
The PACAP/PAC1 Receptor System and Feeding



Abstract
1. Introduction
2. PACAP, Its Receptors, and Its Physiological Roles
3. PACAP and Feeding
3.1. The Role of Endogenous PACAP in Food Intake
3.2. The Effect of Exogenous PACAP and Related Ligands on Food Intake
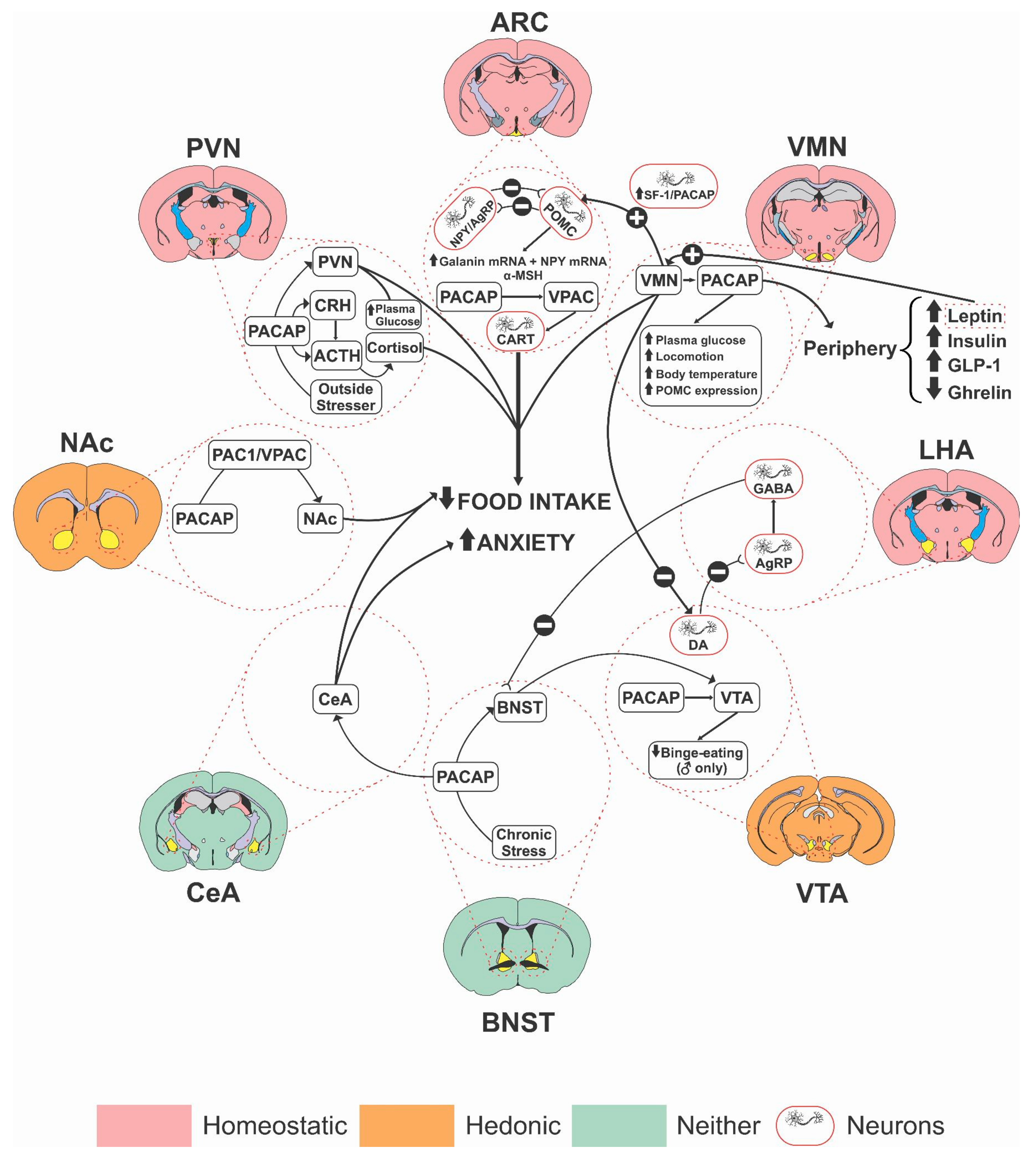
4. Neuroanatomical Sites of PACAP Action in Regulating Food Intake
5. The Role of Other Neurotransmitters in the Actions of PACAP on Food Intake
6. The Role of PACAP Receptor(s) in the Regulatory Actions of PACAP on Feeding
7. Signaling Involved in the Modulatory Actions of PACAP on Food Intake
8. The Action of PACAP on Homeostatic and Hedonic Aspects of Feeding
9. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roh, E.; Song, D.K.; Kim, M.S. Emerging role of the brain in the homeostatic regulation of energy and glucose metabolism. Exp. Mol. Med. 2016, 48, e216. [Google Scholar] [CrossRef]
- Valassi, E.; Scacchi, M.; Cavagnini, F. Neuroendocrine control of food intake. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 158–168. [Google Scholar] [CrossRef]
- Kohno, D.; Gao, H.Z.; Muroya, S.; Kikuyama, S.; Yada, T. Ghrelin directly interacts with neuropeptide-Y-containing neurons in the rat arcuate nucleus: Ca2+ signaling via protein kinase A and N-type channel-dependent mechanisms and cross-talk with leptin and orexin. Diabetes 2003, 52, 948–956. [Google Scholar] [CrossRef]
- Tachibana, T.; Saito, S.; Tomonaga, S.; Takagi, T.; Saito, E.S.; Boswell, T.; Furuse, M. Intracerebroventricular injection of vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide inhibits feeding in chicks. Neurosci. Lett. 2003, 339, 203–206. [Google Scholar] [CrossRef]
- Hirabayashi, T.; Nakamachi, T.; Shioda, S. Discovery of PACAP and its receptors in the brain. J. Headache Pain 2018, 19, 1–8. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Kambe, Y.; Kurihara, T.; Nakamachi, T.; Shintani, N.; Hashimoto, H.; Miyata, A. Pituitary Adenylate Cyclase-Activating Polypeptide in the Ventromedial Hypothalamus Is Responsible for Food Intake Behavior by Modulating the Expression of Agouti-Related Peptide in Mice. Mol. Neurobiol. 2020, 57, 2101–2114. [Google Scholar] [CrossRef]
- Nakamachi, T.; Tanigawa, A.; Konno, N.; Shioda, S.; Matsuda, K. Expression Patterns of PACAP and PAC1R Genes and Anorexigenic Action of PACAP1 and PACAP2 in Zebrafish. Front. Endocrinol. 2019, 10, 227. [Google Scholar] [CrossRef]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- May, V.; Buttolph, T.R.; Girard, B.M.; Clason, T.A.; Parsons, R.L. PACAP-induced ERK activation in HEK cells expressing PAC1 receptors involves both receptor internalization and PKC signaling. Am. J. Physiol. Physiol. 2014, 306, C1068–C1079. [Google Scholar] [CrossRef]
- Merriam, L.A.; Baran, C.N.; Girard, B.M.; Hardwick, J.C.; May, V.; Parsons, R.L. Pituitary adenylate cyclase 1 receptor internalization and endosomal signaling mediate the pituitary adenylate cyclase activating polypeptide-induced increase in guinea pig cardiac neuron excitability. J. Neurosci. 2013, 33, 4614–4622. [Google Scholar] [CrossRef]
- Morley, J.E.; Horowitz, M.; Morley, P.M.; Flood, J.F. Pituitary adenylate cyclase activating polypeptide (PACAP) reduces food intake in mice. Peptides 1992, 13, 1133–1135. [Google Scholar] [CrossRef]
- Mizuno, Y.; Kondo, K.; Terashima, Y.; Arima, H.; Murase, T.; Oiso, Y. Anorectic effect of pituitary adenylate cyclase activating polypeptide (PACAP) in rats: Lack of evidence for involvement of hypothalamic neuropeptide gene expression. J. Neuroendocrinol. 1998, 10, 611–616. [Google Scholar] [CrossRef]
- Matsuda, K.; Maruyama, K.; Nakamachi, T.; Miura, T.; Shioda, S. Effects of pituitary adenylate cyclase-activating polypeptide and vasoactive intestinal polypeptide on food intake and locomotor activity in the goldfish, Carassius auratus. Ann. N. Y. Acad. Sci. 2006, 1070, 417–421. [Google Scholar] [CrossRef]
- Matsuda, K.; Maruyama, K.; Nakamachi, T.; Miura, T.; Uchiyama, M.; Shioda, S. Inhibitory effects of pituitary adenylate cyclase-activating polypeptide (PACAP) and vasoactive intestinal peptide (VIP) on food intake in the goldfish, Carassius auratus. Peptides 2005, 26, 1611–1616. [Google Scholar] [CrossRef]
- Vu, J.P.; Goyal, D.; Luong, L.; Oh, S.; Sandhu, R.; Norris, J.; Parsons, W.; Pisegna, J.R.; Germano, P.M. PACAP intraperitoneal treatment suppresses appetite and food intake via PAC1 receptor in mice by inhibiting ghrelin and increasing GLP-1 and leptin. Am. J. Physiol. Liver Physiol. 2015, 309, G816–G825. [Google Scholar] [CrossRef]
- Nakata, M.; Kohno, D.; Shintani, N.; Nemoto, Y.; Hashimoto, H.; Baba, A.; Yada, T. PACAP deficient mice display reduced carbohydrate intake and PACAP activates NPY-containing neurons in the rat hypothalamic arcuate nucleus. Neurosci. Lett. 2004, 370, 252–256. [Google Scholar] [CrossRef]
- Jozsa, R.; Nemeth, J.; Tamas, A.; Hollosy, T.; Lubics, A.; Jakab, B.; Olah, A.; Lengvari, I.; Arimura, A.; Reglödi, D. Short-term fasting differentially alters PACAP and VIP levels in the brains of rat and chicken. Ann. N. Y. Acad. Sci. 2006, 1070, 354–358. [Google Scholar] [CrossRef]
- Kiss, P.; Reglodi, D.; Tamás, A.; Lubics, A.; Lengvári, I.; Józsa, R.; Somogyvári-Vigh, A.; Szilvássy, Z.; Németh, J. Changes of PACAP levels in the brain show gender differences following short-term water and food deprivation. Gen. Comp. Endocrinol. 2007, 152, 225–230. [Google Scholar] [CrossRef]
- Agarwal, A.; Halvorson, L.M.; Legradi, G. Pituitary adenylate cyclase-activating polypeptide (PACAP) mimics neuroendocrine and behavioral manifestations of stress: Evidence for PKA-mediated expression of the corticotropin-releasing hormone (CRH) gene. Mol. Brain Res. 2005, 138, 45–57. [Google Scholar] [CrossRef]
- Hammack, S.E.; Cheung, J.; Rhodes, K.M.; Schutz, K.C.; Falls, W.A.; Braas, K.M.; May, V. Chronic stress increases pituitary adenylate cyclase-activating peptide (PACAP) and brain-derived neurotrophic factor (BDNF) mRNA expression in the bed nucleus of the stria terminalis (BNST): Roles for PACAP in anxiety-like behavior. Psychoneuroendocrinology 2009, 34, 833–843. [Google Scholar] [CrossRef]
- Lehmann, M.L.; Mustafa, T.; Eiden, A.M.; Herkenham, M.; Eiden, L.E. PACAP-deficient mice show attenuated corticosterone secretion and fail to develop depressive behavior during chronic social defeat stress. Psychoneuroendocrinology 2013, 38, 702–715. [Google Scholar] [CrossRef]
- Stroth, N.; Eiden, L.E. Stress hormone synthesis in mouse hypothalamus and adrenal gland triggered by restraint is dependent on pituitary adenylate cyclase-activating polypeptide signaling. Neuroscience 2010, 165, 1025–1030. [Google Scholar] [CrossRef]
- Stroth, N.; Holighaus, Y.; Ait-Ali, D.; Eiden, L.E. PACAP: A master regulator of neuroendocrine stress circuits and the cellular stress response. Ann. N. Y. Acad. Sci. 2011, 1220, 49–59. [Google Scholar] [CrossRef]
- Stroth, N.; Liu, Y.; Aguilera, G.; Eiden, L.E. Pituitary adenylate cyclase-activating polypeptide controls stimulus-transcription coupling in the hypothalamic-pituitary-adrenal axis to mediate sustained hormone secretion during stress. J. Neuroendocr. 2011, 23, 944–955. [Google Scholar] [CrossRef]
- Tsukiyama, N.; Saida, Y.; Kakuda, M.; Shintani, N.; Hayata, A.; Morita, Y.; Tanida, M.; Tajiri, M.; Hazama, K.; Ogata, K.; et al. PACAP centrally mediates emotional stress-induced corticosterone responses in mice. Stress 2011, 14, 368–375. [Google Scholar] [CrossRef]
- Hawke, Z.; Ivanov, T.R.; Bechtold, D.A.; Dhillon, H.; Lowell, B.B.; Luckman, S.M. PACAP neurons in the hypothalamic ventromedial nucleus are targets of central leptin signaling. J. Neurosci. 2009, 29, 14828–14835. [Google Scholar] [CrossRef]
- Gargiulo, A.T.; Curtis, G.R.; Barson, J.R. Pleiotropic pituitary adenylate cyclase-activating polypeptide (PACAP): Novel insights into the role of PACAP in eating and drug intake. Brain Res. 2020, 1729, 146626. [Google Scholar] [CrossRef]
- Mounien, L.; Bizet, P.; Boutelet, I.; Gourcerol, G.; Basille, M.; Gonzalez, B.; Vaudry, H.; Jegou, S. Expression of PACAP receptor mRNAs by neuropeptide Y neurons in the rat arcuate nucleus. Ann. N. Y. Acad. Sci. 2006, 1070, 457–461. [Google Scholar] [CrossRef]
- Sherwood, N.M.; Adams, B.A.; Isaac, E.R.; Wu, S.; Fradinger, E.A. Knocked down and out: PACAP in development, reproduction and feeding. Peptides 2007, 28, 1680–1687. [Google Scholar] [CrossRef]
- Gray, S.L.; Cummings, K.J.; Jirik, F.R.; Sherwood, N.M. Targeted disruption of the pituitary adenylate cyclase-activating polypeptide gene results in early postnatal death associated with dysfunction of lipid and carbohydrate metabolism. Mol. Endocrinol. 2001, 15, 1739–1747. [Google Scholar] [CrossRef]
- Green, B.D.; Irwin, N.; Cassidy, R.S.; Gault, V.A.; Flatt, P.R. Long-term administration of PACAP receptor antagonist, PACAP(6-27), impairs glucose tolerance and insulin sensitivity in obese diabetic ob/ob mice. Peptides 2006, 27, 2343–2349. [Google Scholar] [CrossRef]
- Hurley, M.M.; Maunze, B.; Block, M.E.; Frenkel, M.M.; Reilly, M.J.; Kim, E.; Chen, Y.; Li, Y.; Baker, D.A.; Liu, Q.S.; et al. Pituitary Adenylate-Cyclase Activating Polypeptide Regulates Hunger- and Palatability-Induced Binge Eating. Front. Neurosci. 2016, 10, 383. [Google Scholar] [CrossRef]
- Bresson-Bépoldin, L.; Jacquot, M.C.; Schlegel, W.; Rawlings, S.R. Multiple splice variants of the pituitary adenylate cyclase-activating polypeptide type 1 receptor detected by RT-PCR in single rat pituitary cells. J. Mol. Endocrinol. 1998, 21, 109–120. [Google Scholar] [CrossRef]
- Burgos, J.R.; Iresjo, B.M.; Smedh, U. Pituitary adenylate cyclase-activating polypeptide 6-38 blocks cocaine- and amphetamine-regulated transcript Peptide-induced hypophagia in rats. PLoS ONE 2013, 8, e72347. [Google Scholar] [CrossRef] [PubMed]
- Hannibal, J.; Georg, B.; Fahrenkrug, J. Altered Circadian Food Anticipatory Activity Rhythms in PACAP Receptor 1 (PAC1) Deficient Mice. PLoS ONE 2016, 11, e0146981. [Google Scholar] [CrossRef] [PubMed]
- Marquez, P.; Bebawy, D.; Lelievre, V.; Coute, A.C.; Evans, C.J.; Waschek, J.A.; Lutfy, K. The role of endogenous PACAP in motor stimulation and conditioned place preference induced by morphine in mice. Psychopharmacology 2009, 204, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Mounien, L.; Rego, J.C.D.; Bizet, P.; Boutelet, I.; Gourcerol, G.; Fournier, A.; Brabet, P.; Costentin, J.; Vaudry, H.; Jégou, S. Pituitary adenylate cyclase-activating polypeptide inhibits food intake in mice through activation of the hypothalamic melanocortin system. Neuropsychopharmacology 2009, 34, 424–435. [Google Scholar] [CrossRef]
- Gonzalez, B.J.; Basille, M.; Vaudry, D.; Fournier, A.; Vaudry, H. Pituitary adenylate cyclase-activating polypeptide promotes cell survival and neurite outgrowth in rat cerebellar neuroblasts. Neuroscience 1997, 78, 419–430. [Google Scholar] [CrossRef]
- Hezareh, M.; Schlegel, W.; Rawlings, S.R. PACAP and VIP stimulate Ca2+ oscillations in rat gonadotrophs through the PACAP/VIP type 1 receptor (PVR1) linked to a pertussis toxin-insensitive G-protein and the activation of phospholipase C-beta. J. Neuroendocrinol. 1996, 8, 367–374. [Google Scholar] [CrossRef]
- Yada, T.; Sakurada, M.; Filipsson, K.; Kikuchi, M.; Ahrén, B. Intraperitoneal PACAP administration decreases blood glucose in GK rats, and in normal and high fat diet mice. Ann. N. Y. Acad. Sci. 2000, 921, 259–263. [Google Scholar] [CrossRef]
- Yu, R.; Yi, T.; Xie, S.; Hong, A. Long-term administration of maxadilan improves glucose tolerance and insulin sensitivity in mice. Peptides 2008, 29, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Yada, T.; Sakurada, M.; Ihida, K.; Nakata, M.; Murata, F.; Arimura, A.; Kikuchi, M. Pituitary adenylate cyclase activating polypeptide is an extraordinarily potent intra-pancreatic regulator of insulin secretion from islet beta-cells. J. Biol. Chem. 1994, 269, 1290–1293. [Google Scholar] [CrossRef]
- Yokota, C.; Kawai, K.; Ohashi, S.; Watanabe, Y.; Yamashita, K. PACAP stimulates glucose output from the perfused rat liver. Peptides 1995, 16, 55–60. [Google Scholar] [CrossRef]
- Lugo, J.M.; Oliva, A.; Morales, A.; Reyes, O.; Garay, H.E.; Herrera, F.; Cabrales, A.; Pérez, E.; Estrada, M.P. The biological role of pituitary adenylate cyclase-activating polypeptide (PACAP) in growth and feeding behavior in juvenile fish. J. Pept. Sci. 2010, 16, 633–643. [Google Scholar] [CrossRef]
- Choi, D.C.; Evanson, N.K.; Furay, A.R.; Ulrich-Lai, Y.M.; Ostrander, M.M.; Herman, J.P. The anteroventral bed nucleus of the stria terminalis differentially regulates hypothalamic-pituitary-adrenocortical axis responses to acute and chronic stress. Endocrinology 2008, 149, 818–826. [Google Scholar] [CrossRef]
- Roman, C.W.; Lezak, K.R.; Kocho-Schellenberg, M.; Garret, M.A.; Braas, K.; May, V.; Hammack, S.E. Excitotoxic lesions of the bed nucleus of the stria terminalis (BNST) attenuate the effects of repeated stress on weight gain: Evidence for the recruitment of BNST activity by repeated, but not acute, stress. Behav. Brain Res. 2012, 227, 300–304. [Google Scholar] [CrossRef]
- Kocho-Schellenberg, M.; Lezak, K.R.; Harris, O.M.; Roelke, E.; Gick, N.; Choi, I.; Edwards, S.; Wasserman, E.; Toufexis, D.J.; Braas, K.M.; et al. PACAP in the BNST produces anorexia and weight loss in male and female rats. Neuropsychopharmacology 2014, 39, 1614–1623. [Google Scholar] [CrossRef]
- Resch, J.M.; Boisvert, J.P.; Hourigan, A.E.; Mueller, C.R.; Yi, S.S.; Choi, S. Stimulation of the hypothalamic ventromedial nuclei by pituitary adenylate cyclase-activating polypeptide induces hypophagia and thermogenesis. Am. J. Physiol. Integr. Comp. Physiol. 2011, 301, R1625–R1634. [Google Scholar] [CrossRef]
- Resch, J.M.; Maunze, B.; Gerhardt, A.K.; Magnuson, S.K.; Phillips, K.A.; Choi, S. Intrahypothalamic pituitary adenylate cyclase-activating polypeptide regulates energy balance via site-specific actions on feeding and metabolism. Am. J. Physiol. Metab. 2013, 305, E1452–E1463. [Google Scholar] [CrossRef]
- Iemolo, A.; Ferragud, A.; Cottone, P.; Sabino, V. Pituitary Adenylate Cyclase-Activating Peptide in the Central Amygdala Causes Anorexia and Body Weight Loss via the Melanocortin and the TrkB Systems. Neuropsychopharmacology 2015, 40, 1846–1855. [Google Scholar] [CrossRef]
- Ferragud, A.; Velazquez-Sanchez, C.; Minnig, M.A.; Sabino, V.; Cottone, P. Pituitary adenylate cyclase-activating polypeptide (PACAP) modulates dependence-induced alcohol drinking and anxiety-like behavior in male rats. Neuropsychopharmacology 2021, 46, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Gargiulo, A.T.; Pirino, B.E.; Curtis, G.R.; Barson, J.R. Effects of pituitary adenylate cyclase-activating polypeptide isoforms in nucleus accumbens subregions on ethanol drinking. Addict. Biol. 2021, 26, e12972. [Google Scholar] [CrossRef]
- Gupta, A.; Gargiulo, A.T.; Curtis, G.R.; Badve, P.S.; Pandey, S.; Barson, J.R. Pituitary Adenylate Cyclase-Activating Polypeptide-27 (PACAP-27) in the Thalamic Paraventricular Nucleus Is Stimulated by Ethanol Drinking. Alcohol. Clin. Exp. Res. 2018, 42, 1650–1660. [Google Scholar] [CrossRef] [PubMed]
- Chance, W.T.; Thompson, H.; Thomas, I.; Fischer, J.E. Anorectic and neurochemical effects of pituitary adenylate cyclase activating polypeptide in rats. Peptides 1995, 16, 1511–1516. [Google Scholar] [CrossRef]
- Shioda, S.; Shuto, Y.; Somogyvari-Vigh, A.; Legradi, G.; Onda, H.; Coy, D.H.; Nakajo, S.; Arimura, A. Localization and gene expression of the receptor for pituitary adenylate cyclase-activating polypeptide in the rat brain. Neurosci. Res. 1997, 28, 345–354. [Google Scholar] [CrossRef]
- Baker, D.A.; Xi, Z.X.; Shen, H.; Swanson, C.J.; Kalivas, P.W. The origin and neuronal function of in vivo nonsynaptic glutamate. J. Neurosci. 2002, 22, 9134–9141. [Google Scholar] [CrossRef]
- Resch, J.M.; Maunze, B.; Phillips, K.A.; Choi, S. Inhibition of food intake by PACAP in the hypothalamic ventromedial nuclei is mediated by NMDA receptors. Physiol. Behav. 2014, 133, 230–235. [Google Scholar] [CrossRef]
- Adams, B.A.; Gray, S.L.; Isaac, E.R.; Bianco, A.C.; Vidal-Puig, A.J.; Sherwood, N.M. Feeding and metabolism in mice lacking pituitary adenylate cyclase-activating polypeptide. Endocrinology 2008, 149, 1571–1580. [Google Scholar] [CrossRef]
- Ohgushi, A.; Bungo, T.; Shimojo, M.; Masuda, Y.; Denbow, D.M.; Furuse, M. Relationships between feeding and locomotion behaviors after central administration of CRF in chicks. Physiol. Behav. 2001, 72, 287–289. [Google Scholar] [CrossRef]
- Bernier, N.J.; Bedard, N.; Peter, R.E. Effects of cortisol on food intake, growth, and forebrain neuropeptide Y and corticotropin-releasing factor gene expression in goldfish. Gen. Comp. Endocrinol. 2004, 135, 230–240. [Google Scholar] [CrossRef]
- Missig, G.; Roman, C.W.; Vizzard, M.A.; Braas, K.M.; Hammack, S.E.; May, V. Parabrachial nucleus (PBn) pituitary adenylate cyclase activating polypeptide (PACAP) signaling in the amygdala: Implication for the sensory and behavioral effects of pain. Neuropharmacology 2014, 86, 38–48. [Google Scholar] [CrossRef]
- Lezak, K.R.; Roelke, E.; Harris, O.M.; Choi, I.; Edwards, S.; Gick, N.; Cocchiaro, G.; Missig, G.; Roman, C.W.; Braas, K.M.; et al. Pituitary adenylate cyclase-activating polypeptide (PACAP) in the bed nucleus of the stria terminalis (BNST) increases corticosterone in male and female rats. Psychoneuroendocrinology 2014, 45, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Miles, O.W.; Thrailkill, E.A.; Linden, A.K.; May, V.; Bouton, M.E.; Hammack, S.E. Pituitary Adenylate Cyclase-Activating Peptide in the Bed Nucleus of the Stria Terminalis Mediates Stress-Induced Reinstatement of Cocaine Seeking in Rats. Neuropsychopharmacology 2018, 43, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, T.; Saito, E.S.; Takahashi, H.; Saito, S.; Tomonaga, S.; Boswell, T.; Furuse, M. Anorexigenic effects of pituitary adenylate cyclase-activating polypeptide and vasoactive intestinal peptide in the chick brain are mediated by corticotrophin-releasing factor. Regul. Pept. 2004, 120, 99–105. [Google Scholar] [CrossRef]
- Chang, R.; Hernandez, J.; Gastelum, C.; Guadagno, K.; Perez, L.; Wagner, E.J. Pituitary Adenylate Cyclase-Activating Polypeptide Excites Proopiomelanocortin Neurons: Implications for the Regulation of Energy Homeostasis. Neuroendocrinology 2021, 111, 45–69. [Google Scholar] [CrossRef] [PubMed]
- Dore, R.; Iemolo, A.; Smith, K.L.; Wang, X.; Cottone, P.; Sabino, V. CRF mediates the anxiogenic and anti-rewarding, but not the anorectic effects of PACAP. Neuropsychopharmacology 2013, 38, 2160–2169. [Google Scholar] [CrossRef]
- Tanida, M.; Mori, M. Nesfatin-1 stimulates renal sympathetic nerve activity in rats. NeuroReport 2011, 22, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Sághy, É.; Payrits, M.; Helyes, Z.; Reglődi, D.; Bánki, E.; Tóth, G.; Couvineau, A.; Szőke, É. Stimulatory effect of pituitary adenylate cyclase-activating polypeptide 6-38, M65 and vasoactive intestinal polypeptide 6-28 on trigeminal sensory neurons. Neuroscience 2015, 308, 144–156. [Google Scholar] [CrossRef]
- Macdonald, D.S.; Weerapura, M.; Beazely, M.A.; Martin, L.; Czerwinski, W.; Roder, J.C.; Orser, B.A.; MacDonald, J.F. Modulation of NMDA receptors by pituitary adenylate cyclase activating peptide in CA1 neurons requires G alpha q, protein kinase C, and activation of Src. J. Neurosci. 2005, 25, 11374–11384. [Google Scholar] [CrossRef]
- Yaka, R.; He, D.Y.; Phamluong, K.; Ron, D. Pituitary adenylate cyclase-activating polypeptide (PACAP(1-38)) enhances N-methyl-D-aspartate receptor function and brain-derived neurotrophic factor expression via RACK1. J. Biol. Chem. 2003, 278, 9630–9638. [Google Scholar] [CrossRef]
- Costa, L.; Santangelo, F.; Volsi, G.L.; Ciranna, L. Modulation of AMPA receptor-mediated ion current by pituitary adenylate cyclase-activating polypeptide (PACAP) in CA1 pyramidal neurons from rat hippocampus. Hippocampus 2009, 19, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Le, N.; Hernandez, J.; Gastelum, C.; Perez, L.; Vahrson, I.; Sayers, S.; Wagner, E.J. Pituitary Adenylate Cyclase Activating Polypeptide Inhibits A(10) Dopamine Neurons and Suppresses the Binge-like Consumption of Palatable Food. Neuroscience 2021, 478, 49–64. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Study By | Subject(s) | Drug Dose Route | Category | Brain Area/Peptide/Peptide Precursors, etc. | Findings |
|---|---|---|---|---|---|
| Introduction | |||||
| Tachibana et al. (2003) | Male Chicks | ICV | Effect of various peptides on food intake | VIP, PACAP, GRF, GLP-1 | VIP, PACAP, GRF, and GLP-1 decreased food intake in chicks, but VIP and PACAP with lesser potency. |
| PACAP and Feeding | |||||
| Matsuda et al. (2006) | N/A | N/A | Effect of PACAP and VIP in feeding | VMN, ARC, PVN, PACAP, VIP, NPY, AgRP, CRH, CART, leptin, orexin | Looks at the role of PACAP and VIP in feeding in vertebrates., as well as the role of orexin, NPY, AgRP, melanin-concentrating hormone, proopiomelanocortin-derived peptides, galanin, galanin-like peptide, ghrelin, CRH, cholecystokinin, CART, and leptin. |
| Matsuda et al. (2005) | Unspecified goldfish | IP VIP, ICV VIP, IP PACAP, ICV PACAP | Effect of PACAP and VIP in feeding | PACAP, VIP | ICV and IP PACAP, and VIP reduced food intake in the goldfish. |
| Mizuno et al. (1998) | Male Sprague–Dawley rats | ICV | PACAP and involvement of hypothalamic neuropeptide gene expression | Hypothalamus | The hypophagic effect of PACAP is not mediated by NPY or CRH. |
| Nakamachi et al. (2019) | Male and female adult short-fin strain zebrafish | ICV | Effect of PACAP on Anorexigenic Action | PACAP1, PACAP2 | The expression levels and distribution of PACAP and PAC1R genes are different in zebrafish, but the hypophagic effect of PACAP is similar to those seen in other vertebrates. |
| Endogenous PACAP | |||||
| Burgos et al. (2013) | Male Sprague–Dawley rats | ICV PACAP, ICV CART | PACAP and CART on Food Intake | PACAP6-38, CART | PACAP6-38 reduced the ability of CART to decrease food intake and body weight (without altering motor activity). |
| Gargiulo et al. (2020) | N/A | N/A | Food and Drug Addiction Alters PACAP respons+es | PACAP27, PACAP38, PAC1 R, VIP, VPAC1, VPAC2 | Review of the literature on the anorectic effects of PACAP and about the dysregulation of PACAP and its receptors following palatable food and addictive drugs. |
| Gray et al. (2001) | Male PAC1R (−/−) and (+/+) mice | IV Glucose (ITT) | Role of PACAP in lipid and carbohydrate metabolism | PACAP, insulin | PACAP regulates carbohydrate and fat metabolism. |
| Jozsa et al. (2006) | Male Wistar rats and broiler chickens | N/A | Measure levels of PACAP and VIP in response to starvation | hypothalamus, telencephalon, brainstem | PACAP levels are increased in response to starvation in both species. |
| Kiss et al. (2007) | Male and female rats | N/A | Role of Endogenous PACAP in food and water deprivation in various brain areas | PACAP38/hypothalamus, brainstem, and telencephalon | PACAP levels were changed in a sexually dimorphic manner in the hypothalamus, brainstem, and telencephalon in mice after food deprivation. |
| Sherwood et al. (2007) | N/A | N/A | Effect of PACAP on reproduction, energy homeostasis, and brain development | PACAP, VPAC, insulin, glucagon | PACAP is crucial for zebrafish brain development and reproduction, but not for body mass or food intake in mice maintained near thermoneutrality. |
| Hannibal et al. (2016) | Male and female PAC1R (−/−) and (+/+) mice | N/A | Role of food in anticipatory activity | PACAP | PAC1 KO mice showed an earlier onset of anticipatory activity than WT at the 10-lux light intensity. Both KO and WT had greater FAA in the SPP 10 and 300 lux, and FPP 300 lux light intensities than FPP 300 intensity. |
| Nakata et al. (2004) | Male PACAP (−/−) & (+/+) and rats | Superfusion | Effect of PACAP on carb intake and NPY in the ARC | PACAP, VIP, NPY, Maxadillian | NPY mRNA was reduced in PACAP-deficient mice. Carbohydrate but not fat intake was reduced in PACAP deficient mice. NPY neuron is not the sole target for PACAP’s actions in the ARC. |
| Exogenous PACAP | |||||
| Green et al. (2006) | Unspecified ob/ob mice | IV PACAP27, IV PACAP6-38, IP PACAP27, IP PACAP6-38, IP Glucose, IP Insulin | Role of endogenous PACAP in glucose and insulin levels in ob/ob mice | PACAP27, PACAP38, PACAP6-38, insulin | PACAP is involved in glucose homeostasis but less in insulin release in ob/ob mice. |
| Lugo et al. (2010) | Unspecified African catfish, Nile tilapia | IP PACAP, IP VIP | Role of GH on PACAP and VIP effects on food intake | PACAP, VIP, GH | PACAP increased food consumption in juvenile tilapia and may be involved in growth. |
| Morley et al. (1992) | Male SW mice | ICV | Effect of PACAP on food intake | PACAP, NPY | PACAP reduced basal and NPY-stimulated food intake. |
| Mounien et al. (2006) | Male Swiss albinos CD1 and C57BL/6 | Central injection | Expression of PACAP by NPY in ARC | PACAP, NPY, PAC1R, VPACR | PACAP may regulate the activity of NPY, and the inhibitory effect of PACAP on food intake may be affected by NPY neurotransmission. |
| Mounien et al. (2009) | Unspecified Rat | ICV | Role of PACAP through Melanocortin system | VIP, PACAP, SHU9119 | A high proportion of POMC neurons express PACAP receptors. PACAP may inhibit food intake through a melanocortin-dependent pathway. |
| Yokota et al. (1995) | Male Wistar-lmamichi rat | Infusion | Effect of PACAP on glucose | PACAP, theophylline, VIP, glucagon | PACAP stimulates glucose output from the perfused rat liver, and Ca2+ rather than cAMP plays an important role in this action. |
| Yada et al. (1994) | Unspecified Rat | Not Specified | Effect of PACAP on Insulin Secretion | PACAP, VIP, insulin | PACAP increases insulin release from rat islets in a glucose-dependent manner. VIP is less potent. |
| Yada et al. (2000) | Female GK rats and C57B/6J mice | IP | PACAP effect in blood glucose in GK rats (diabetic) and normal high fat diet mice (glucose intolerant) | PACAP, insulin | PACAP reduced hyperglycemia in GK rats and mice fed an HFD. |
| Yu et al. (2008) | Male NIH Mice | IP | Effect of chronic maxadilan on glucose levels and insulin sensitivity | M65, maxadilan, insulin | Maxadilan-induced hyperglycemia was blocked by M65. Acute maxadilan decreased feeding. Chronic maxadilan increased body weight and decreased body fat. PACAP increased glucose tolerance and insulin sensitivity. |
| Vu et al. (2015) | PAC1R (−/−) and (+/+) | IP | Effect of PACAP on other neurotransmitters to decrease food intake | PACAP, ghrelin, GLP-1, leptin, insulin, glucagon, PYY | PACAP decreased food intake and ghrelin in wild-type but not KO mice. Postprandial levels of active ghrelin and insulin were different in fasted KO than WT mice. |
| Tachibana et al. (2004) | Male Chicks | ICV | Effect of CRH on PACAP and VIP in chicks | PACAP, VIP, astressin, PACAP6-38 | CRF neurons modulate PACAP- and VIP-induced hypophagia. |
| Sites of PACAP action | |||||
| Resch et al. (2011) | Male Sprague–Dawley rats | Microinjections | Effect of PACAP in VMN on food intake and thermogenesis | PACAP, VMN | PACAP input to the VMN can influence energy homeostasis. |
| Resch et al. (2013) | Male Sprague–Dawley rats | Microinjections | PACAP regulates feeding | VMN, PVN, PACAP. VIP 6-28, PACAP6-38 | PACAP injected in the PVN decreased meal size, duration of the meal, and total time spent eating. PACAP in both the PVN and VMN increased plasma glucose but not pancreatic hormone levels. PACAP in the VMN increased core body temperature and spontaneous locomotor activity. |
| Kocho-Schellenberg et al. (2014) | Male and female Sprague–Dawley rats | ICV injection of 1.0 μg/0.25 μL PACAP38 | PACAP BNST localization on food/water intake | PACAP38, BNST | Infusion of PACAP38 in the posterior (but not anterior) BNST rats induced hypophagic effects. |
| Iemolo et al. (2015) | Male Wistar rats | PACAP 0–1 ug/rat in the CeA and BlA | Role of PACAP in response to stress | CeA/PACAP | PACAP in the CeA exerts anorectic effects via melanocortin and TrKB. |
| Hurley, M.M., et al. (2016) | Male Sprague–Dawley rats | PACAP 50 pmol/0.25 uL/side/AMPA 74.5 ng/side/Intra VMN administration | Role of PACAP in obesity | VMN/PACAP, AMPA | PACAP injection into the NAc reduces hedonic feeding, while its injection into the VMN decreases homeostatic feeding. |
| Liu et al. (2015) | N/A | N/A | Effects of PACAP on Hedonic Food Intake | ARC, LHA, VTA, NAc, GABA, DA, POMC, NPY, AgRP | Researchers observed the interaction between food intake and reward circuits in the NAc and VTA. |
| Adams et al. (2008) | Male and female PAC1 (−/−) and (+/+) | N/A | Role of endogenous PACAP | Hypothalamic nuclei | PACAP is involved in maintaining energy homeostasis, possibly via cold-sensing mechanisms. |
| Nguyen et al. (2020) | Male PACAP (−/−) and (+/+) | ICV | Effect on PACAP in VMH on AgRP | PACAP. VIP 6-28, PACAP6-38, AgRP, VMH | PACAP regulates feeding in VMH by modulating the expression of AgRP. Knockout mice fasted for two days had significantly lower food consumption during refeeding than PACAP (+/+) mice. |
| Chance et al. (1995) | Male Sprague–Dawley rats | IHT Injections of CRG, PACAP, and NPY | Effect of PACAP on NPY-dependent food intake | NPY, PACAP, hypothalamus | PACAP did not alter the hypothalamic NPY level but reduced food intake when given before NPY. |
| Role of PACAP receptor(s) | |||||
| Dore et al. (2013) | Male Wistar rats | ICV PACAP (PACAP-38) and ICV D-Phe-CRF (12–41) | PACAP and CRF signaling pathways in PVN | PACAP and CRF signaling pathways in PVN | CRF may mediate the anxiogenic and anti-rewarding effects of PACAP but not its anorectic action. |
| Signaling | |||||
| Resch et al. (2014) | Male Sprague–Dawley rats | Injection | NMDA mediates PACAP in VMH | VMN, glutamate, Src kinase, PACAP | PACAP in the VMN increases glutamate signaling through NMDA receptors and phosphorylation of the Src kinase to regulate food intake. |
| Homeostatic and Hedonic Aspects | |||||
| Le et al. (2021) | Male and female C57BL/6 mice | Local injection | Pituitary Adenylate Cyclase Activating Polypeptide Inhibits A 10 Dopamine Neurons and Suppresses the Binge-like Consumption of Palatable Food | VMN, VTA, DA | PACAP injected into the VTA reduced hedonic food intake and dopaminergic neuronal activity. |
| Chang et al. (2020) | Male and female mice | IV | Effect of PACAP on POMC neurons in mediating homeostatic food intake | PACAP, CNO, E2, ETOH, STX, PACAP6-38 | PACAP-stimulated POMC neurons were attenuated in mice on HFD. Elevation of PACAP in the VMN and ARC by chemogenetic and optogenetic stimulation decreased energy intake and increased energy expenditure. |
| Chang et al. (2019) | Male and female eGFP-POMC mice, PACAP-Cre mice | Local injection | Effect of PACAP on POMC neurons in mediating homeostatic food intake | POMC, TRPC channels, VMN, ARC | PACAP activates POMC neurons via the PAC1 R involving TRPC5 channels at the VMN and ARC, mediated by estradiol, although these effects are not seen in HFD mice. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sureshkumar, K.; Saenz, A.; Ahmad, S.M.; Lutfy, K. The PACAP/PAC1 Receptor System and Feeding. Brain Sci. 2022, 12, 13. https://doi.org/10.3390/brainsci12010013
Sureshkumar K, Saenz A, Ahmad SM, Lutfy K. The PACAP/PAC1 Receptor System and Feeding. Brain Sciences. 2022; 12(1):13. https://doi.org/10.3390/brainsci12010013
Chicago/Turabian StyleSureshkumar, Keerthana, Andrea Saenz, Syed M. Ahmad, and Kabirullah Lutfy. 2022. "The PACAP/PAC1 Receptor System and Feeding" Brain Sciences 12, no. 1: 13. https://doi.org/10.3390/brainsci12010013
APA StyleSureshkumar, K., Saenz, A., Ahmad, S. M., & Lutfy, K. (2022). The PACAP/PAC1 Receptor System and Feeding. Brain Sciences, 12(1), 13. https://doi.org/10.3390/brainsci12010013