
Functional Connectivity Signatures Underlying Simultaneous Language Translation in Interpreters and Non-Interpreters of Mandarin and English: An fNIRS Study



Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Materials
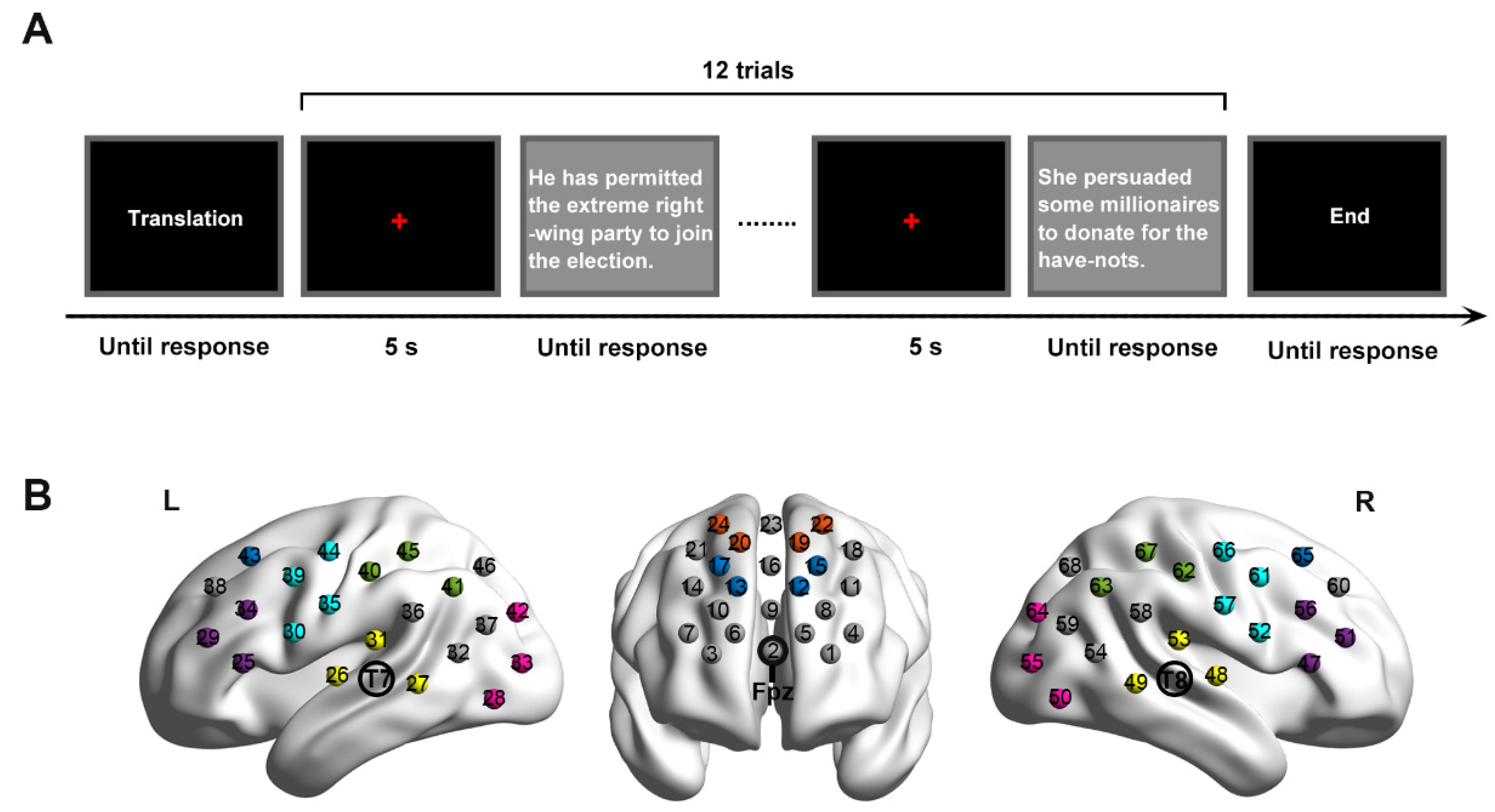
2.3. Procedure
2.4. fNIRS Data Acquisition
2.5. Data Analysis
2.5.1. Behavioral Performance
2.5.2. fNIRS Data Analysis
3. Results
3.1. Behavioral Performance
3.2. Functional Connectivity in IG
3.3. Functional Connectivity in NIG
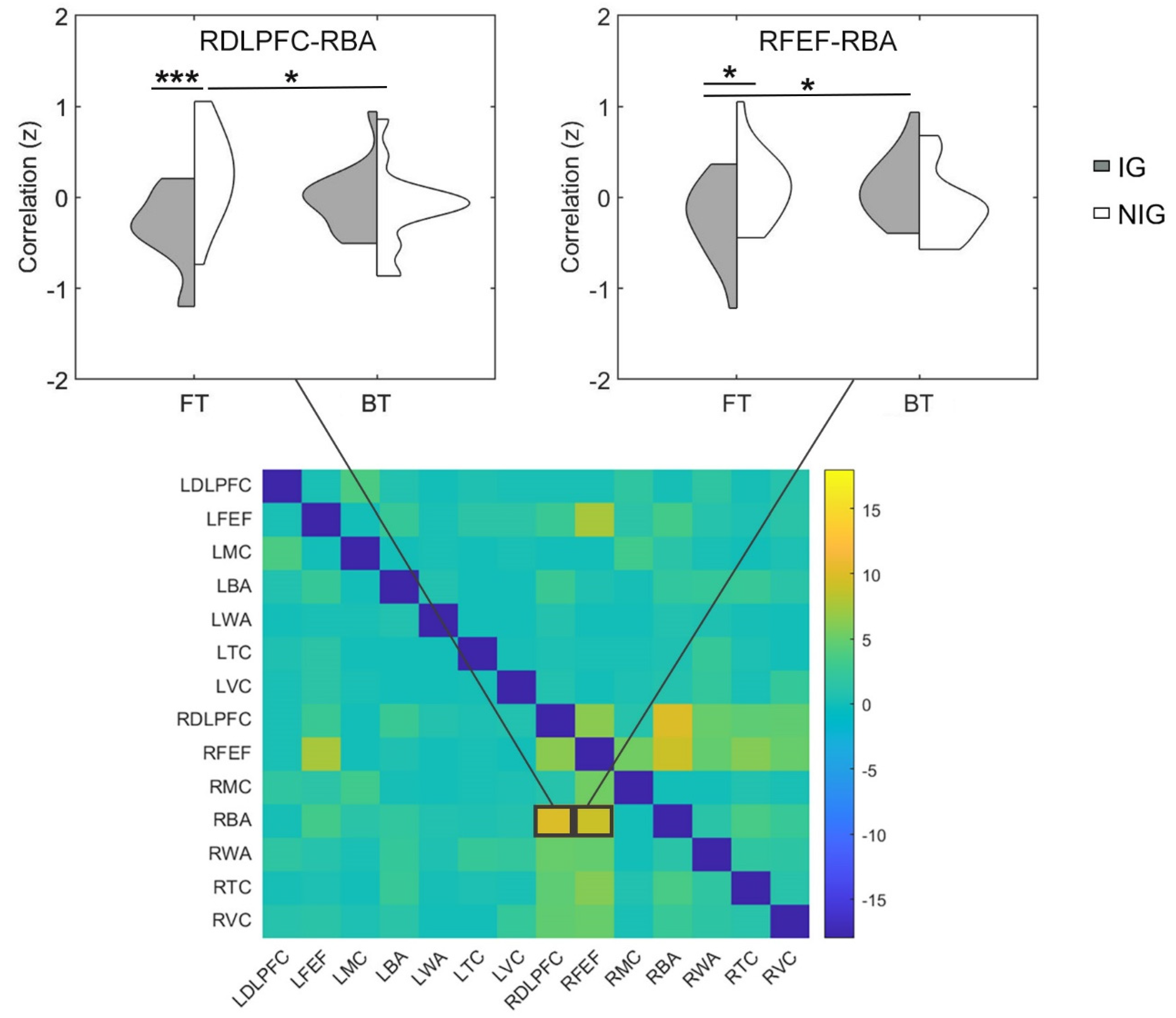
3.4. Functional Connectivity Difference between IG and NIG
3.5. Associations between Functional Connectivity and Sight Translation Performance
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bilalić, M.; Langner, R.; Ulrich, R.; Grodd, W. Many faces of expertise: Fusiform face area in chess experts and novices. J. Neurosci. 2011, 31, 10206–10214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo-Merino, B.; Glaser, D.E.; Grèzes, J.; Passingham, R.E.; Haggard, P. Action observation and acquired motor skills: An FMRI study with expert dancers. Cereb. Cortex 2005, 15, 1243–1249. [Google Scholar] [CrossRef] [Green Version]
- Jeon, H.A.; Friederici, A.D. What does “Being an Expert” mean to the brain? Functional specificity and connectivity in expertise. Cereb. Cortex 2017, 27, 5603–5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, N.; Lotze, M.; Eickhoff, S.B. Cognitive expertise: An ALE meta-analysis. Hum. Brain Mapp. 2016, 37, 262–272. [Google Scholar] [CrossRef]
- Becker, M.; Schubert, T.; Strobach, T.; Gallinat, J.; Kühn, S. Simultaneous interpreters vs. professional multilingual controls: Group differences in cognitive control as well as brain structure and function. Neuroimage 2016, 134, 250–260. [Google Scholar] [CrossRef]
- Elmer, S.; Kühnis, J. Functional connectivity in the left dorsal stream facilitates simultaneous language translation: An EEG study. Front. Hum. Neurosci. 2016, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Elmer, S.; Hänggi, J.; Jäncke, L. Processing demands upon cognitive, linguistic, and articulatory functions promote grey matter plasticity in the adult multilingual brain: Insights from simultaneous interpreters. Cortex 2014, 54, 179–189. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Hu, Y.; Yang, Y.; Li, D.; Hu, Y. Optical mapping of brain activity underlying directionality and its modulation by expertise in Mandarin/English interpreting. Front. Hum. Neurosci. 2021, 15, 649578. [Google Scholar] [CrossRef]
- Zheng, B.; Báez, S.; Su, L.; Xiang, X.; Weis, S.; Ibáñez, A.; García, A.M. Semantic and attentional networks in bilingual processing: fMRI connectivity signatures of translation directionality. Brain Cogn. 2020, 143, 105584. [Google Scholar] [CrossRef]
- Jost, L.B.; Radman, N.; Buetler, K.A.; Annoni, J.M. Behavioral and electrophysiological signatures of word translation processes. Neuropsychologia 2018, 109, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Van de Putte, E.; De Baene, W.; García-Pentón, L.; Woumans, E.; Dijkgraaf, A.; Duyck, W. Anatomical and functional changes in the brain after simultaneous interpreting training: A longitudinal study. Cortex 2018, 99, 243–257. [Google Scholar] [CrossRef] [PubMed]
- García, A.M.; Mikulan, E.; Ibáñez, A. A neuroscientific toolkit for translation studies. In Reembedding Translation Process Research; Muñoz Martín, R., Ed.; John Benjamins: Amsterdam, The Netherlands; Philadelphia, PA, USA, 2016; pp. 21–46. [Google Scholar]
- Hervais-Adelman, A.; Moser-Mercer, B.; Murray, M.M.; Golestani, N. Cortical thickness increases after simultaneous interpretation training. Neuropsychologia 2017, 98, 212–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hervais-Adelman, A.; Moser-Mercer, B.; Golestani, N. Brain functional plasticity associated with the emergence of expertise in extreme language control. NeuroImage 2015, 114, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Hervais-Adelman, A.; Moser-Mercer, B.; Golestani, N. Executive control of language in the bilingual brain: Integrating the evidence from neuroimaging to neuropsychology. Front. Psychol. 2011, 2, 234. [Google Scholar] [CrossRef] [Green Version]
- Quaresima, V.; Ferrari, M.; van der Sluijs, M.C.; Menssen, J.; Colier, W.N. Lateral frontal cortex oxygenation changes during translation and language switching revealed by non-invasive near-infrared multi-point measurements. Brain Res. Bull. 2002, 59, 235–243. [Google Scholar] [CrossRef]
- Rinne, J.O.; Tommola, J.; Laine, M.; Krause, B.J.; Schmidt, D.; Kaasinen, V.; Teräs, M.; Sipilä, H.; Sunnari, M. The translating brain: Cerebral activation patterns during simultaneous interpretation. Neurosci. Lett. 2000, 294, 85–88. [Google Scholar] [CrossRef]
- Kurz, I. Watching the brain at work—An exploratory study of EEG changes during simultaneous interpreting (SI). Interpret. Newsl. 1995, 6, 3–16. [Google Scholar]
- García, A.M. The Neurocognition of Translation and Interpreting; John Benjamins: Amsterdam, The Netherlands; Philadelphia, PA, USA, 2019. [Google Scholar]
- Tomasi, D.; Volkow, N.D. Network connectivity predicts language processing in healthy adults. Hum. Brain Mapp. 2020, 41, 3696–3708. [Google Scholar] [CrossRef]
- Tomasi, D.; Volkow, N.D. Resting functional connectivity of language networks: Characterization and reproducibility. Mol. Psychiatry 2012, 17, 841–854. [Google Scholar] [CrossRef]
- Gullifer, J.W.; Chai, X.J.; Whitford, V.; Pivneva, I.; Baum, S.; Klein, D.; Titone, D. Bilingual experience and resting-state brain connectivity: Impacts of L2 age of acquisition and social diversity of language use on control networks. Neuropsychologia 2018, 117, 123–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Lu, J.; Zhang, H.; Zhang, J.; Yao, C.; Zhuang, D.; Qiu, T.; Guo, Q.; Hu, X.; Mao, Y.; et al. Direct evidence from intraoperative electrocortical stimulation indicates shared and distinct speech production center between Chinese and English languages. Hum. Brain Mapp. 2015, 36, 4972–4985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, D.; Milner, B.; Zatorre, R.J.; Meyer, E.; Evans, A.C. The neural substrates underlying word generation: A bilingual functional-imaging study. Proc. Natl. Acad. Sci. USA 1995, 92, 2899–2903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Wang, M.Y.; Li, D.; Yuan, Z. Optical mapping of brain activation during the English to Chinese and Chinese to English sight translation. Biomed. Opt. Express 2017, 8, 5399–5411. [Google Scholar] [CrossRef] [Green Version]
- Shinozuka, K.; Niioka, K.; Tokuda, T.; Kyutoku, Y.; Okuno, K.; Takahashi, T.; Dan, I. Language Familiarity and Proficiency Leads to Differential Cortical Processing During Translation Between Distantly Related Languages. Front. Hum. Neurosci. 2021, 15, 26. [Google Scholar] [CrossRef] [PubMed]
- Tommola, J.; Laine, M.; Sunnari, M.; Rinne, J.O. Images of shadowing and interpreting. Interpreting 2000, 5, 147–167. [Google Scholar] [CrossRef]
- Janyan, A.; Popivanov, I.; Andonova, E. Concreteness effect and word cognate status: ERPs in single word translation. In Brain Talk: Discourse with and in the Brain; Alter, K., Horne, M., Lindgren, M., Roll, M., von Koss Torkildsen, J., Eds.; Lunds Universitet: Lund, Sweden, 2009; pp. 21–30. [Google Scholar]
- Dronkers, N.F.; Plaisant, O.; Iba-Zizen, M.T.; Cabanis, E.A. Paul Broca’s historic cases: High resolution MR imaging of the brains of Leborgne and Lelong. Brain 2007, 130, 1432–1441. [Google Scholar] [CrossRef] [Green Version]
- Flinker, A.; Korzeniewska, A.; Shestyuk, A.Y.; Franaszczuk, P.J.; Dronkers, N.F.; Knight, R.T.; Crone, N.E. Redefining the role of Broca’s area in speech. Proc. Natl. Acad. Sci. USA 2015, 112, 2871–2875. [Google Scholar] [CrossRef] [Green Version]
- Tate, M.C.; Herbet, G.; Moritz-Gasser, S.; Tate, J.E.; Duffau, H. Probabilistic map of critical functional regions of the human cerebral cortex: Broca’s area revisited. Brain 2014, 137, 2773–2782. [Google Scholar] [CrossRef]
- Tettamanti, M.; Weniger, D. Broca’s area: A supramodal hierarchical processor? Cortex 2006, 42, 491–494. [Google Scholar] [CrossRef]
- Binder, J.R. Current controversies on Wernicke’s area and its role in language. Curr. Neurol. Neurosci. Rep. 2017, 17, 58. [Google Scholar] [CrossRef]
- Harpaz, Y.; Levkovitz, Y.; Lavidor, M. Lexical ambiguity resolution in Wernicke’s area and its right homologue. Cortex 2009, 45, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Kovelman, I.; Shalinsky, M.H.; Berens, M.S.; Petitto, L.A. Shining new light on the brain’s “bilingual signature”: A functional near infrared spectroscopy investigation of semantic processing. NeuroImage 2008, 39, 1457–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, C.J.; Green, D.W.; Von Studnitz, R. A functional imaging study of translation and language switching. Brain 1999, 122, 2221–2235. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, A.E.; Dapretto, M.; Mazziotta, J.; Bookheimer, S. Language switching and language representation in Spanish–English bilinguals: An fMRI study. Neuroimage 2001, 14, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.A.; Luciana, M. (Eds.) Handbook of Developmental Cognitive Neuroscience; MIT Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Hartwigsen, G.; Baumgaertner, A.; Price, C.J.; Koehnke, M.; Ulmer, S.; Siebner, H.R. Phonological decisions require both the left and right supramarginal gyri. Proc. Natl. Acad. Sci. USA 2010, 107, 16494–16499. [Google Scholar] [CrossRef] [Green Version]
- Brownsett, S.L.E.; Wise, R.J.S. The contribution of the parietal lobes to speaking and writing. Cereb. Cortex 2010, 20, 517–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.-J.; Kim, J.J.; Lee, S.-K.; Seok, J.H.; Chun, J.; Kim, D.I.; Lee, J.D. Corpus callosal connection mapping using cortical gray matter parcellation and DT-MRI. Hum. Brain Mapp. 2008, 29, 503–516. [Google Scholar] [CrossRef]
- Yuan, Z. Spatiotemporal and time-frequency analysis of functional near infrared spectroscopy brain signals using independent component analysis. J. Biomed. Opt 2013, 18, 106011. [Google Scholar] [CrossRef]
- Ibáñez, A.J.; Macizo, P.; Bajo, M.T. Language access and language selection in professional translators. Acta Psychol. 2010, 135, 257–266. [Google Scholar] [CrossRef]
- Yudes, C.; Macizo, P.; Bajo, T. The influence of expertise in simultaneous interpreting on non-verbal executive processes. Front. Psychol. 2011, 2, 309. [Google Scholar] [CrossRef] [Green Version]
- Hulstijn, J.H. Measuring second language proficiency. In Experimental Methods in Language Acquisition Research; Blom, E., Unsworth, S., Eds.; John Benjamins: Amsterdam, The Netherlands; Philadelphia, PA, USA, 2010; pp. 185–199. [Google Scholar]
- Scherer, L.C.; Fonseca, R.P.; Amiri, M.; Adrover-Roig, D.; Marcotte, K.; Giroux, F.; Senhadji, N.; Benali, H.; Lesage, F.; Ansaldo, A.I. Syntactic processing in bilinguals: An fNIRS study. Brain Lang. 2012, 121, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Ilg, G.; Lambert, S. Teaching consecutive interpreting. Interpreting 1996, 1, 69–99. [Google Scholar] [CrossRef]
- Lee, J. What skills do student interpreters need to learn in sight translation training? Meta 2012, 57, 694–714. [Google Scholar] [CrossRef] [Green Version]
- Agrifoglio, M. Sight translation and interpreting: A comparative analysis of constraints and failures. Interpreting 2004, 6, 43–67. [Google Scholar] [CrossRef]
- Kameyama, M.; Fukuda, M.; Uehara, T.; Mikuni, M. Sex and age dependencies of cerebral blood volume changes during cognitive activation: A multichannel near-infrared spectroscopy study. NeuroImage 2004, 22, 1715–1721. [Google Scholar] [CrossRef]
- Hori, H.; Nagamine, M.; Soshi, T.; Okabe, S.; Kim, Y.; Kunugi, H. Schizotypal traits in healthy women predict prefrontal activation patterns during a verbal fluency task: A near-infrared spectroscopy study. Neuropsychobiology 2008, 57, 61–69. [Google Scholar] [CrossRef]
- Schecklmann, M.; Ehlis, A.C.; Plichta, M.M.; Fallgatter, A.J. Influence of muscle activity on brain oxygenation during verbal fluency assessed with functional near-infrared spectroscopy. Neuroscience 2010, 171, 434–442. [Google Scholar] [CrossRef]
- Friederici, A.D.; Hahne, A.; Von Cramon, D.Y. First-pass versus second-pass parsing processes in a Wernicke’s and a Broca’s aphasic: Electrophysiological evidence for a double dissociation. Brain Lang. 1998, 62, 311–341. [Google Scholar] [CrossRef]
- Ye, J.C.; Tak, S.; Jang, K.E.; Jung, J.; Jang, J. NIRS-SPM: Statistical parametric mapping for near-infrared spectroscopy. NeuroImage 2009, 44, 428–447. [Google Scholar] [CrossRef]
- Xiao, X.; Zhu, H.; Liu, W.J.; Yu, X.T.; Duan, L.; Li, Z.; Zhu, C.Z. Semi-automatic 10/20 identification method for MRI-free probe placement in transcranial brain mapping techniques. Front. Neurosci. 2017, 11, 4. [Google Scholar] [CrossRef]
- Singh, A.K.; Okamoto, M.; Dan, H.; Jurcak, V.; Dan, I. Spatial rIGistration of multichannel multi-subject fNIRS data to MNI space without MRI. NeuroImage 2005, 27, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, Y. Functional near-infrared optical imaging: Utility and limitations in human brain mapping. Psychophysiology 2003, 40, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Kirilina, E.; Jelzow, A.; Heine, A.; Niessing, M.; Wabnitz, H.; Brühl, R.; Ittermann, B.; Jacobs, A.M.; Tachtsidis, I. The physiological origin of task-evoked systemic artefacts in functional near infrared spectroscopy. NeuroImage 2012, 61, 70–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Noah, J.A.; Joy Hirsch, J. Separation of the global and local components in functional near-infrared spectroscopy signals using principal component spatial filtering. Neurophotonics 2016, 3, 015004. [Google Scholar] [CrossRef]
- Cui, X.; Bray, S.; Reiss, A.L. Functional near infrared spectroscopy (NIRS) signal improvement based on negative correlation between oxygenated and deoxygenated hemoglobin dynamics. NeuroImage 2010, 49, 3039–3046. [Google Scholar] [CrossRef] [Green Version]
- Hertrich, I.; Dietrich, S.; Ackermann, H. The margins of the language network in the brain. Front. Commun. 2020, 5, 519955. [Google Scholar] [CrossRef]
- Ren, H.; Wang, M.; He, Y.; Du, Z.; Zhang, J.; Zhang, Z.; Li, D.; Yuan, Z. A novel phase analysis method for examining fNIRS neuroimaging data associated with Chinese/English sight translation. Behav. Brain Res. 2019, 361, 151–158. [Google Scholar] [CrossRef]
- Liu, H.; Cao, F. L1 and L2 processing in the bilingual brain: A meta-analysis of neuroimaging studies. Brain Lang. 2016, 159, 60–73. [Google Scholar] [CrossRef]
- Tan, L.H.; Laird, A.R.; Li, K.; Fox, P.T. Neuroanatomical correlates of phonological processing of Chinese characters and alphabetic words: A meta-analysis. Hum. Brain Mapp. 2005, 25, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Du, F.; Li, J.; Yu, H.; Tang, C.; Jiang, R. Functional magnetic resonance imaging based on Chinese tasks to protect language function in epileptics. Brain Behav. 2021, 11, e01979. [Google Scholar] [CrossRef]
- Liu, H.; Tong, J.; de Bruin, A.; Li, W.; He, Y.; Li, B. Is inhibition involved in voluntary language switching? Evidence from transcranial direct current stimulation over the right dorsolateral prefrontal cortex. Int. J. Psychophysiol. 2020, 147, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Zhang, J.; Couto, T.A.; Xu, S.; Luan, P.; Yuan, Z. Optical mapping of brain activation and connectivity in occipitotemporal cortex during Chinese character recognition. Brain Topogr. 2018, 31, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Zhang, X.; Noah, J.A.; Dravida, S.; Hirsch, J. Bidirectional connectivity between Broca’s area and Wernicke’s area during interactive verbal communication. Brain Connect. 2021. [Google Scholar] [CrossRef] [PubMed]
- Hickok, G.; Poeppel, D. The cortical organization of speech processing. Nat. Rev. Neurosci. 2007, 8, 393–402. [Google Scholar] [CrossRef]
- Hertrich, I.; Dietrich, S.; Ackermann, H. The role of the supplementary motor area for speech and language processing. Neurosci. Biobehav. Rev. 2016, 68, 602–610. [Google Scholar] [CrossRef]
- Ferbinteanu, J. Memory systems 2018-towards a new paradigm. Neurobiol. Learn. Mem. 2019, 157, 61–78. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | IG (n = 16) | NIG (n = 16) | ||
|---|---|---|---|---|
| Mean | SD | Mean | SD | |
| Age (years) | 25.00 | 1.08 | 23.88 | 1.59 |
| Education | Bachelor’s degree | Bachelor’s degree | ||
| Working memory span amplitude (maximum) | 2.75 | 0.45 | 2.88 | 0.72 |
| L2 proficiency score (maximum: 6) | 4.56 | 0.81 | 4.69 | 0.60 |
| Age of first acquisition of L2 (years) | 10.19 | 2.04 | 8.63 | 2.80 |
| Total time length since L2 acquisition (years) | 14.75 | 2.91 | 15.19 | 2.14 |
| Mandarin Sentences M (SD) | English Sentences M (SD) | p | |
|---|---|---|---|
| Word count | 10 (0.52) | 10 (0.35) | 1 |
| Notional word density | 78.95% (0.01) | 78.09% (0.01) | 0.71 |
| Pronoun density | 10.51% (0.00) | 11.67% (0.00) | 0.22 |
| Translatability | 3.76 (0.46) | 3.80 (0.92) | 0.86 |
| Word frequency | Mandarin sentences M (SD) | English sentences M (SD) | p |
| 1–1000 | 69.46% (0.01) | 68.60% (0.01) | 0.72 |
| 1001–2000 | 8.36% (0.00) | 8.40% (0.00) | 0.98 |
| 2001–3000 | 4.55% (0.00) | 4.14% (0.00) | 0.82 |
| 3001–4000 | 0.83% (0.00) | 1.67% (0.00) | 0.47 |
| 4001–5000 | 2.64% (0.00) | 2.93% (0.00) | 0.86 |
| 5000 plus | 14.15% (0.01) | 14.26% (0.01) | 0.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Hu, Y. Functional Connectivity Signatures Underlying Simultaneous Language Translation in Interpreters and Non-Interpreters of Mandarin and English: An fNIRS Study. Brain Sci. 2022, 12, 273. https://doi.org/10.3390/brainsci12020273
He Y, Hu Y. Functional Connectivity Signatures Underlying Simultaneous Language Translation in Interpreters and Non-Interpreters of Mandarin and English: An fNIRS Study. Brain Sciences. 2022; 12(2):273. https://doi.org/10.3390/brainsci12020273
Chicago/Turabian StyleHe, Yan, and Yinying Hu. 2022. "Functional Connectivity Signatures Underlying Simultaneous Language Translation in Interpreters and Non-Interpreters of Mandarin and English: An fNIRS Study" Brain Sciences 12, no. 2: 273. https://doi.org/10.3390/brainsci12020273
APA StyleHe, Y., & Hu, Y. (2022). Functional Connectivity Signatures Underlying Simultaneous Language Translation in Interpreters and Non-Interpreters of Mandarin and English: An fNIRS Study. Brain Sciences, 12(2), 273. https://doi.org/10.3390/brainsci12020273