool for non-invasive neuromodulation. Int. Rev. Psychiatry 2017, 29, 168–177. [Google Scholar] [CrossRef]
- Goetz, S.M.; Deng, Z.D. The development and modelling of devices and paradigms for transcranial magnetic stimulation. Int. Rev. Psychiatry 2017, 29, 115–145. [Google Scholar] [CrossRef]
- Bonmassar, G.; Lee, S.W.; Freeman, D.K.; Polasek, M.; Fried, S.I.; Gale, J.T. Microscopic magnetic stimulation of neural tissue. Nat. Commun. 2012, 3, 921. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, W.D. Ultrasound-biophysics mechanisms. Prog. Biophys. Mol. Biol. 2007, 93, 212–255. [Google Scholar] [CrossRef] [Green Version]
- Mueller, J.; Legon, W.; Opitz, A.; Sato, T.F.; Tyler, W.J. Transcranial focused ultrasound modulates intrinsic and evoked EEG dynamics. Brain Stimul. 2014, 7, 900–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legon, W.; Sato, T.F.; Opitz, A.; Mueller, J.; Barbour, A.; Williams, A.; Tyler, W.J. Transcranial focused ultrasound modulates the activity of primary somatosensory cortex in humans. Nat. Neurosci. 2014, 17, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Munn, Z.; Peters, M.D.J.; Stern, C.; Tufanaru, C.; McArthur, A.; Aromataris, E. Systematic review or scoping review? Guidance for authors when choosing between a systematic or scoping review approach. BMC Med. Res. Methodol. 2018, 18, 143. [Google Scholar] [CrossRef] [PubMed]
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L.; et al. PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and Explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.S.; Zhang, J.J.Y.; Nga, V.D.W.; Ng, C.H.; Tai, B.C.; Higgins, J.; Syn, N. Tenets for the Proper Conduct and Use of Meta-Analyses: A Practical Guide for Neurosurgeons. World Neurosurg. 2022, in press. [Google Scholar]
- Lee, K.S.; Zhang, J.J.Y.; Alamri, A.; Chari, A. Neurosurgery Education in the Medical School Curriculum: A Scoping Review. World Neurosurg. 2020, 144, e631–e642. [Google Scholar] [CrossRef]
- Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 1960, 20, 37–47. [Google Scholar] [CrossRef]
- Chen, X.; Wang, D.; Zhang, L.; Yao, H.; Zhu, H.; Zhao, N.; Peng, X.; Yang, K. Neuroprotective Effect of Low-Intensity Pulsed Ultrasound on the Mouse MPTP/MPP. Ultrasound Med. Biol. 2021, 47, 2321–2330. [Google Scholar] [CrossRef]
- Dong, Y.; Liu, D.; Zhao, Y.; Yuan, Y.; Wang, W.; Wu, S.; Liang, X.; Wang, Z.; Liu, L. Assessment of Neuroprotective Effects of Low-Intensity Transcranial Ultrasound Stimulation in a Parkinson’s Disease Rat Model by Fractional Anisotropy and Relaxation Time T2. Front. Neurosci. 2021, 15, 590354. [Google Scholar] [CrossRef] [PubMed]
- Karmacharya, M.B.; Hada, B.; Park, S.R.; Choi, B.H. Low-Intensity Ultrasound Decreases α-Synuclein Aggregation via Attenuation of Mitochondrial Reactive Oxygen Species in MPP(+)-Treated PC12 Cells. Mol. Neurobiol. 2017, 54, 6235–6244. [Google Scholar] [CrossRef]
- Sung, C.Y.; Chiang, P.K.; Tsai, C.W.; Yang, F.Y. Low-Intensity Pulsed Ultrasound Enhances Neurotrophic Factors and Alleviates Neuroinflammation in a Rat Model of Parkinson’s Disease. Cereb. Cortex 2021. [Google Scholar] [CrossRef] [PubMed]
- Tarnaud, T.; Joseph, W.; Martens, L.; Tanghe, E. Computational Modeling of Ultrasonic Subthalamic Nucleus Stimulation. IEEE Trans. Biomed. Eng. 2019, 66, 1155–1164. [Google Scholar] [CrossRef]
- Wang, Z.; Yan, J.; Wang, X.; Yuan, Y.; Li, X. Transcranial Ultrasound Stimulation Directly Influences the Cortical Excitability of the Motor Cortex in Parkinsonian Mice. Mov. Disord. 2020, 35, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Lu, X.; Peng, D.; Wang, G.; Chen, C.; Liu, W.; Wu, W.; Mason, T.J. Ultrasonic stimulation of the brain to enhance the release of dopamine—A potential novel treatment for Parkinson’s disease. Ultrason. Sonochem. 2020, 63, 104955. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhao, Z.; Wang, Z.; Wang, X.; Yan, J.; Li, X. The Effect of Low-Intensity Transcranial Ultrasound Stimulation on Behavior in a Mouse Model of Parkinson’s Disease Induced by MPTP. IEEE Trans. Neural Syst. Rehabil. Eng. 2020, 28, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Feng, Y.; Shi, A.; Zhang, L.; Guo, S.; Wan, M. Neuroprotective Effect of Low-Intensity Pulsed Ultrasound against MPP. Ultrasound Med. Biol. 2017, 43, 1986–1999. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Niu, L.; Meng, L.; Lin, Z.; Zou, J.; Xia, X.; Huang, X.; Zhou, W.; Bian, T.; Zheng, H. Noninvasive Ultrasound Deep Brain Stimulation for the Treatment of Parkinson’s Disease Model Mouse. Research 2019, 2019, 1748489. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Niu, L.; Xia, X.; Lin, Z.; Liu, X.; Su, M.; Guo, R.; Meng, L.; Zheng, H. Wearable Ultrasound Improves Motor Function in an MPTP Mouse Model of Parkinson’s Disease. IEEE Trans. Biomed. Eng. 2019, 66, 3006–3013. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.; McDonnell, J.M.; Korsmeyer, S.J. BCL-2 family members and the mitochondria in apoptosis. Genes Dev. 1999, 13, 1899–1911. [Google Scholar] [CrossRef] [Green Version]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Sas, K.; Robotka, H.; Toldi, J.; Vécsei, L. Mitochondria, metabolic disturbances, oxidative stress and the kynurenine system, with focus on neurodegenerative disorders. J. Neurol. Sci. 2007, 257, 221–239. [Google Scholar] [CrossRef] [PubMed]
- Rekha, K.R.; Selvakumar, G.P. Gene expression regulation of Bcl2, Bax and cytochrome-C by geraniol on chronic MPTP/probenecid induced C57BL/6 mice model of Parkinson’s disease. Chem. Biol. Interact. 2014, 217, 57–66. [Google Scholar] [CrossRef]
- Robinson, E.J.; Aguiar, S.P.; Kouwenhoven, W.M.; Starmans, D.S.; von Oerthel, L.; Smidt, M.P.; van der Heide, L.P. Survival of midbrain dopamine neurons depends on the Bcl2 factor Mcl1. Cell Death Discov. 2018, 4, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kluck, R.M.; Bossy-Wetzel, E.; Green, D.R.; Newmeyer, D.D. The release of cytochrome c from mitochondria: A primary site for Bcl-2 regulation of apoptosis. Science 1997, 275, 1132–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Caldas, M.; Carvalho, A.N.; Rodrigues, E.; Henderson, C.J.; Wolf, C.R.; Rodrigues, C.M.; Gama, M.J. Tauroursodeoxycholic acid prevents MPTP-induced dopaminergic cell death in a mouse model of Parkinson’s disease. Mol. Neurobiol. 2012, 46, 475–486. [Google Scholar] [CrossRef]
- Choi, S.; Oh, J.Y.; Kim, S.J. Ginsenoside Rh2 induces Bcl-2 family proteins-mediated apoptosis in vitro and in xenografts in vivo models. J. Cell Biochem. 2011, 112, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Offen, D.; Beart, P.M.; Cheung, N.S.; Pascoe, C.J.; Hochman, A.; Gorodin, S.; Melamed, E.; Bernard, R.; Bernard, O. Transgenic mice expressing human Bcl-2 in their neurons are resistant to 6-hydroxydopamine and 1-methyl-4-phenyl-1,2,3,6- tetrahydropyridine neurotoxicity. Proc. Natl. Acad. Sci. USA 1998, 95, 5789–5794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Matthews, R.T.; Schulz, J.B.; Klockgether, T.; Liao, A.W.; Martinou, J.C.; Penney, J.B.; Hyman, B.T.; Beal, M.F. 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyride neurotoxicity is attenuated in mice overexpressing Bcl-2. J. Neurosci. 1998, 18, 8145–8152. [Google Scholar] [CrossRef]
- Okochi, M.; Walter, J.; Koyama, A.; Nakajo, S.; Baba, M.; Iwatsubo, T.; Meijer, L.; Kahle, P.J.; Haass, C. Constitutive phosphorylation of the Parkinson’s disease associated alpha-synuclein. J. Biol. Chem. 2000, 275, 390–397. [Google Scholar] [CrossRef] [Green Version]
- Simón-Sánchez, J.; Schulte, C.; Bras, J.M.; Sharma, M.; Gibbs, J.R.; Berg, D.; Paisan-Ruiz, C.; Lichtner, P.; Scholz, S.W.; Hernandez, D.G.; et al. Genome-wide association study reveals genetic risk underlying Parkinson’s disease. Nat. Genet. 2009, 41, 1308–1312. [Google Scholar] [CrossRef]
- Devine, M.J.; Ryten, M.; Vodicka, P.; Thomson, A.J.; Burdon, T.; Houlden, H.; Cavaleri, F.; Nagano, M.; Drummond, N.J.; Taanman, J.W.; et al. Parkinson’s disease induced pluripotent stem cells with triplication of the α-synuclein locus. Nat. Commun. 2011, 2, 440. [Google Scholar] [CrossRef]
- Lin, L.F.; Doherty, D.H.; Lile, J.D.; Bektesh, S.; Collins, F. GDNF: A glial cell line-derived neurotrophic factor for midbrain dopaminergic neurons. Science 1993, 260, 1130–1132. [Google Scholar] [CrossRef]
- Fan, C.H.; Ting, C.Y.; Lin, C.Y.; Chan, H.L.; Chang, Y.C.; Chen, Y.Y.; Liu, H.L.; Yeh, C.K. Noninvasive, Targeted, and Non-Viral Ultrasound-Mediated GDNF-Plasmid Delivery for Treatment of Parkinson’s Disease. Sci. Rep. 2016, 6, 19579. [Google Scholar] [CrossRef] [PubMed]
- Pouliopoulos, A.N.; Kwon, N.; Jensen, G.; Meaney, A.; Niimi, Y.; Burgess, M.T.; Ji, R.; McLuckie, A.J.; Munoz, F.A.; Kamimura, H.A.S.; et al. Safety evaluation of a clinical focused ultrasound system for neuronavigation guided blood-brain barrier opening in non-human primates. Sci. Rep. 2021, 11, 15043. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.T.; Wei, K.C.; Liu, H.L. Theranostic Strategy of Focused Ultrasound Induced Blood-Brain Barrier Opening for CNS Disease Treatment. Front. Pharmacol. 2019, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Carpentier, A.; Canney, M.; Vignot, A.; Reina, V.; Beccaria, K.; Horodyckid, C.; Karachi, C.; Leclercq, D.; Lafon, C.; Chapelon, J.Y.; et al. Clinical trial of blood-brain barrier disruption by pulsed ultrasound. Sci. Transl. Med. 2016, 8, 343re342. [Google Scholar] [CrossRef]
- Fedak, K.M.; Bernal, A.; Capshaw, Z.A.; Gross, S. Applying the Bradford Hill criteria in the 21st century: How data integration has changed causal inference in molecular epidemiology. Emerg. Themes. Epidemiol. 2015, 12, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyler, W.J.; Tufail, Y.; Finsterwald, M.; Tauchmann, M.L.; Olson, E.J.; Majestic, C. Remote excitation of neuronal circuits using low-intensity, low-frequency ultrasound. PLoS ONE 2008, 3, e3511. [Google Scholar] [CrossRef]
- Tufail, Y.; Yoshihiro, A.; Pati, S.; Li, M.M.; Tyler, W.J. Ultrasonic neuromodulation by brain stimulation with transcranial ultrasound. Nat. Protoc. 2011, 6, 1453–1470. [Google Scholar] [CrossRef]
- Kubanek, J.; Shukla, P.; Das, A.; Baccus, S.A.; Goodman, M.B. Ultrasound Elicits Behavioral Responses through Mechanical Effects on Neurons and Ion Channels in a Simple Nervous System. J. Neurosci. 2018, 38, 3081–3091. [Google Scholar] [CrossRef] [Green Version]
- Tufail, Y.; Matyushov, A.; Baldwin, N.; Tauchmann, M.L.; Georges, J.; Yoshihiro, A.; Tillery, S.I.; Tyler, W.J. Transcranial pulsed ultrasound stimulates intact brain circuits. Neuron 2010, 66, 681–694. [Google Scholar] [CrossRef] [Green Version]
- Kamimura, H.A.; Wang, S.; Chen, H.; Wang, Q.; Aurup, C.; Acosta, C.; Carneiro, A.A.; Konofagou, E.E. Focused ultrasound neuromodulation of cortical and subcortical brain structures using 1.9 MHz. Med. Phys. 2016, 43, 5730. [Google Scholar] [CrossRef]
- Pouget, P.; Frey, S.; Ahnine, H.; Attali, D.; Claron, J.; Constans, C.; Aubry, J.F.; Arcizet, F. Neuronavigated Repetitive Transcranial Ultrasound Stimulation Induces Long-Lasting and Reversible Effects on Oculomotor Performance in Non-human Primates. Front. Physiol. 2020, 11, 1042. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.; Mittelstein, D.R.; Hurt, R.; Lacroix, J.; Shapiro, M.G. Focused ultrasound excites neurons via mechanosensitive calcium accumulation and ion channel amplification. Nat Commun 2022, 13, 493. [Google Scholar] [CrossRef] [PubMed]
- Krasovitski, B.; Frenkel, V.; Shoham, S.; Kimmel, E. Intramembrane cavitation as a unifying mechanism for ultrasound-induced bioeffects. Proc. Natl. Acad. Sci. USA 2011, 108, 3258–3263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.S.; Bystritsky, A.; Lee, J.H.; Zhang, Y.; Fischer, K.; Min, B.K.; McDannold, N.J.; Pascual-Leone, A.; Jolesz, F.A. Focused ultrasound modulates region-specific brain activity. Neuroimage 2011, 56, 1267–1275. [Google Scholar] [CrossRef] [Green Version]
- Kubanek, J. Neuromodulation with transcranial focused ultrasound. Neurosurg. Focus 2018, 44, E14. [Google Scholar] [CrossRef] [Green Version]
- Jerusalem, A.; Al-Rekabi, Z.; Chen, H.; Ercole, A.; Malboubi, M.; Tamayo-Elizalde, M.; Verhagen, L.; Contera, S. Electrophysiological-mechanical coupling in the neuronal membrane and its role in ultrasound neuromodulation and general anaesthesia. Acta Biomater. 2019, 97, 116–140. [Google Scholar] [CrossRef]
- Prieto, M.L.; Ömer, O.; Khuri-Yakub, B.T.; Maduke, M.C. Dynamic response of model lipid membranes to ultrasonic radiation force. PLoS ONE 2013, 8, e77115. [Google Scholar] [CrossRef]
- Renzhiglova, E.; Ivantsiv, V.; Xu, Y. Difference frequency magneto-acousto-electrical tomography (DF-MAET): Application of ultrasound-induced radiation force to imaging electrical current density. IEEE Trans. Ultrason. Ferroelectr. Freq. Control 2010, 57, 2391–2402. [Google Scholar] [CrossRef]
- Kubanek, J.; Shi, J.; Marsh, J.; Chen, D.; Deng, C.; Cui, J. Ultrasound modulates ion channel currents. Sci. Rep. 2016, 6, 24170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto, M.L.; Firouzi, K.; Khuri-Yakub, B.T.; Maduke, M. Activation of Piezo1 but Not Na. Ultrasound Med. Biol. 2018, 44, 1217–1232. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Shi, Y.; Lu, L.; Liu, L.; Cai, Y.; Zheng, H.; Liu, X.; Yan, F.; Zou, C.; Sun, C.; et al. Targeted delivery of GDNF through the blood-brain barrier by MRI-guided focused ultrasound. PLoS ONE 2012, 7, e52925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Deng, J.; Wang, F.; Chen, S.; Liu, Y.; Wang, Z.; Cheng, Y. Targeted gene delivery to the mouse brain by MRI-guided focused ultrasound-induced blood-brain barrier disruption. Exp. Neurol. 2012, 233, 350–356. [Google Scholar] [CrossRef]
- Allen, N.J.; Barres, B.A. Neuroscience: Glia—More than just brain glue. Nature 2009, 457, 675–677. [Google Scholar] [CrossRef]
- De Luca, C.; Colangelo, A.M.; Virtuoso, A.; Alberghina, L.; Papa, M. Neurons, Glia, Extracellular Matrix and Neurovascular Unit: A Systems Biology Approach to the Complexity of Synaptic Plasticity in Health and Disease. Int. J. Mol. Sci. 2020, 21, 1539. [Google Scholar] [CrossRef] [Green Version]
- Grosche, J.; Matyash, V.; Möller, T.; Verkhratsky, A.; Reichenbach, A.; Kettenmann, H. Microdomains for neuron-glia interaction: Parallel fiber signaling to Bergmann glial cells. Nat. Neurosci. 1999, 2, 139–143. [Google Scholar] [CrossRef]
- Tremblay, M.E.; Cookson, M.R.; Civiero, L. Glial phagocytic clearance in Parkinson’s disease. Mol. Neurodegener. 2019, 14, 16. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.J.; Lee, J.M.; Kim, H.B.; Lee, J.; Han, S.; Bae, J.Y.; Hong, G.S.; Koh, W.; Kwon, J.; Hwang, E.S.; et al. Ultrasonic Neuromodulation via Astrocytic TRPA1. Curr. Biol. 2020, 30, 948. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.J.; Mannaioni, G.; Yuan, H.; Woo, D.H.; Gingrich, M.B.; Traynelis, S.F. Astrocytic control of synaptic NMDA receptors. J. Physiol. 2007, 581, 1057–1081. [Google Scholar] [CrossRef]
- Clasadonte, J.; Dong, J.; Hines, D.J.; Haydon, P.G. Astrocyte control of synaptic NMDA receptors contributes to the progressive development of temporal lobe epilepsy. Proc. Natl. Acad. Sci. USA 2013, 110, 17540–17545. [Google Scholar] [CrossRef] [Green Version]
- Blackmore, D.G.; Turpin, F.; Palliyaguru, T.; Evans, H.T.; Chicoteau, A.; Lee, W.; Pelekanos, M.; Nguyen, N.; Song, J.; Sullivan, R.K.P.; et al. Low-intensity ultrasound restores long-term potentiation and memory in senescent mice through pleiotropic mechanisms including NMDAR signaling. Mol. Psychiatry 2021. [Google Scholar] [CrossRef]
- Lee, K.S.; Young, A.; King, H.; Jenkins, A.T.A.; Davies, A. Variation in Definitions of Burn Wound Infection Limits the Validity of Systematic Review Findings in Burn Care: A Systematic Review of Reviews. Burns, 2021; in press. [Google Scholar] [CrossRef] [PubMed]
- Björklund, A.; Cenci-Nilsson, A. Recent Advances in Parkinson’s Disease; Elsevier: Amsterdam, The Netherlands, 2020; Volume 252. [Google Scholar]
- Folloni, D.; Verhagen, L.; Mars, R.B.; Fouragnan, E.; Constans, C.; Aubry, J.F.; Rushworth, M.F.S.; Sallet, J. Manipulation of Subcortical and Deep Cortical Activity in the Primate Brain Using Transcranial Focused Ultrasound Stimulation. Neuron 2019, 101, 1109–1116. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, M.; Shimizu, K.; Furuya, H.; Sakamoto, T.; Ohnishi, H.; Karasawa, J. Effect of isoflurane on motor-evoked potentials induced by direct electrical stimulation of the exposed motor cortex with single, double, and triple stimuli in rats. Anesthesiology 1996, 85, 1176–1183. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Kim, M.; Kim, H.; Shin, H.; Youn, I. Ketamine Inhibits Ultrasound Stimulation-Induced Neuromodulation by Blocking Cortical Neuron Activity. Ultrasound Med. Biol. 2018, 44, 635–646. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soldner, F.; Hockemeyer, D.; Beard, C.; Gao, Q.; Bell, G.W.; Cook, E.G.; Hargus, G.; Blak, A.; Cooper, O.; Mitalipova, M.; et al. Parkinson’s disease patient-derived induced pluripotent stem cells free of viral reprogramming factors. Cell 2009, 136, 964–977. [Google Scholar] [CrossRef] [Green Version]
- Clennell, B.; Steward, T.G.J.; Elley, M.; Shin, E.; Weston, M.; Drinkwater, B.W.; Whitcomb, D.J. Transient ultrasound stimulation has lasting effects on neuronal excitability. Brain Stimul. 2021, 14, 217–225. [Google Scholar] [CrossRef]
- Legon, W.; Adams, S.; Bansal, P.; Patel, P.D.; Hobbs, L.; Ai, L.; Mueller, J.K.; Meekins, G.; Gillick, B.T. A retrospective qualitative report of symptoms and safety from transcranial focused ultrasound for neuromodulation in humans. Sci. Rep. 2020, 10, 5573. [Google Scholar] [CrossRef]
- Simonsohn, U.; Nelson, L.D.; Simmons, J.P. P-curve: A key to the file-drawer. J. Exp. Psychol. Gen. 2014, 143, 534–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
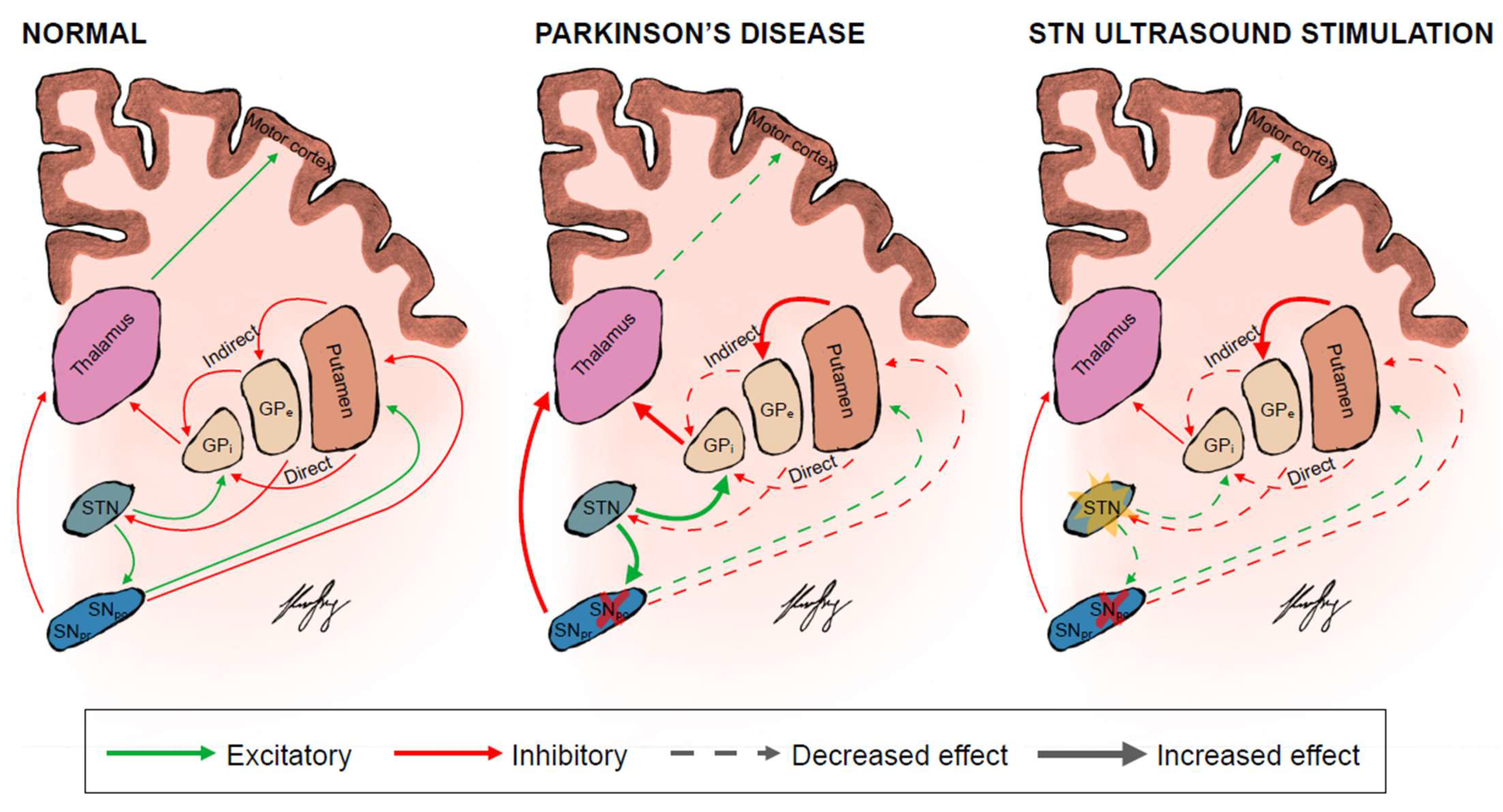


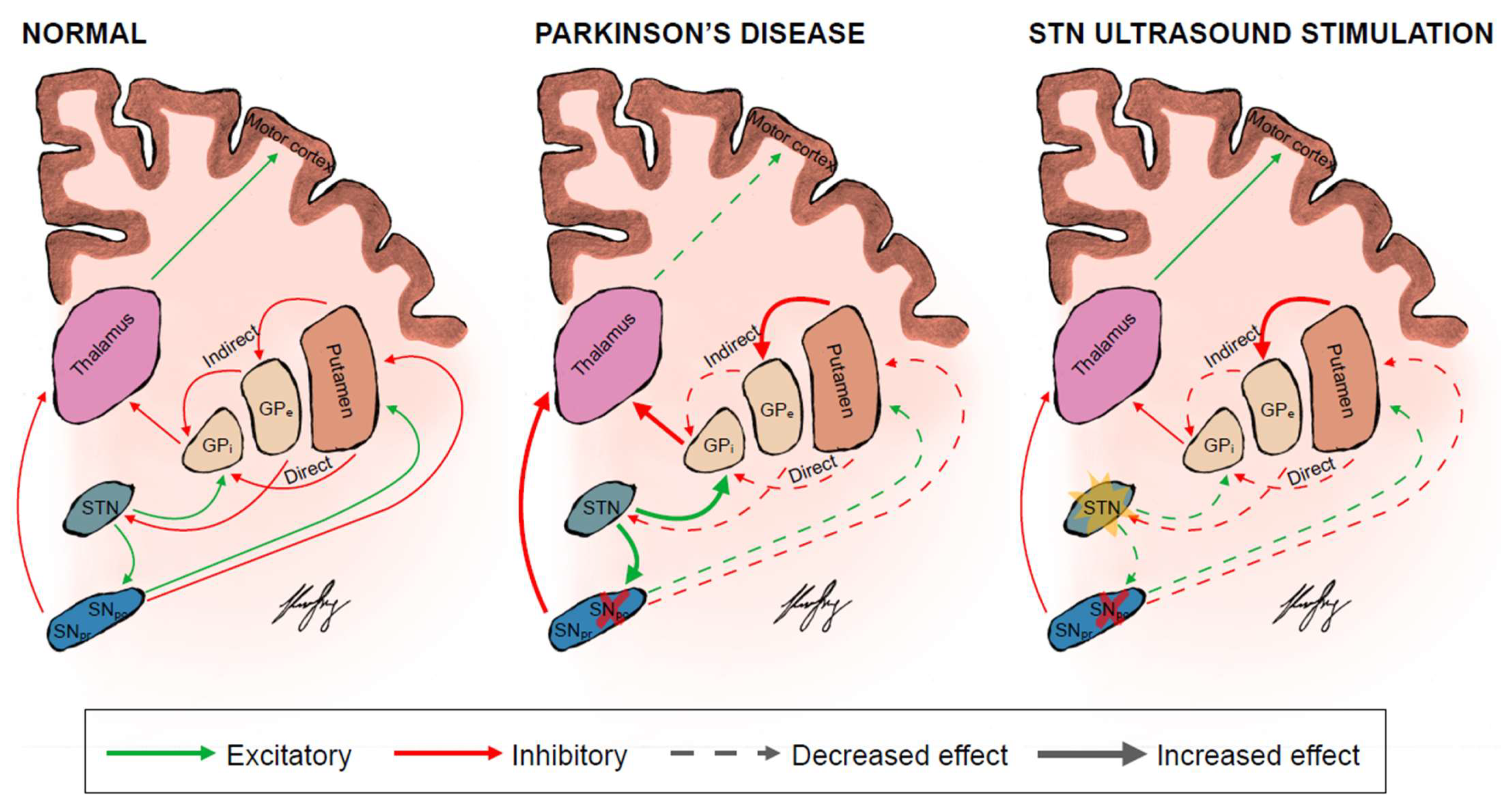
{kind=link}
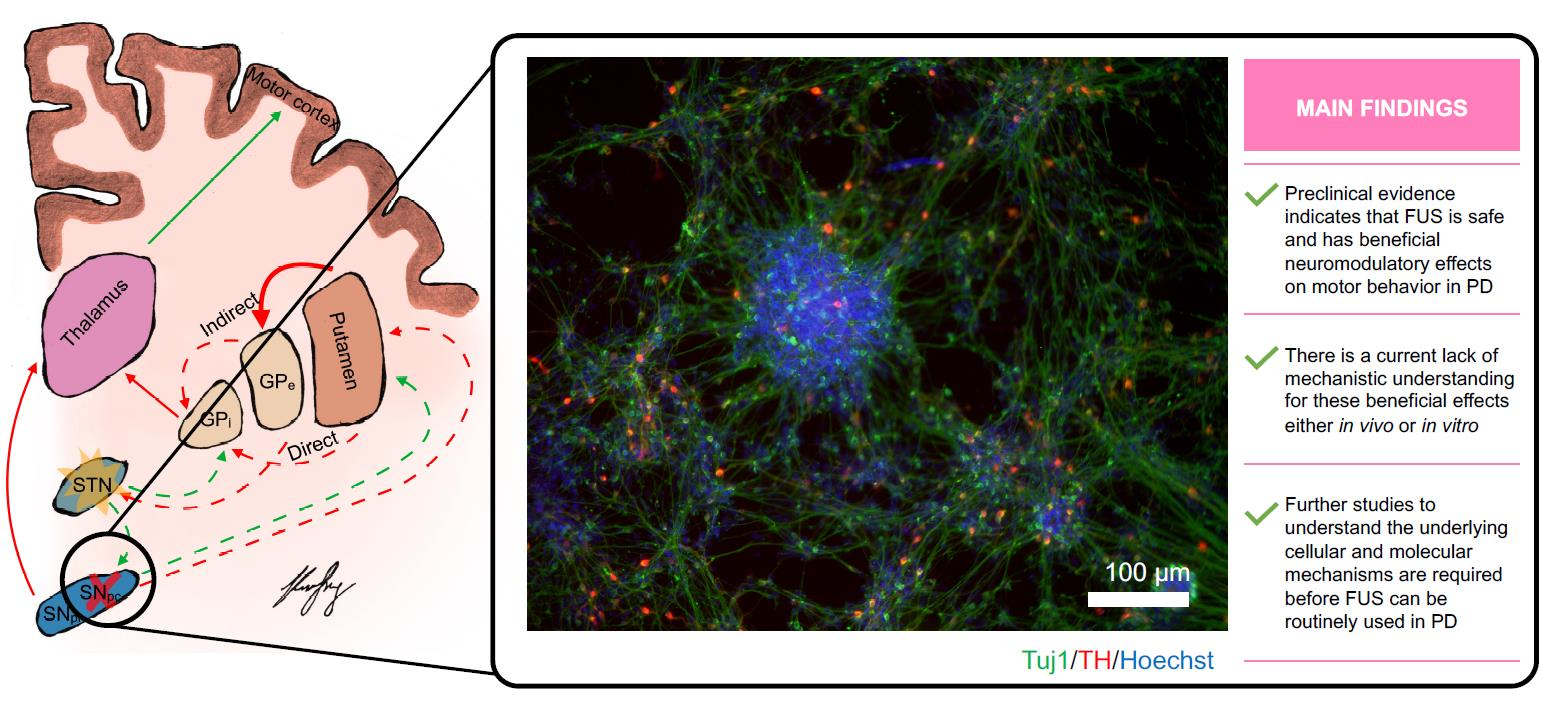
{kind=link}
{kind=link}
| Authors and Year | Country | Study Design | Sample Size, Types of Subjects/Participants | Intervention (Control) | Outcome Measures/Indicators | Main Findings Relating to FUS in PD |
|---|---|---|---|---|---|---|
| Chen X et al., 2021 | China | In vivo and in vitro | N = 5 per group. MPTP-induced C57BL/6 PD mice models. MPP+-induced N2a cells | LIFUS In vivo:
| In vivo measures:
| Efficacy:
|
| Dong Y et al., 2021 | China | In vivo | N = 20. 6-OHDA-induced Sprague-Dawley hemi-PD rat models | LIFUS
|
|
|
| Karmacharya MB et al., 2017 | Korea | In vitro | MPP+-induced PC12 cell PD models | Low-intensity ultrasound
|
|
|
| Sung CY. et al., 2021 | Taiwan | In vivo | N = 20. 6-OHDA-induced Sprague–Dawley PD rat models | LIFUS
|
|
|
| Tarnaud T et al., 2019 | Belgium | Computational modelling study | A computational model for ultrasonic stimulation of the STN is created by combining the Otsuka model with the bilayer sonophore model | LIFUS
|
|
|
| Wang Z et al., 2020 | China | In vivo | N = 11. MPTP-induced C57BL/6 PD mice models | LIFUS
|
|
|
| Xu T et al., 2020 | China | In vivo and in vitro | N = 12 per group. MPTP-induced C57BL/6 PD mice models and PC12 cells | LIFUS
| In vivo measures: Locomotor behavior
| Efficacy
|
| Yuan Y et al. 2020 | China | In vivo | N = 8 per group. MPTP-induced C57BL/6 PD mice models | LIFUS
|
|
|
| Zhao L et al., 2017 | China | In vitro | PC12 cells exposed to MPP+-induced neurotoxicity | Low-intensity ultrasound
|
|
|
| Zhou H et al., 2019a | China | In vivo | N = 8. MPTP induced C57BL/6 PD mice models | LIFUS
|
| Efficacy
|
| Zhou H et al., 2019b | China | In vivo | N = 8. MPTP-induced C57BL/6 PD mice models | LIFUS
|
| Efficacy
|
| Outcome Measures/Indicators |
|---|
| Preclinical—in vitro |
Changes in expression at the gene, RNA and protein level
|
Changes in mitochondrial integrity
|
| Preclinical—in vivo |
Changes in expression at the gene, RNA and protein level
|
Electrophysiological and synaptic properties
|
Behavioral outcomes
|
| Clinical |
Functional outcomes
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.S.; Clennell, B.; Steward, T.G.J.; Gialeli, A.; Cordero-Llana, O.; Whitcomb, D.J. Focused Ultrasound Stimulation as a Neuromodulatory Tool for Parkinson’s Disease: A Scoping Review. Brain Sci. 2022, 12, 289. https://doi.org/10.3390/brainsci12020289
Lee KS, Clennell B, Steward TGJ, Gialeli A, Cordero-Llana O, Whitcomb DJ. Focused Ultrasound Stimulation as a Neuromodulatory Tool for Parkinson’s Disease: A Scoping Review. Brain Sciences. 2022; 12(2):289. https://doi.org/10.3390/brainsci12020289
Chicago/Turabian StyleLee, Keng Siang, Benjamin Clennell, Tom G. J. Steward, Andriana Gialeli, Oscar Cordero-Llana, and Daniel J. Whitcomb. 2022. "Focused Ultrasound Stimulation as a Neuromodulatory Tool for Parkinson’s Disease: A Scoping Review" Brain Sciences 12, no. 2: 289. https://doi.org/10.3390/brainsci12020289
APA StyleLee, K. S., Clennell, B., Steward, T. G. J., Gialeli, A., Cordero-Llana, O., & Whitcomb, D. J. (2022). Focused Ultrasound Stimulation as a Neuromodulatory Tool for Parkinson’s Disease: A Scoping Review. Brain Sciences, 12(2), 289. https://doi.org/10.3390/brainsci12020289