
Increased Threshold and Reduced Firing Rate of Auditory Cortex Neurons after Cochlear Implant Insertion


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Audiogram
2.3. Cortical Surgery and Cochlear Implantation
2.4. Recording of Auditory Cortex Neurons
2.5. Experimental Protocol
2.6. Quantification of the Cortical Responses
2.7. Statistical Analysis
3. Results
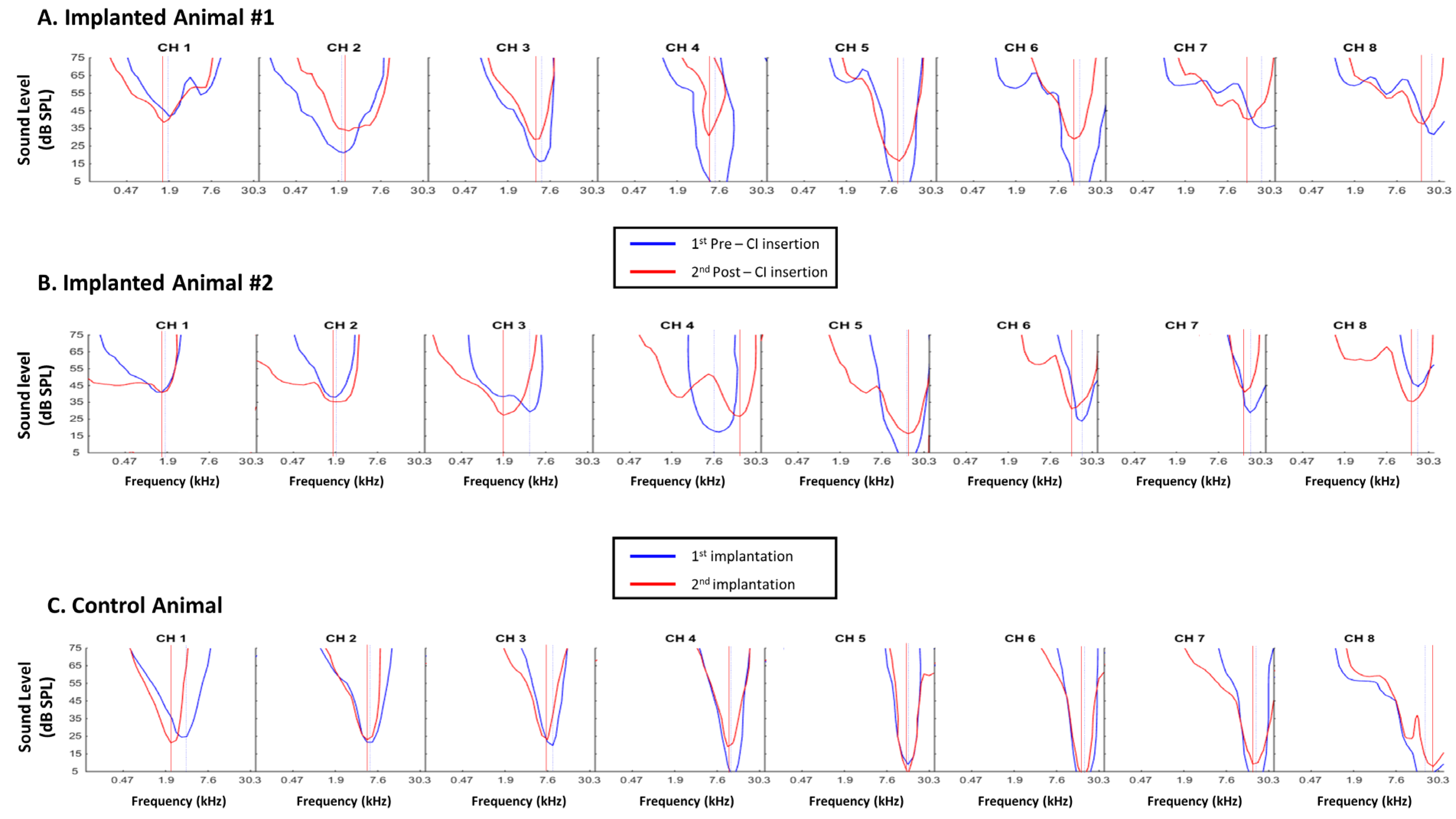
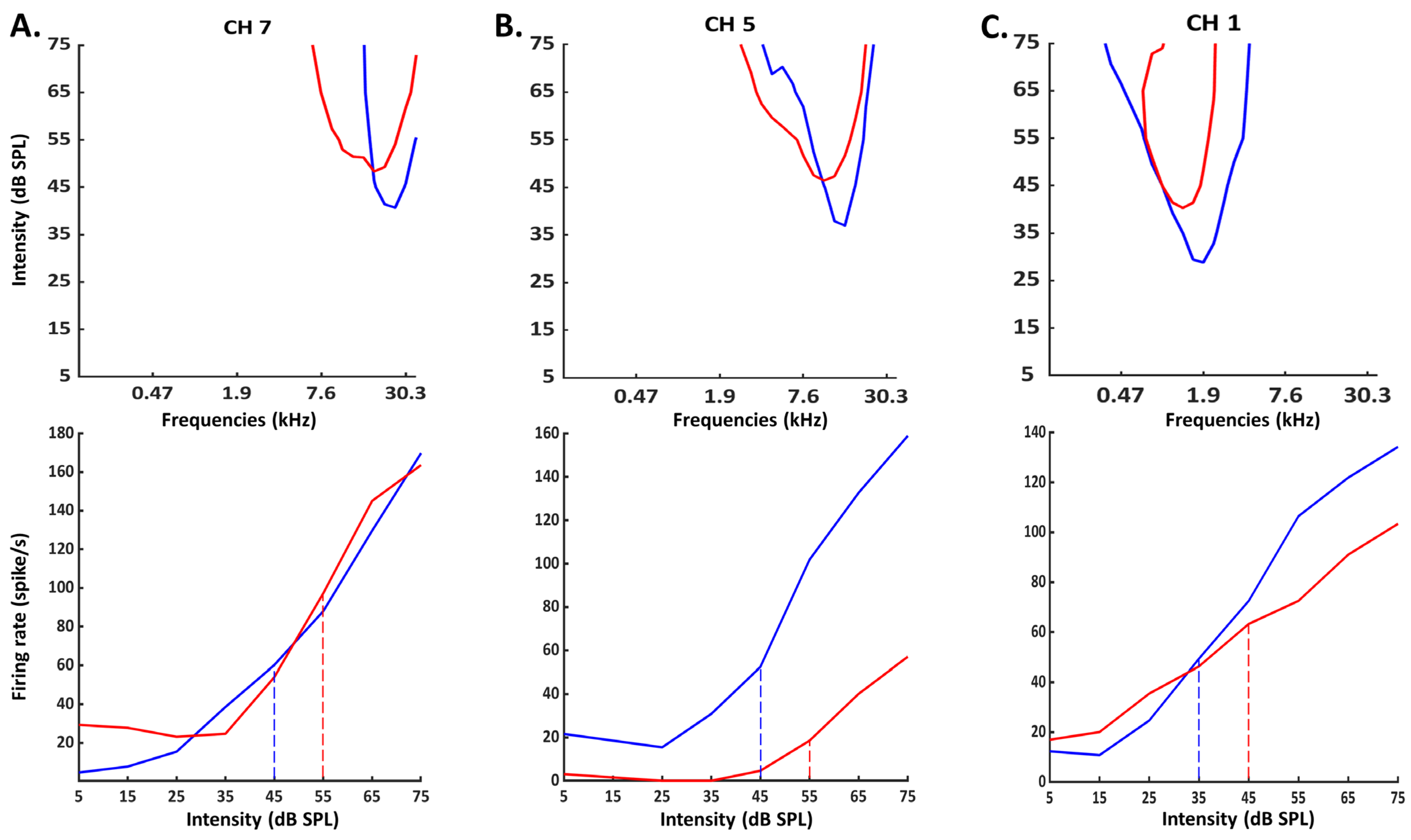
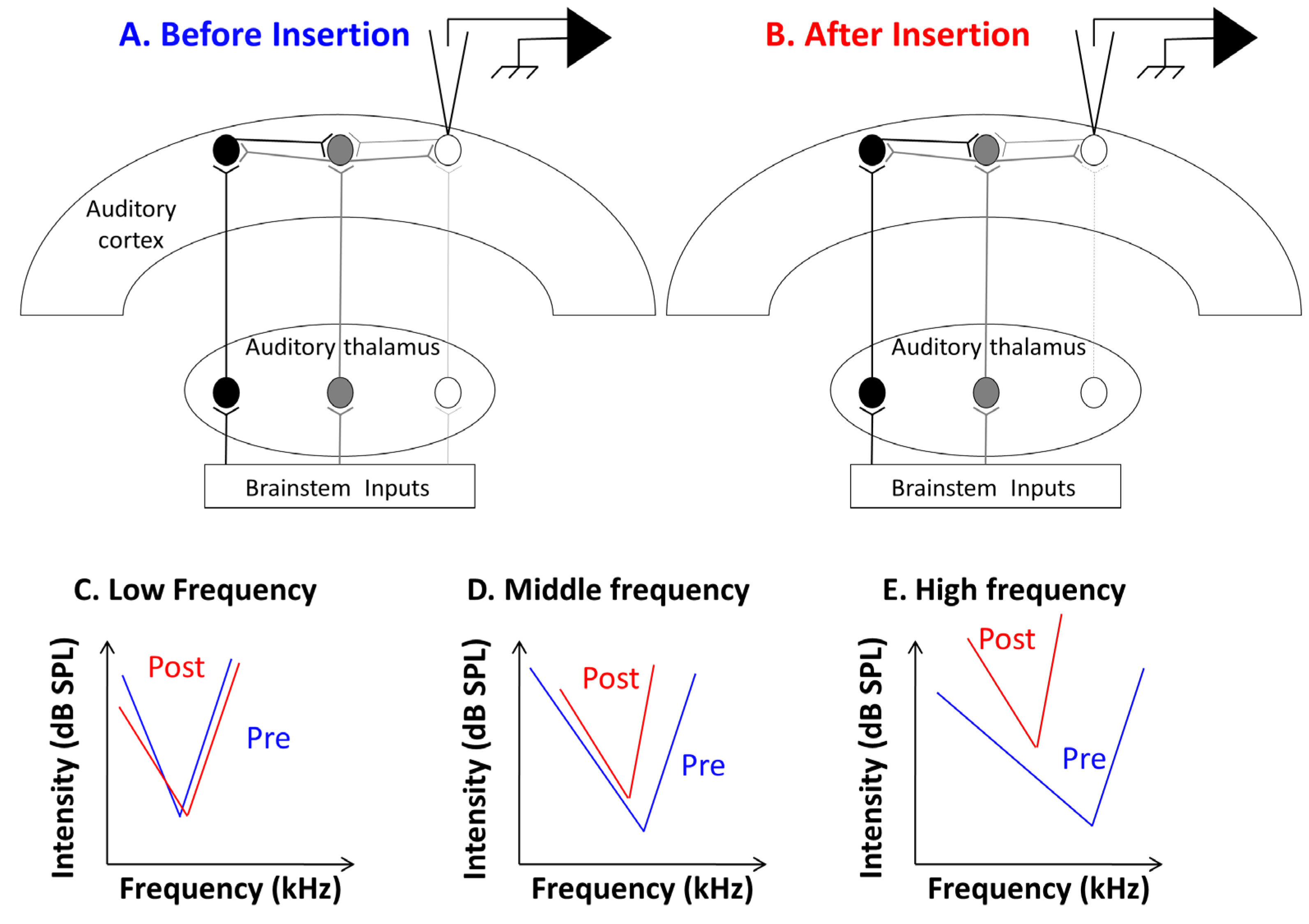
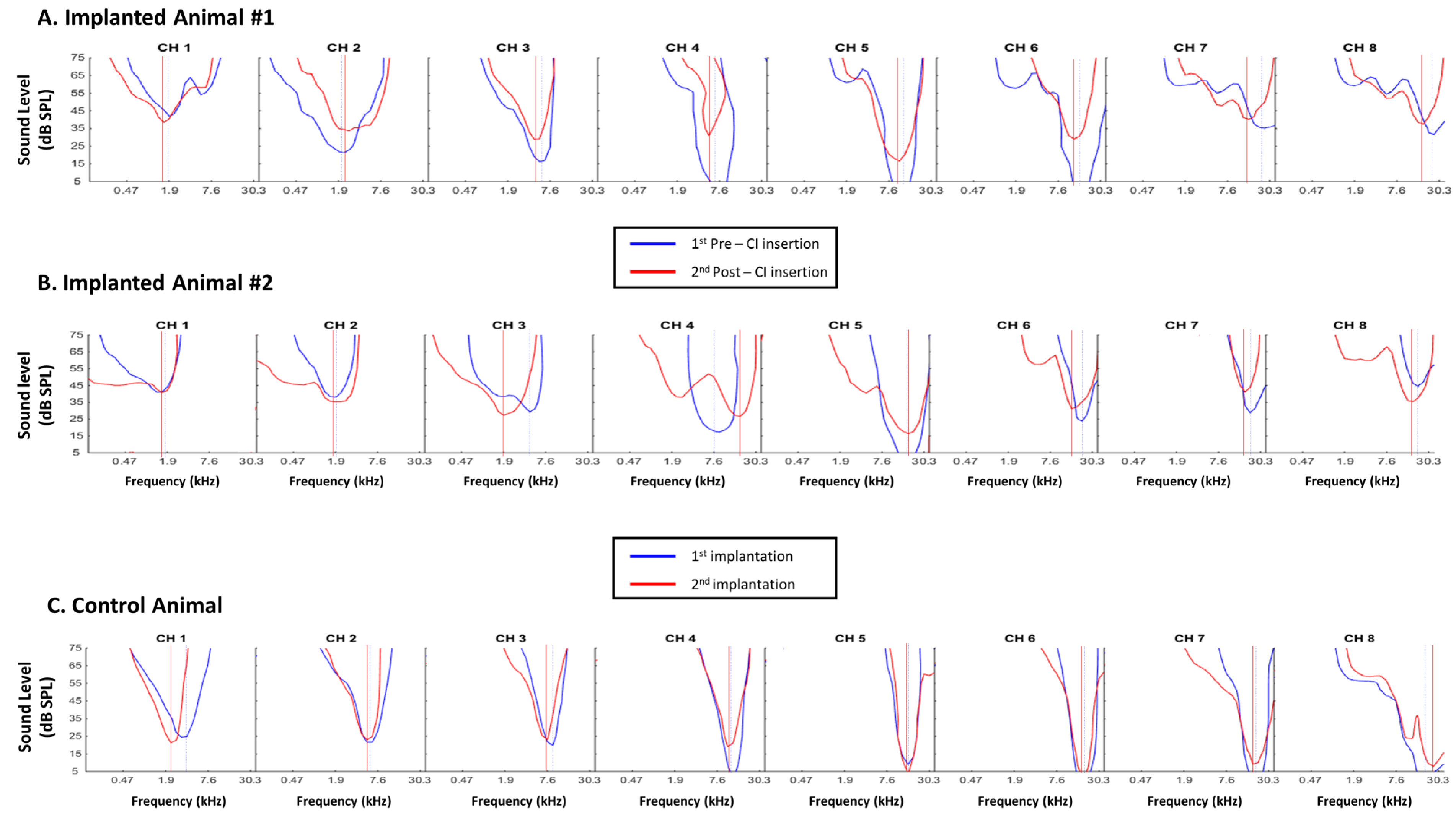
3.1. Consequences of Cochlear Implantation on Frequency Response Areas
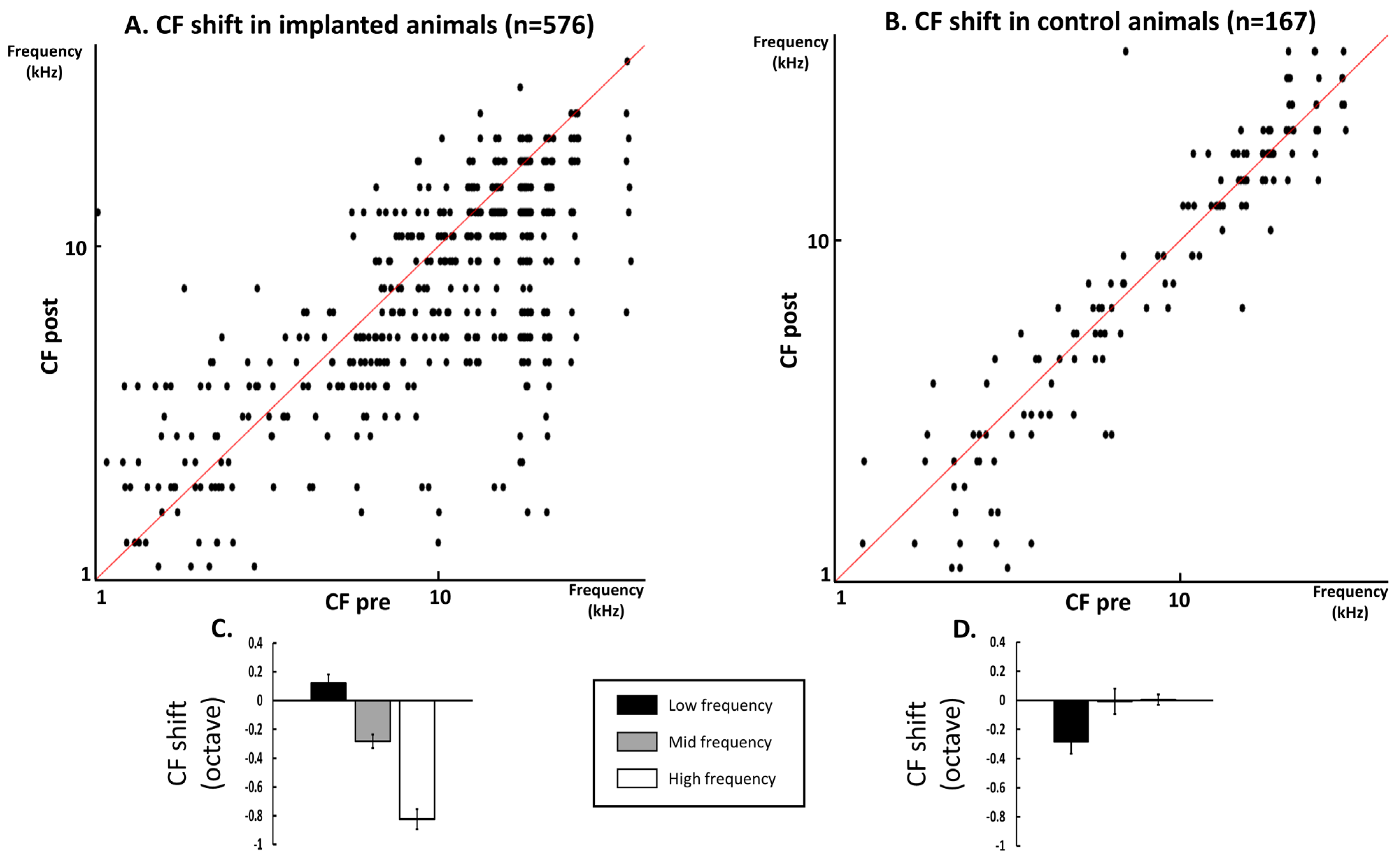
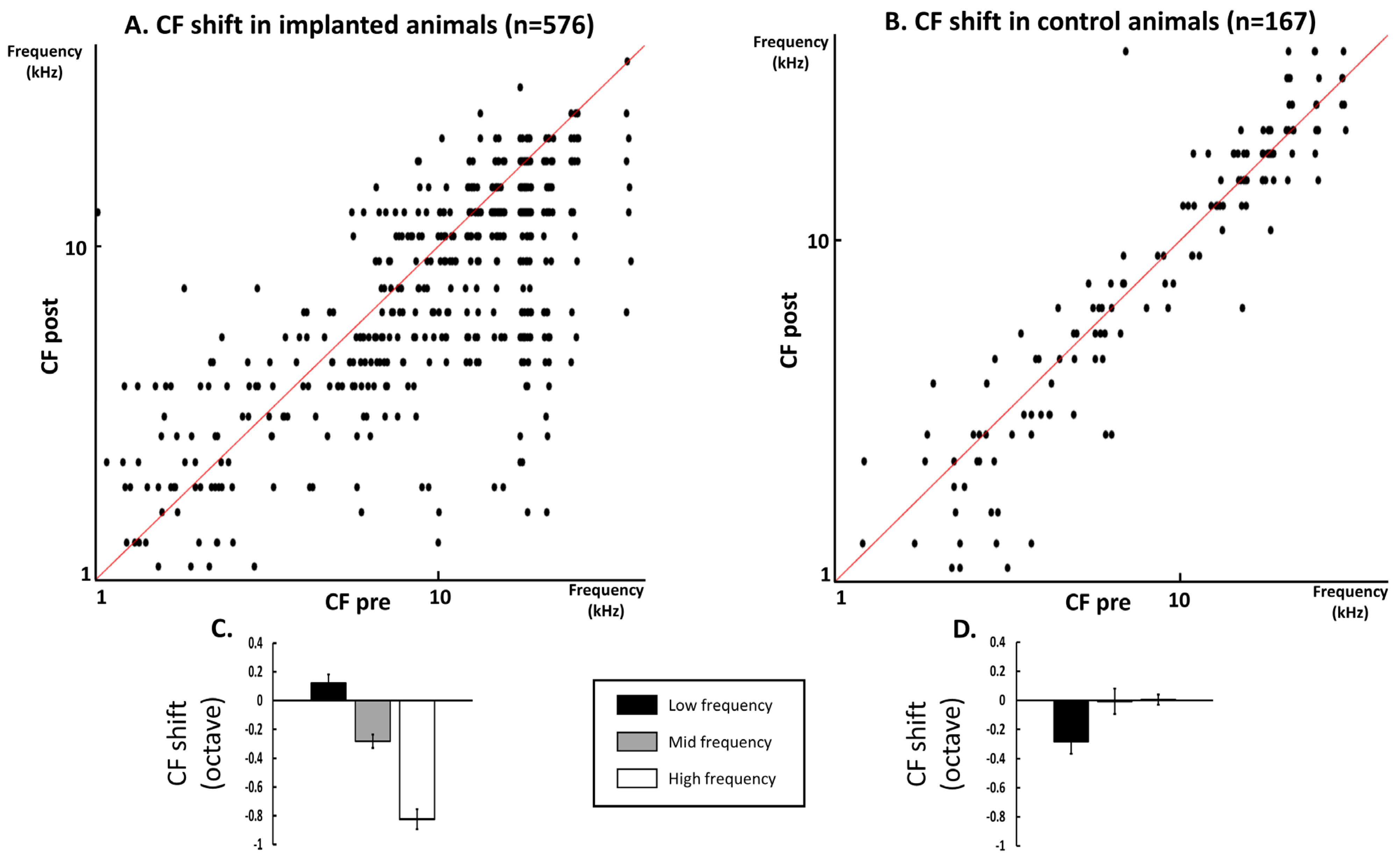
3.2. Quantification of the CF Values on the Whole Database
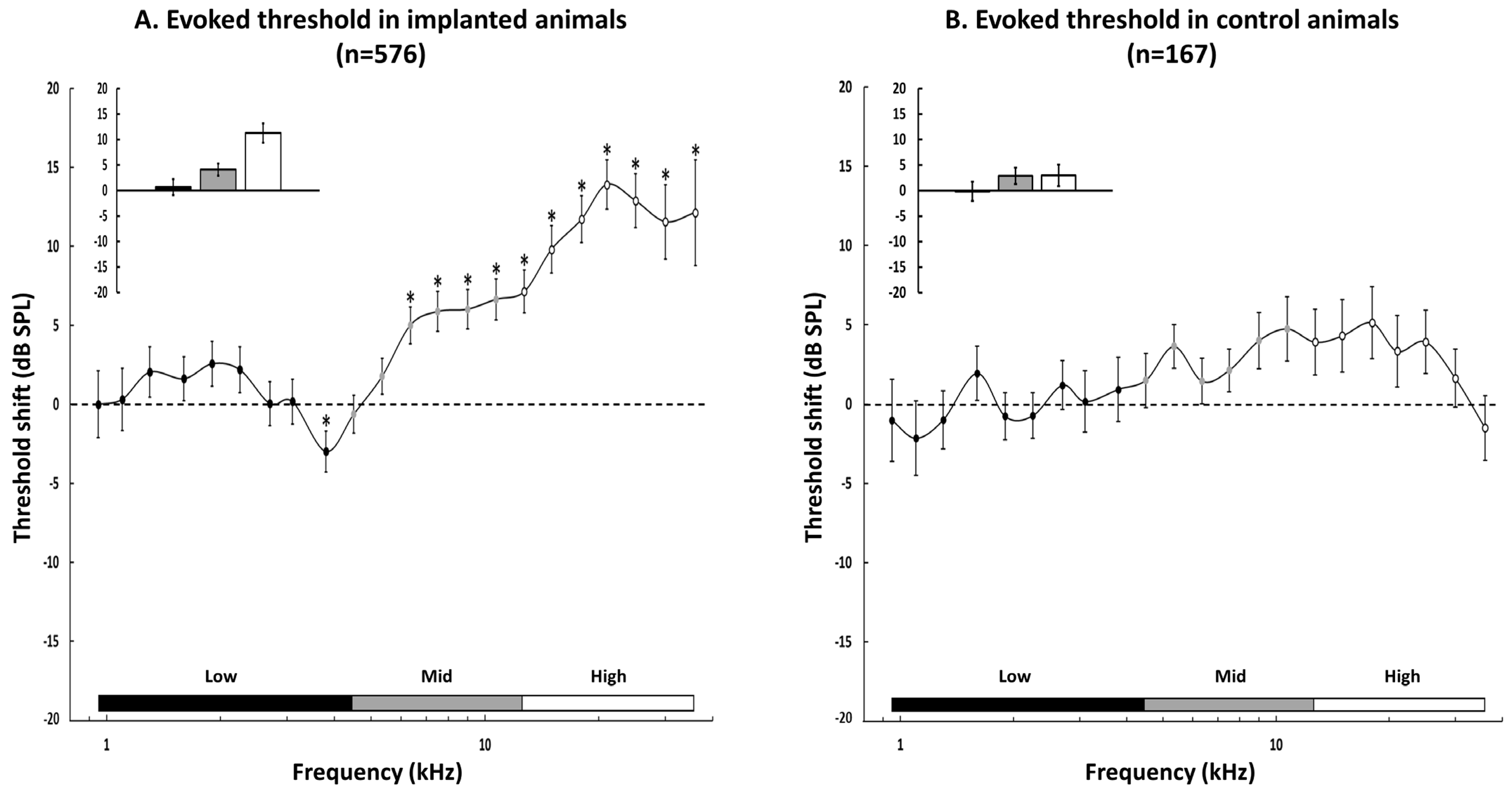
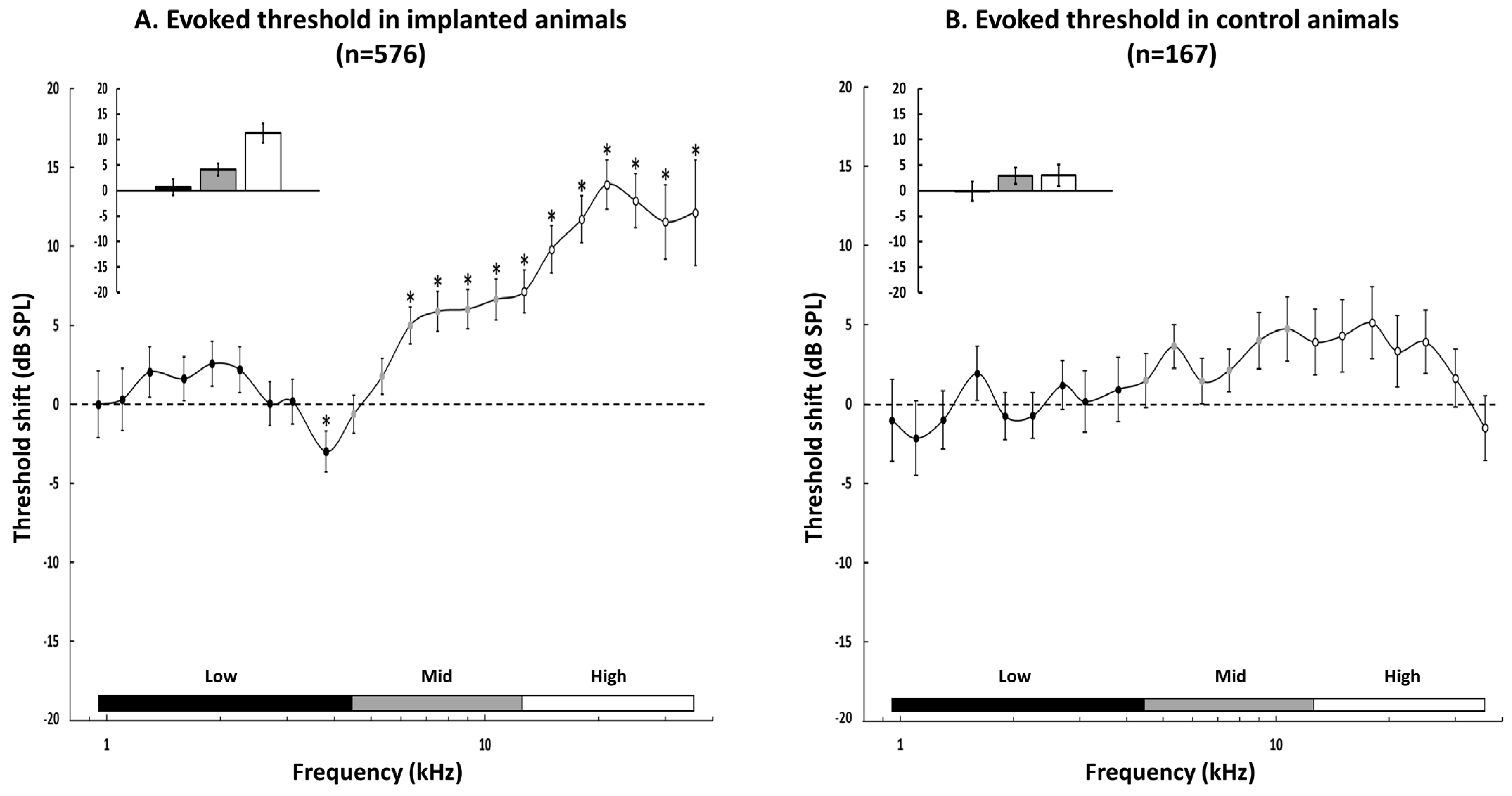
3.3. Group Data for the Changes in the Acoustic Threshold
3.4. Group Data for the Evoked Firing Rate
4. Discussion
4.1. Potential Neuronal Mechanisms Underlying the Consequences of CI Insertion
4.2. Clinical Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, Y.; Mosnier, I.; Borel, S.; Ambert-Dahan, E.; Bouccara, D.; Bozorg-Grayeli, A.; Ferrary, E.; Sterkers, O. Evolution of electrode array diameter for hearing preservation in cochlear implantation. Acta Oto-Laryngol. 2013, 133, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Mamelle, E.; Granger, B.; Sterkers, O.; Lahlou, G.; Ferrary, E.; Nguyen, Y.; Mosnier, I. Long-term residual hearing in cochlear implanted adult patients who were candidates for electro-acoustic stimulation. Eur. Arch. Oto-Rhino-Laryngol. 2020, 277, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Gantz, B.J.; Hansen, M.; Turner, C.W.; Oleson, J.J.; Reiss, L.; Parkinson, A.J. Hybrid 10 Clinical Trial. Audiol. Neurotol. 2009, 14, 32–38. [Google Scholar] [CrossRef]
- Maria, P.L.S.; Domville-Lewis, C.; Sucher, C.M.; Chester-Browne, R.; Atlas, M.D. Hearing Preservation Surgery for Cochlear Implantation—Hearing and Quality of Life After 2 Years. Otol. Neurotol. 2013, 34, 526–531. [Google Scholar] [CrossRef]
- Gstoettner, W.; Helbig, S.; Settevendemie, C.; Baumann, U.; Wagenblast, J.; Arnoldner, C. A new electrode for residual hearing preservation in cochlear implantation: First clinical results. Acta Oto-Laryngol. 2009, 129, 372–379. [Google Scholar] [CrossRef]
- Maria, P.L.S.; Gluth, M.B.; Yuan, Y.; Atlas, M.D.; Blevins, N.H. Hearing Preservation Surgery for Cochlear Implantation. Otol. Neurotol. 2014, 35, e256–e269. [Google Scholar] [CrossRef]
- Elliott, S.J.; Ni, G.; Verschuur, C.A. Modelling the effect of round window stiffness on residual hearing after cochlear implantation. Hear. Res. 2016, 341, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Snels, C.; IntHout, J.; Mylanus, E.; Huinck, W.; Dhooge, I. Hearing Preservation in Cochlear Implant Surgery: A Meta-Analysis. Otol. Neurotol. 2019, 40, 145–153. [Google Scholar] [CrossRef]
- Jia, H.; François, F.; Bourien, J.; Eybalin, M.; Lloyd, R.; Van De Water, T.; Puel, J.-L.; Venail, F. Prevention of trauma-induced cochlear fibrosis using intracochlear application of anti-inflammatory and antiproliferative drugs. Neuroscience 2016, 316, 261–278. [Google Scholar] [CrossRef]
- Wilk, M.; Hessler, R.; Mugridge, K.; Jolly, C.; Fehr, M.; Lenarz, T.; Scheper, V. Impedance Changes and Fibrous Tissue Growth after Cochlear Implantation Are Correlated and Can Be Reduced Using a Dexamethasone Eluting Electrode. PLoS ONE 2016, 11, e0147552. [Google Scholar] [CrossRef]
- Giordano, P.; Hatzopoulos, S.; Giarbini, N.; Prosser, S.; Petruccelli, J.; Simoni, E.; Faccioli, C.; Astolfi, L.; Martini, A. A Soft-Surgery Approach to Minimize Hearing Damage Caused by the Insertion of a Cochlear Implant Electrode. Otol. Neurotol. 2014, 35, 1440–1445. [Google Scholar] [CrossRef] [PubMed]
- Drouillard, M.; Torres, R.; Mamelle, E.; De Seta, D.; Sterkers, O.; Ferrary, E.; Nguyen, Y. Influence of electrode array stiffness and diameter on hearing in cochlear implanted guinea pig. PLoS ONE 2017, 12, e0183674. [Google Scholar] [CrossRef] [Green Version]
- Adel, Y.; Tillein, J.; Petzold, H.; Weissgerber, T.; Baumann, U. Band-Limited Chirp-Evoked Compound Action Potential in Guinea Pig: Comprehensive Neural Measure for Cochlear Implantation Monitoring. Ear Hear. 2020, 42, 142–162. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Baumhoff, P.; Kral, A. Cochlear Implant Stimulation of a Hearing Ear Generates Separate Electrophonic and Electroneural Responses. J. Neurosci. 2016, 36, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Kylén, P.; Stjernvall, J.-E.; Arlinger, S. Variables Affecting the Drill-Generated Noise Levels in Ear Surgery. Acta Oto-Laryngol. 1977, 84, 252–259. [Google Scholar] [CrossRef]
- Pau, H.W.; Just, T.; Bornitz, M.; Lasurashvilli, N.; Zahnert, T. Noise Exposure of the Inner Ear During Drilling a Cochleostomy for Cochlear Implantation. Laryngoscope 2007, 117, 535–540. [Google Scholar] [CrossRef]
- Gourévitch, B.; Edeline, J.-M. Age-related changes in the guinea pig auditory cortex: Relationship with brainstem changes and comparison with tone-induced hearing loss. Eur. J. Neurosci. 2011, 34, 1953–1965. [Google Scholar] [CrossRef]
- Occelli, F.; Hasselmann, F.; Bourien, J.; Eybalin, M.; Puel, J.-L.; Desvignes, N.; Wiszniowski, B.; Edeline, J.-M.; Gourévitch, B. Age-related Changes in Auditory Cortex Without Detectable Peripheral Alterations: A Multi-level Study in Sprague–Dawley Rats. Neuroscience 2019, 404, 184–204. [Google Scholar] [CrossRef] [Green Version]
- Rutkowski, R.G.; Weinberger, N.M. Encoding of learned importance of sound by magnitude of representational area in primary auditory cortex. Proc. Natl. Acad. Sci. USA 2005, 102, 13664–13669. [Google Scholar] [CrossRef] [Green Version]
- Recanzone, G.H.; Schreiner, C.; Merzenich, M.M. Plasticity in the frequency representation of primary auditory cortex following discrimination training in adult owl monkeys. J. Neurosci. 1993, 13, 87–103. [Google Scholar] [CrossRef]
- Fernández, C. Dimensions of the Cochlea (Guinea Pig). J. Acoust. Soc. Am. 1952, 24, 519–523. [Google Scholar] [CrossRef]
- Greenwood, D.D. A cochlear frequency-position function for several species—29 years later. J. Acoust. Soc. Am. 1990, 87, 2592–2605. [Google Scholar] [CrossRef]
- Gourévitch, B.; Doisy, T.; Avillac, M.; Edeline, J.-M. Follow-up of latency and threshold shifts of auditory brainstem responses after single and interrupted acoustic trauma in guinea pig. Brain Res. 2009, 1304, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Aushana, Y.; Souffi, S.; Edeline, J.-M.; Lorenzi, C.; Huetz, C. Robust Neuronal Discrimination in Primary Auditory Cortex Despite Degradations of Spectro-temporal Acoustic Details: Comparison Between Guinea Pigs with Normal Hearing and Mild Age-Related Hearing Loss. J. Assoc. Res. Otolaryngol. 2018, 19, 163–180. [Google Scholar] [CrossRef]
- Souffi, S.; Lorenzi, C.; Varnet, L.; Huetz, C.; Edeline, J.-M. Noise-Sensitive but More Precise Subcortical Representations Coexist with Robust Cortical Encoding of Natural Vocalizations. J. Neurosci. 2020, 40, 5228–5246. [Google Scholar] [CrossRef]
- Souffi, S.; Lorenzi, C.; Huetz, C.; Edeline, J.-M. Robustness to Noise in the Auditory System: A Distributed and Predictable Property. ENEURO 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Gaucher, Q.; Edeline, J.-M. Stimulus-specific effects of noradrenaline in auditory cortex: Implications for the discrimination of communication sounds. J. Physiol. 2015, 593, 1003–1020. [Google Scholar] [CrossRef] [Green Version]
- Occelli, F.; Suied, C.; Pressnitzer, D.; Edeline, J.-M.; Gourévitch, B. A Neural Substrate for Rapid Timbre Recognition? Neural and Behavioral Discrimination of Very Brief Acoustic Vowels. Cereb. Cortex 2016, 26, 2483–2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edeline, J.-M.; Pham, P.; Weinberger, N.M. Rapid development of learning-induced receptive field plasticity in the auditory cortex. Behav. Neurosci. 1993, 107, 539–551. [Google Scholar] [CrossRef]
- Edeline, J.-M.; Dutrieux, G.; Manunta, Y.; Hennevin, E. Diversity of receptive field changes in auditory cortex during natural sleep. Eur. J. Neurosci. 2001, 14, 1865–1880. [Google Scholar] [CrossRef]
- Kaur, S.; Lazar, R.; Metherate, R. Intracortical Pathways Determine Breadth of Subthreshold Frequency Receptive Fields in Primary Auditory Cortex. J. Neurophysiol. 2004, 91, 2551–2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metherate, R.; Kaur, S.; Kawai, H.; Lazar, R.; Liang, K.; Rose, H.J. Spectral integration in auditory cortex: Mechanisms and modulation. Hear. Res. 2005, 206, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.W.; Reiss, L.; Gantz, B.J. Combined acoustic and electric hearing: Preserving residual acoustic hearing. Hear. Res. 2008, 242, 164–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Seta, D.; Torres, R.; Russo, F.Y.; Ferrary, E.; Kazmitcheff, G.; Heymann, D.; Amiaud, J.; Sterkers, O.; Bernardeschi, D.; Nguyen, Y. Damage to inner ear structure during cochlear implantation: Correlation between insertion force and radio-histological findings in temporal bone specimens. Hear. Res. 2017, 344, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, R.; Drouillard, M.; De Seta, D.; Bensimon, J.-L.; Ferrary, E.; Sterkers, O.; Bernardeschi, D.; Nguyen, Y. Cochlear Implant Insertion Axis into the Basal Turn: A Critical Factor in Electrode Array Translocation. Otol. Neurotol. 2018, 39, 168–176. [Google Scholar] [CrossRef]
- Torres, R.; Jia, H.; Drouillard, M.; Bensimon, J.-L.; Sterkers, O.; Ferrary, E.; Nguyen, Y. An Optimized Robot-Based Technique for Cochlear Implantation to Reduce Array Insertion Trauma. Otolaryngol. Neck Surg. 2018, 159, 900–907. [Google Scholar] [CrossRef]
- Robertson, D.; Irvine, D.R.F. Plasticity of frequency organization in auditory cortex of guinea pigs with partial unilateral deafness. J. Comp. Neurol. 1989, 282, 456–471. [Google Scholar] [CrossRef]
- Rajan, R.; Irvine, D.R.F.; Wise, L.Z.; Heil, P. Effect of unilateral partial cochlear lesions in adult cats on the representation of lesioned and unlesioned cochleas in primary auditory cortex. J. Comp. Neurol. 1993, 338, 17–49. [Google Scholar] [CrossRef]
- Wang, J.; Salvi, R.J.; Powers, N. Plasticity of response properties of inferior colliculus neurons following acute cochlear damage. J. Neurophysiol. 1996, 75, 171–183. [Google Scholar] [CrossRef]
- Rajan, R.; Irvine, D. Neuronal responses across cortical field A1 in plasticity induced by peripheral auditory organ damage. Audiol. Neurotol. 1998, 3, 123–144. [Google Scholar] [CrossRef]
- Wang, Y.; Hirose, K.; Liberman, M.C. Dynamics of Noise-Induced Cellular Injury and Repair in the Mouse Cochlea. J. Assoc. Res. Otolaryngol. 2002, 3, 248–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irvine, D.R.; Rajan, R.; Smith, S. Effects of restricted cochlear lesions in adult cats on the frequency organization of the inferior colliculus. J. Comp. Neurol. 2003, 467, 354–374. [Google Scholar] [CrossRef] [PubMed]
- Kamke, M.R.; Brown, M.; Irvine, D. Plasticity in the tonotopic organization of the medial geniculate body in adult cats following restricted unilateral cochlear lesions. J. Comp. Neurol. 2003, 459, 355–367. [Google Scholar] [CrossRef]
- Noreña, A.J.; Tomita, M.; Eggermont, J.J. Neural Changes in Cat Auditory Cortex After a Transient Pure-Tone Trauma. J. Neurophysiol. 2003, 90, 2387–2401. [Google Scholar] [CrossRef]
- Tan, A.; Wehr, M. Balanced tone-evoked synaptic excitation and inhibition in mouse auditory cortex. Neuroscience 2009, 163, 1302–1315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.I.; Tan, A.; Schreiner, C.; Merzenich, M.M. Topography and synaptic shaping of direction selectivity in primary auditory cortex. Nature 2003, 424, 201–205. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, B.-H.; Wu, G.; Kim, Y.-J.; Xiao, Z.; Tao, H.W.; Zhang, L.I. Preceding Inhibition Silences Layer 6 Neurons in Auditory Cortex. Neuron 2010, 65, 706–717. [Google Scholar] [CrossRef] [Green Version]
- Gaucher, Q.; Huetz, C.; Gourévitch, B.; Edeline, J.-M. Cortical Inhibition Reduces Information Redundancy at Presentation of Communication Sounds in the Primary Auditory Cortex. J. Neurosci. 2013, 33, 10713–10728. [Google Scholar] [CrossRef] [Green Version]
- Gaucher, Q.; Edeline, J.-M.; Gourévitch, B. How different are the local field potentials and spiking activities? Insights from multi-electrodes arrays. J. Physiol. 2012, 106, 93–103. [Google Scholar] [CrossRef]
- Brosch, M.; Schreiner, C.E. Time Course of Forward Masking Tuning Curves in Cat Primary Auditory Cortex. J. Neurophysiol. 1997, 77, 923–943. [Google Scholar] [CrossRef]
- Noreña, A.; Eggermont, J.J. Comparison between local field potentials and unit cluster activity in primary auditory cortex and anterior auditory field in the cat. Hear. Res. 2002, 166, 202–213. [Google Scholar] [CrossRef]
- Eggermont, J.J.; Munguia, R.; Pienkowski, M.; Shaw, G. Comparison of LFP-Based and Spike-Based Spectro-Temporal Receptive Fields and Cross-Correlation in Cat Primary Auditory Cortex. PLoS ONE 2011, 6, e20046. [Google Scholar] [CrossRef] [PubMed]
- Lenarz, T.; Buechner, A.; Lesinski-Schiedat, A.; Timm, M.; Salcher, R. Hearing Preservation with a New Atraumatic Lateral Wall Electrode. Otol. Neurotol. 2020, 41, e993–e1003. [Google Scholar] [CrossRef] [PubMed]
- Kwacz, M.; Mrowka, M.; Wysocki, J. Round Window Membrane Motion before and after Stapedotomy Surgery—An Experi-mental Study. Acta Bioeng. Biomech. 2011, 13, 27–33. [Google Scholar]
- Vittoria, S.; Lahlou, G.; Torres, R.; Daoudi, H.; Mosnier, I.; Mazalaigue, S.; Ferrary, E.; Nguyen, Y.; Sterkers, O. Robot-based assistance in middle ear surgery and cochlear implantation: First clinical report. Eur. Arch. Oto-Rhino-Laryngol. 2021, 278, 77–85. [Google Scholar] [CrossRef]
- Daoudi, H.; Lahlou, G.; Torres, R.; Sterkers, O.; Lefeuvre, V.; Ferrary, E.; Mosnier, I.; Nguyen, Y. Robot-assisted Cochlear Implant Electrode Array Insertion in Adults: A Comparative Study with Manual Insertion. Otol. Neurotol. 2021, 42, e438–e444. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Partouche, E.; Adenis, V.; Gnansia, D.; Stahl, P.; Edeline, J.-M. Increased Threshold and Reduced Firing Rate of Auditory Cortex Neurons after Cochlear Implant Insertion. Brain Sci. 2022, 12, 205. https://doi.org/10.3390/brainsci12020205
Partouche E, Adenis V, Gnansia D, Stahl P, Edeline J-M. Increased Threshold and Reduced Firing Rate of Auditory Cortex Neurons after Cochlear Implant Insertion. Brain Sciences. 2022; 12(2):205. https://doi.org/10.3390/brainsci12020205
Chicago/Turabian StylePartouche, Elie, Victor Adenis, Dan Gnansia, Pierre Stahl, and Jean-Marc Edeline. 2022. "Increased Threshold and Reduced Firing Rate of Auditory Cortex Neurons after Cochlear Implant Insertion" Brain Sciences 12, no. 2: 205. https://doi.org/10.3390/brainsci12020205
APA StylePartouche, E., Adenis, V., Gnansia, D., Stahl, P., & Edeline, J.-M. (2022). Increased Threshold and Reduced Firing Rate of Auditory Cortex Neurons after Cochlear Implant Insertion. Brain Sciences, 12(2), 205. https://doi.org/10.3390/brainsci12020205