

Hand and Foot Selection in Mental Body Rotations Involves Motor-Cognitive Interactions


Abstract
1. Introduction
2. Study 1
2.1. Methods of Study 1
2.1.1. Participants of Study 1
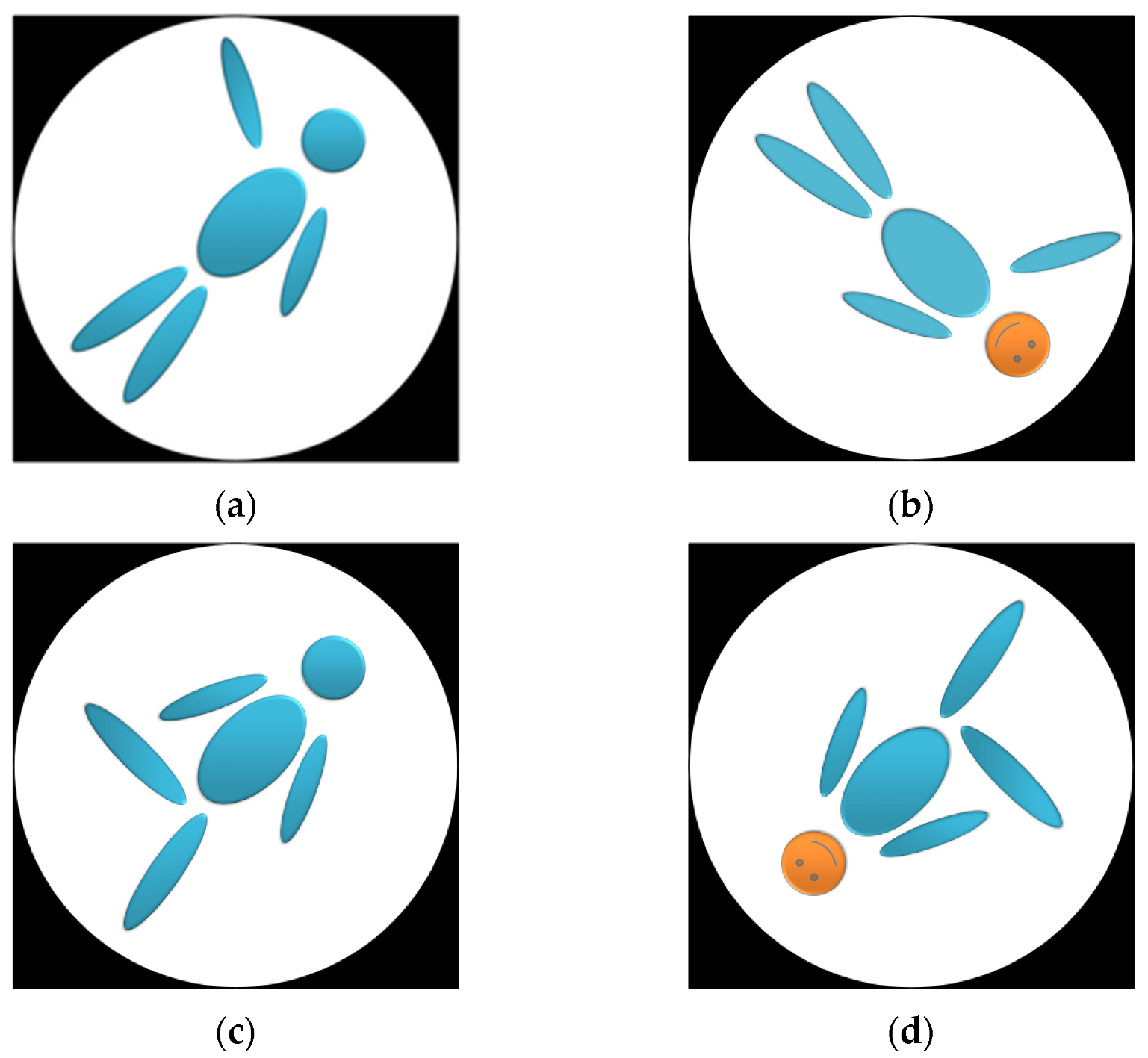
2.1.2. The Stimulus Material of Study 1
2.1.3. Task and Procedure of Study 1
2.1.4. Data Analysis of Study 1
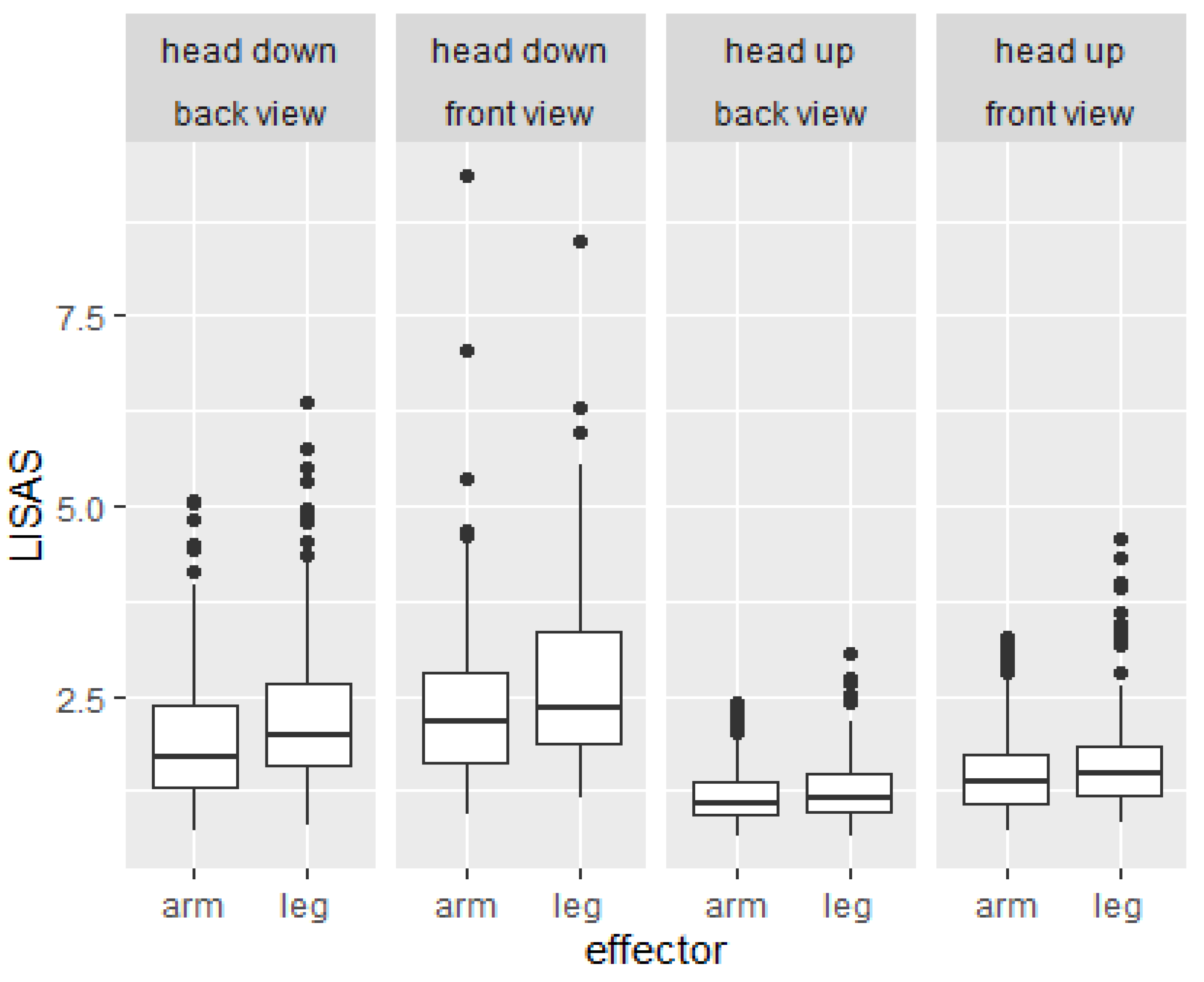
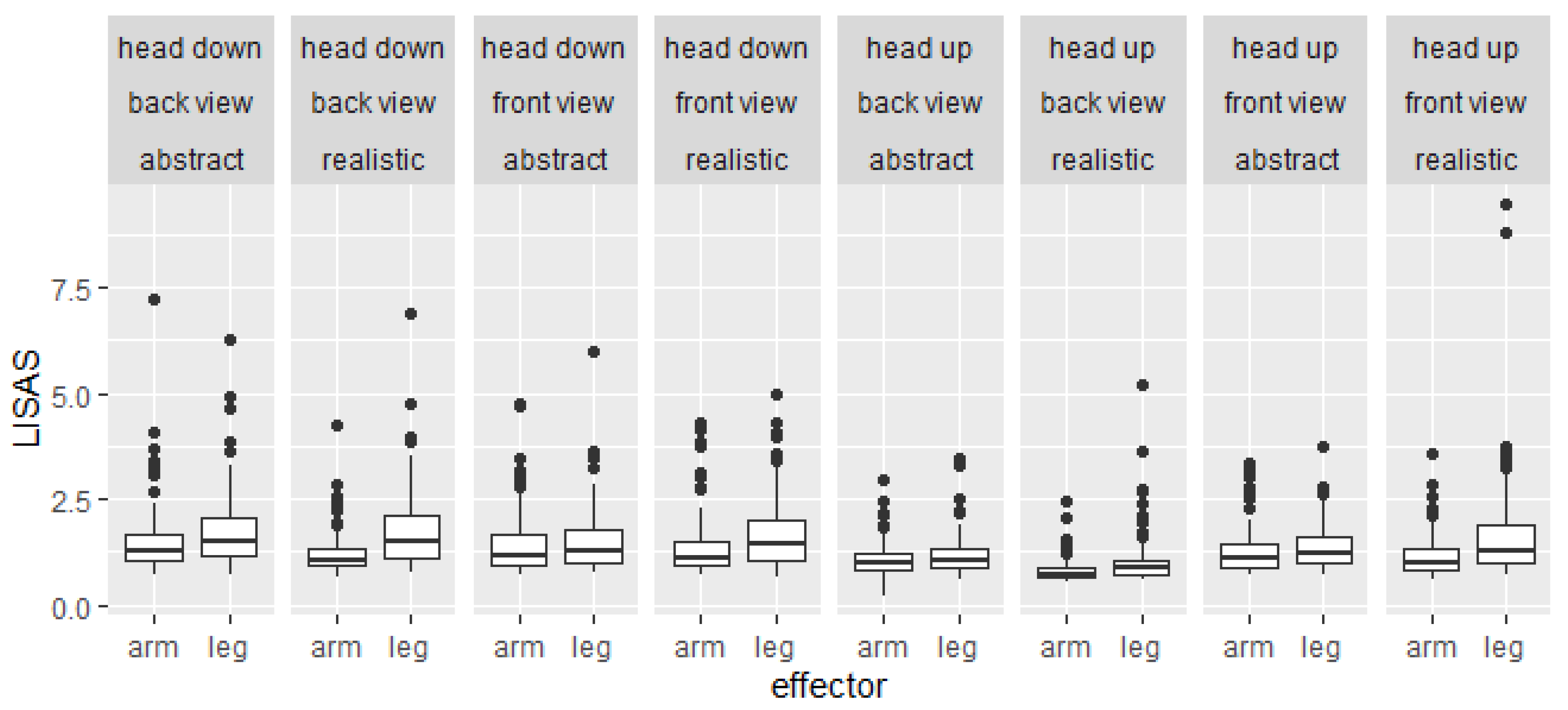
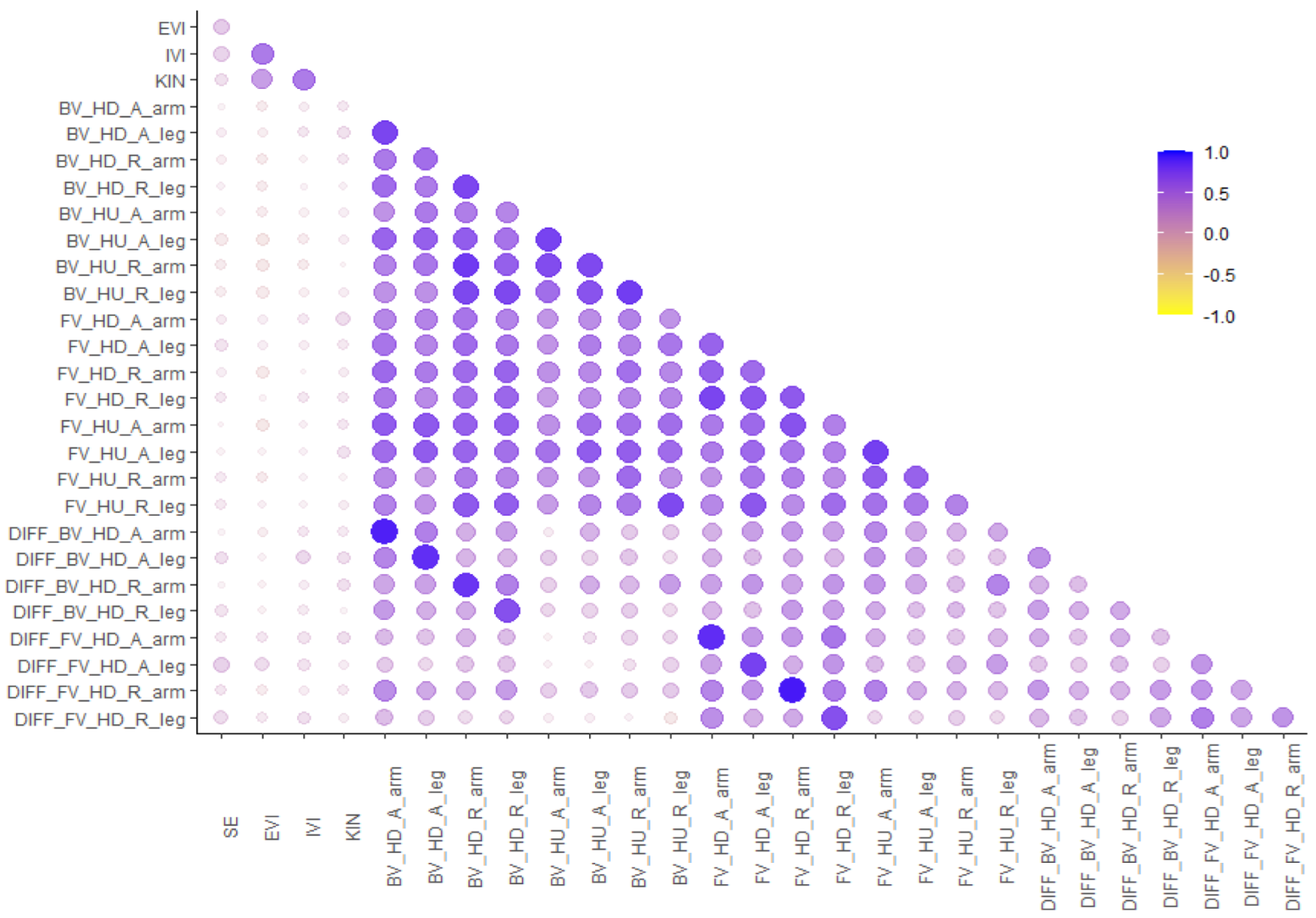
2.2. Results of Study 1
2.3. Discussion of Study 1
3. Study 2
3.1. Methods of Study 2
3.1.1. Participants of Study 2
3.1.2. The Stimulus Material of Study 2
3.1.3. Task and Procedure of Study 2
3.1.4. Data Analysis of Study 2
3.2. Results of Study 2
3.3. Discussion of Study 2
4. General Discussion
Limitations and Perspectives
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeannerod, M. Neural Simulation of Action: A Unifying Mechanism for Motor Cognition. Neuroimage 2001, 14, S103–S109. [Google Scholar] [CrossRef] [PubMed]
- Iachini, T. Mental Imagery and Embodied Cognition: A Multimodal Approach. J. Ment. Imag. 2011, 35, 1–66. [Google Scholar]
- Kosslyn, S.M.; Brunn, J.; Cave, K.R.; Wallach, R.W. Individual Differences in Mental Imagery Ability: A Computational Analysis. Cognition 1984, 18, 195–243. [Google Scholar] [CrossRef]
- Dahm, S.F. On the Assessment of Motor Imagery Ability: A Research Commentary. Imagin. Cogn. Pers. 2020, 39, 397–408. [Google Scholar] [CrossRef]
- Dahm, S.F.; Rieger, M. Is Imagery Better than Reality? Performance in Imagined Dart Throwing. Hum. Mov. Sci. 2019, 66, 38–52. [Google Scholar] [CrossRef]
- Kilteni, K.; Andersson, B.J.; Houborg, C.; Ehrsson, H.H. Motor Imagery Involves Predicting the Sensory Consequences of the Imagined Movement. Nat. Commun. 2018, 9, 1617. [Google Scholar] [CrossRef]
- Roberts, R.; Callow, N.; Hardy, L.; Markland, D.; Bringer, J. Movement Imagery Ability: Development and Assessment of a Revised Version of the Vividness of Movement Imagery Questionnaire. J. Sport Exerc. Psychol. 2008, 30, 200–221. [Google Scholar] [CrossRef]
- Grush, R. The Emulation Theory of Representation: Motor Control, Imagery, and Perception. Behav. Brain Sci. 2004, 27, 377–396. [Google Scholar] [CrossRef]
- Lorey, B.; Naumann, T.; Pilgramm, S.; Petermann, C.; Bischoff, M.; Zentgraf, K.; Stark, R.; Vaitl, D.; Munzert, J. How Equivalent Are the Action Execution, Imagery, and Observation of Intransitive Movements? Revisiting the Concept of Somatotopy during Action Simulation. Brain Cogn. 2013, 81, 139–150. [Google Scholar] [CrossRef]
- Barsalou, L.W. Perceptual Symbol Systems. Behav. Brain Sci. 1999, 22, 577–660. [Google Scholar] [CrossRef]
- Barsalou, L.W. Grounded Cognition: Past, Present, and Future. Top. Cogn. Sci. 2010, 2, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Lorey, B.; Bischoff, M.; Pilgramm, S.; Stark, R.; Munzert, J.; Zentgraf, K. The Embodied Nature of Motor Imagery: The Influence of Posture and Perspective. Exp. Brain Res. 2009, 194, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Gallese, V. Embodied Simulation: From Neurons to Phenomenal Experience. Phenom. Cogn. Sci. 2005, 4, 23–48. [Google Scholar] [CrossRef]
- Wolpert, D.M.; Diedrichsen, J.; Flanagan, J.R. Principles of Sensorimotor Learning. Nat. Rev. Neurosci. 2011, 12, 739–751. [Google Scholar] [CrossRef]
- Bart, V.K.E.; Koch, I.; Rieger, M. Inhibitory Mechanisms in Motor Imagery: Disentangling Different Forms of Inhibition Using Action Mode Switching. Psychol. Res. 2021, 85, 1418–1438. [Google Scholar] [CrossRef]
- Rieger, M.; Dahm, S.F.; Koch, I. Inhibition in Motor Imagery: A Novel Action Mode Switching Paradigm. Psychon. Bull. Rev. 2017, 24, 459–466. [Google Scholar] [CrossRef]
- Steggemann, Y.; Engbert, K.; Weigelt, M. Selective Effects of Motor Expertise in Mental Body Rotation Tasks: Comparing Object-Based and Perspective Transformations. Brain Cogn. 2011, 76, 97–105. [Google Scholar] [CrossRef]
- Hyun, J.-S.; Luck, S.J. Visual Working Memory as the Substrate for Mental Rotation. Psychon. Bull. Rev. 2007, 14, 154–158. [Google Scholar] [CrossRef]
- Seepanomwan, K.; Caligiore, D.; Cangelosi, A.; Baldassarre, G. Generalisation, Decision Making, and Embodiment Effects in Mental Rotation: A Neurorobotic Architecture Tested with a Humanoid Robot. Neural Netw. 2015, 72, 31–47. [Google Scholar] [CrossRef]
- Xue, J.; Li, C.; Quan, C.; Lu, Y.; Yue, J.; Zhang, C. Uncovering the Cognitive Processes Underlying Mental Rotation: An Eye-Movement Study. Sci. Rep. 2017, 7, 10076. [Google Scholar] [CrossRef]
- Parsons, L.M. Imagined Spatial Transformation of One’s Body. J. Exp. Psychol. Gen. 1987, 116, 172–191. [Google Scholar] [CrossRef] [PubMed]
- Barhoun, P.; Fuelscher, I.; Do, M.; He, J.L.; Bekkali, S.; Cerins, A.; Youssef, G.J.; Williams, J.; Enticott, P.G.; Hyde, C. Mental Rotation Performance in Young Adults with and without Developmental Coordination Disorder. Hum. Mov. Sci. 2021, 77, 102787. [Google Scholar] [CrossRef] [PubMed]
- Voyer, D.; Jansen, P. Motor Expertise and Performance in Spatial Tasks: A Meta-Analysis. Hum. Mov. Sci. 2017, 54, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Zacks, J.M.; Mires, J.; Tversky, B.; Hazeltine, E. Mental Spatial Transformations of Objects and Perspective. Spat. Cogn. Comput. 2000, 2, 315–332. [Google Scholar] [CrossRef]
- Böffel, C.; Müsseler, J. Perceived Ownership of Avatars Influences Visual Perspective Taking. Front. Psychol. 2018, 9, 743. [Google Scholar] [CrossRef]
- Zacks, J.M.; Michelon, P. Transformations of Visuospatial Images. Behav. Cogn. Neurosci. Rev. 2005, 4, 96–118. [Google Scholar] [CrossRef]
- Vingerhoets, G.; de Lange, F.P.; Vandemaele, P.; Deblaere, K.; Achten, E. Motor Imagery in Mental Rotation: An FMRI Study. NeuroImage 2002, 17, 1623–1633. [Google Scholar] [CrossRef]
- Tomasino, B.; Rumiati, R.I.; Umiltà, C.A. Selective Deficit of Motor Imagery as Tapped by a Left-Right Decision of Visually Presented Hands. Brain Cogn. 2003, 53, 376–380. [Google Scholar] [CrossRef]
- Feng, T.; Zhang, Z.; Ji, Z.; Jia, B.; Li, Y. Selective Effects of Sport Expertise on the Stages of Mental Rotation Tasks with Object-Based and Egocentric Transformations. Adv. Cogn. Psychol. 2017, 13, 248–256. [Google Scholar] [CrossRef]
- Habacha, H.; Lejeune-Poutrain, L.; Molinaro, C. Realistic Stimuli Reveal Selective Effects of Motor Expertise during a Mental Body Rotation Task. Am. J. Psychol. 2017, 130, 47–62. [Google Scholar] [CrossRef]
- Jost, L.; Jansen, P. Manual Training of Mental Rotation Performance: Visual Representation of Rotating Figures Is the Main Driver for Improvements. Q. J. Exp. Psychol. 2022, 75, 695–711. [Google Scholar] [CrossRef] [PubMed]
- Kaltner, S.; Riecke, B.E.; Jansen, P. Embodied Mental Rotation: A Special Link between Egocentric Transformation and the Bodily Self. Front. Psychol. 2014, 5, 505. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, L.; Lenggenhager, B.; Blanke, O. The Timing of Temporoparietal and Frontal Activations during Mental Own Body Transformations from Different Visuospatial Perspectives. Hum. Brain Mapp. 2009, 30, 1801–1812. [Google Scholar] [CrossRef]
- Barhoun, P.; Fuelscher, I.; Kothe, E.J.; He, J.L.; Youssef, G.J.; Enticott, P.G.; Williams, J.; Hyde, C. Motor Imagery in Children with DCD: A Systematic and Meta-Analytic Review of Hand-Rotation Task Performance. Neurosci. Biobehav. Rev. 2019, 99, 282–297. [Google Scholar] [CrossRef]
- Vandierendonck, A. A Comparison of Methods to Combine Speed and Accuracy Measures of Performance: A Rejoinder on the Binning Procedure. Behav. Res. Methods 2017, 49, 653–673. [Google Scholar] [CrossRef]
- Vandierendonck, A. On the Utility of Integrated Speed-Accuracy Measures When Speed-Accuracy Trade-off Is Present. J. Cogn. 2021, 4, 26. [Google Scholar] [CrossRef]
- Vannuscorps, G.; Pillon, A.; Andres, M. Effect of Biomechanical Constraints in the Hand Laterality Judgment Task: Where Does It Come From? Front. Hum. Neurosci. 2012, 6, 299. [Google Scholar] [CrossRef] [PubMed]
- van Elk, M.; Blanke, O. Imagined Own-Body Transformations during Passive Self-Motion. Psychol. Res. 2014, 78, 18–27. [Google Scholar] [CrossRef]
- Peters, M. Neuropsychological Identification of Motor Problems: Can We Learn Something from the Feet and Legs That Hands and Arms Will Not Tell Us? Neuropsychol. Rev. 1990, 1, 165–183. [Google Scholar] [CrossRef]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- The Belmont Report: Ethical Principles and Guidelines for the Protection of Human Subjects of Research; DHEW Publication; The Commission for Sale by the Supt. of Docs., U.S. Government Printing Office: Bethesda, MD, USA; Washington, DC, USA, 1978.
- Muraki, E.J.; Dahm, S.F.; Pexman, P.M. Meaning in Hand: Investigating Shared Mechanisms of Motor Imagery and Sensorimotor Simulation in Language Processing. PsyarXiv 2022, preprint. [Google Scholar]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Peirce, J.; Gray, J.R.; Simpson, S.; MacAskill, M.; Höchenberger, R.; Sogo, H.; Kastman, E.; Lindeløv, J.K. PsychoPy2: Experiments in Behavior Made Easy. Behav. Res. Methods 2019, 51, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Bridges, D.; Pitiot, A.; MacAskill, M.R.; Peirce, J.W. The Timing Mega-Study: Comparing a Range of Experiment Generators, Both Lab-Based and Online. PeerJ 2020, 8, e9414. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. Rstatix, 2021. Available online: https://rpkgs.datanovia.com/rstatix/ (accessed on 5 January 2022).
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio Team: Boston, MA, USA, 2022. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; ISBN 9783319242750. [Google Scholar]
- Schwarzer, R. Optimistische Kompetenzerwartung: Zur Erfassung Einer Personellen Bewältigungsressource. Diagnostica 1994, 40, 105–123. [Google Scholar]
- Dahm, S.F.; Bart, V.K.E.; Pithan, J.M.; Rieger, M. Deutsche Übersetzung und Validierung des VMIQ-2 zur Erfassung der Lebhaftigkeit von Handlungsvorstellungen. Z. Für Sportpsychol. 2019, 26, 151–158. [Google Scholar] [CrossRef]
- Dahm, S.F. Validation of a Computer-Based Version of the Vividness of Movement Imagery Questionnaire. PTAD 2022, 1–13. [Google Scholar] [CrossRef]
- Serdar, C.C.; Cihan, M.; Yücel, D.; Serdar, M.A. Sample Size, Power and Effect Size Revisited: Simplified and Practical Approaches in Pre-Clinical, Clinical and Laboratory Studies. Biochem. Med. 2021, 31, 010502. [Google Scholar] [CrossRef]
- Blender Online Community. Blender—A 3D Modelling and Rendering Package; Stichting Blender Foundation: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Mathôt, S.; March, J. Conducting Linguistic Experiments Online with OpenSesame and OSWeb. Lang. Learn 2022, 1–32. [Google Scholar] [CrossRef]
- Lange, K.; Kühn, S.; Filevich, E. “Just Another Tool for Online Studies” (JATOS): An Easy Solution for Setup and Management of Web Servers Supporting Online Studies. PLoS ONE 2015, 10, e0130834. [Google Scholar] [CrossRef]
- Burton, L.J.; Fogarty, G.J. The Factor Structure of Visual Imagery and Spatial Abilities. Intelligence 2003, 31, 289–318. [Google Scholar] [CrossRef]
- Shepard, R.N.; Cooper, L.A. Mental Images and Their Transformations; The MIT Press: Cambridge, MA, USA, 1986; p. 364. ISBN 9780262192002. [Google Scholar]
- Marks, D.F. Visual Imagery Differences in the Recall of Pictures. Br. J. Psychol. 1973, 64, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.E.; Guillot, A.; Di Rienzo, F.; Cumming, J. Comparing Self-Report and Mental Chronometry Measures of Motor Imagery Ability. Eur. J. Sport Sci. 2015, 15, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Zell, E.; Strickhouser, J.E.; Sedikides, C.; Alicke, M.D. The Better-than-Average Effect in Comparative Self-Evaluation: A Comprehensive Review and Meta-Analysis. Psychol. Bull. 2020, 146, 118–149. [Google Scholar] [CrossRef]
- Dahm, S.F.; Rieger, M. Cognitive Constraints on Motor Imagery. Psychol. Res. 2016, 80, 235–247. [Google Scholar] [CrossRef]
- Pietsch, S.; Jansen, P. Different Mental Rotation Performance in Students of Music, Sport and Education. Learn. Individ. Differ. 2012, 22, 159–163. [Google Scholar] [CrossRef]
- McInnes, K.; Friesen, C.; Boe, S. Specific Brain Lesions Impair Explicit Motor Imagery Ability: A Systematic Review of the Evidence. Arch. Phys. Med. Rehabil. 2016, 97, 478–489. [Google Scholar] [CrossRef]
- Boe, S.; Gionfriddo, A.; Kraeutner, S.; Tremblay, A.; Little, G.; Bardouille, T. Laterality of Brain Activity during Motor Imagery Is Modulated by the Provision of Source Level Neurofeedback. NeuroImage 2014, 101, 159–167. [Google Scholar] [CrossRef]
- Guillot, A.; Debarnot, U.; Louis, M.; Hoyek, N.; Collet, C. Motor Imagery and Motor Performance: Evidence from the Sport Science Literature; Oxford University Press: Oxford, UK, 2010; ISBN 9780191701412. [Google Scholar]
- Simonsmeier, B.A.; Androniea, M.; Buecker, S.; Frank, C. The Effects of Imagery Interventions in Sports: A Meta-Analysis. Int. J. Sport Exerc. Psychol. 2021, 14, 186–207. [Google Scholar] [CrossRef]
- Durand, E.; Masson-Trottier, M.; Sontheimer, A.; Ansaldo, A.I. Increased Links between Language and Motor Areas: A Proof-of-Concept Study on Resting-State Functional Connectivity Following Personalized Observation, Execution and Mental Imagery Therapy in Chronic Aphasia. Brain Cogn. 2021, 148, 105659. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, N.F.; De Buglio, M.; Trimarchi, P.D.; Chielli, A.; Bricolo, E. Mental Practice Promotes Motor Anticipation: Evidence from Skilled Music Performance. Front. Hum. Neurosci. 2013, 7, 451. [Google Scholar] [CrossRef] [PubMed]
- Isaac, A.R.; Marks, D.F.; Russell, D.G. An Instrument for Assessing Imagery of Movement: The Vividness of Movement Imagery Questionnaire (VMIQ). J. Ment. Imag. 1986, 10, 23–30. [Google Scholar]
- Milton, J.; Small, S.L.; Solodkin, A. Imaging Motor Imagery: Methodological Issues Related to Expertise. Methods 2008, 45, 336–341. [Google Scholar] [CrossRef]
- Adams, I.L.J.; Lust, J.M.; Wilson, P.H.; Steenbergen, B. Development of Motor Imagery and Anticipatory Action Planning in Children with Developmental Coordination Disorder—A Longitudinal Approach. Hum. Mov. Sci. 2017, 55, 296–306. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F | p | ηp² | |
|---|---|---|---|
| Abstractness | 5.2 | 0.025 | 0.41 |
| Perspective | 29.4 | <0.001 | 0.20 |
| Rotation | 137.2 | <0.001 | 0.53 |
| Limb | 93.0 | <0.001 | 0.44 |
| Abstractness × Perspective | 42.0 | <0.001 | 0.26 |
| Abstractness × Rotation | 0.1 | 0.803 | <0.01 |
| Abstractness × Limb | 30.6 | <0.001 | 0.20 |
| Perspective × Rotation | 67.3 | <0.001 | 0.36 |
| Perspective × Limb | 0.2 | 0.643 | <0.01 |
| Rotation × Limb | 2.9 | 0.094 | 0.02 |
| Abstractness × Perspective × Rotation | 0.3 | 0.565 | <0.01 |
| Abstractness × Perspective × Limb | 5.8 | 0.018 | 0.05 |
| Abstractness × Rotation × Limb | 0.3 | 0.564 | <0.01 |
| Perspective × Rotation × Limb | 22.3 | <0.001 | 0.16 |
| Abstractness × Perspective × Rotation × Limb | 3.5 | 0.065 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dahm, S.F.; Muraki, E.J.; Pexman, P.M. Hand and Foot Selection in Mental Body Rotations Involves Motor-Cognitive Interactions. Brain Sci. 2022, 12, 1500. https://doi.org/10.3390/brainsci12111500
Dahm SF, Muraki EJ, Pexman PM. Hand and Foot Selection in Mental Body Rotations Involves Motor-Cognitive Interactions. Brain Sciences. 2022; 12(11):1500. https://doi.org/10.3390/brainsci12111500
Chicago/Turabian StyleDahm, Stephan Frederic, Emiko J. Muraki, and Penny M. Pexman. 2022. "Hand and Foot Selection in Mental Body Rotations Involves Motor-Cognitive Interactions" Brain Sciences 12, no. 11: 1500. https://doi.org/10.3390/brainsci12111500
APA StyleDahm, S. F., Muraki, E. J., & Pexman, P. M. (2022). Hand and Foot Selection in Mental Body Rotations Involves Motor-Cognitive Interactions. Brain Sciences, 12(11), 1500. https://doi.org/10.3390/brainsci12111500