
The Bacterial Enzyme RfxCas13d Is Less Neurotoxic Than PspCas13b and Could Be a Promising RNA Editing and Interference Tool in the Nervous System


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
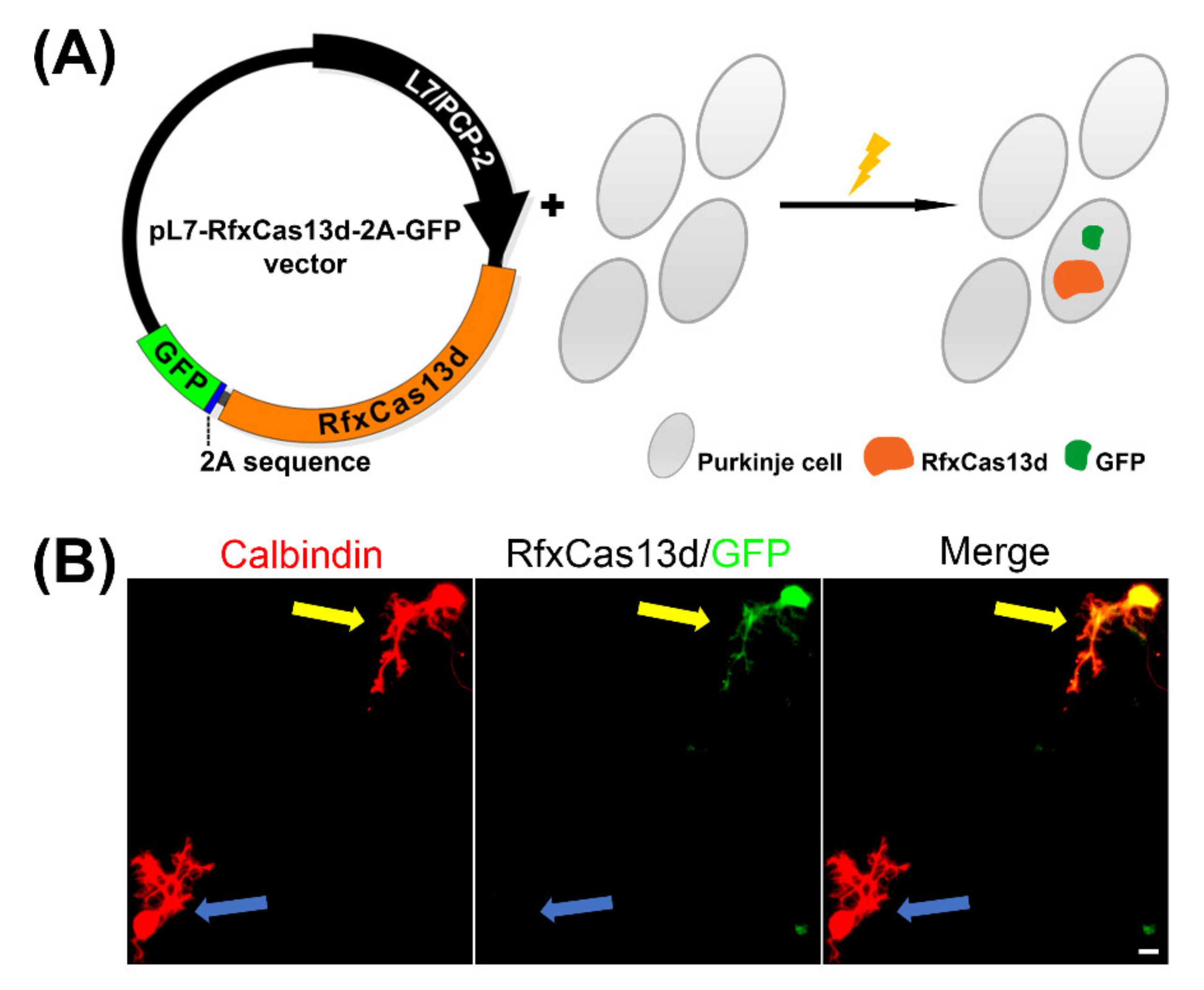
2.1. Purkinje Cell-Specific CRISPR-RfxCas13d Expression in Cerebellar Purkinje Neuron Cultures
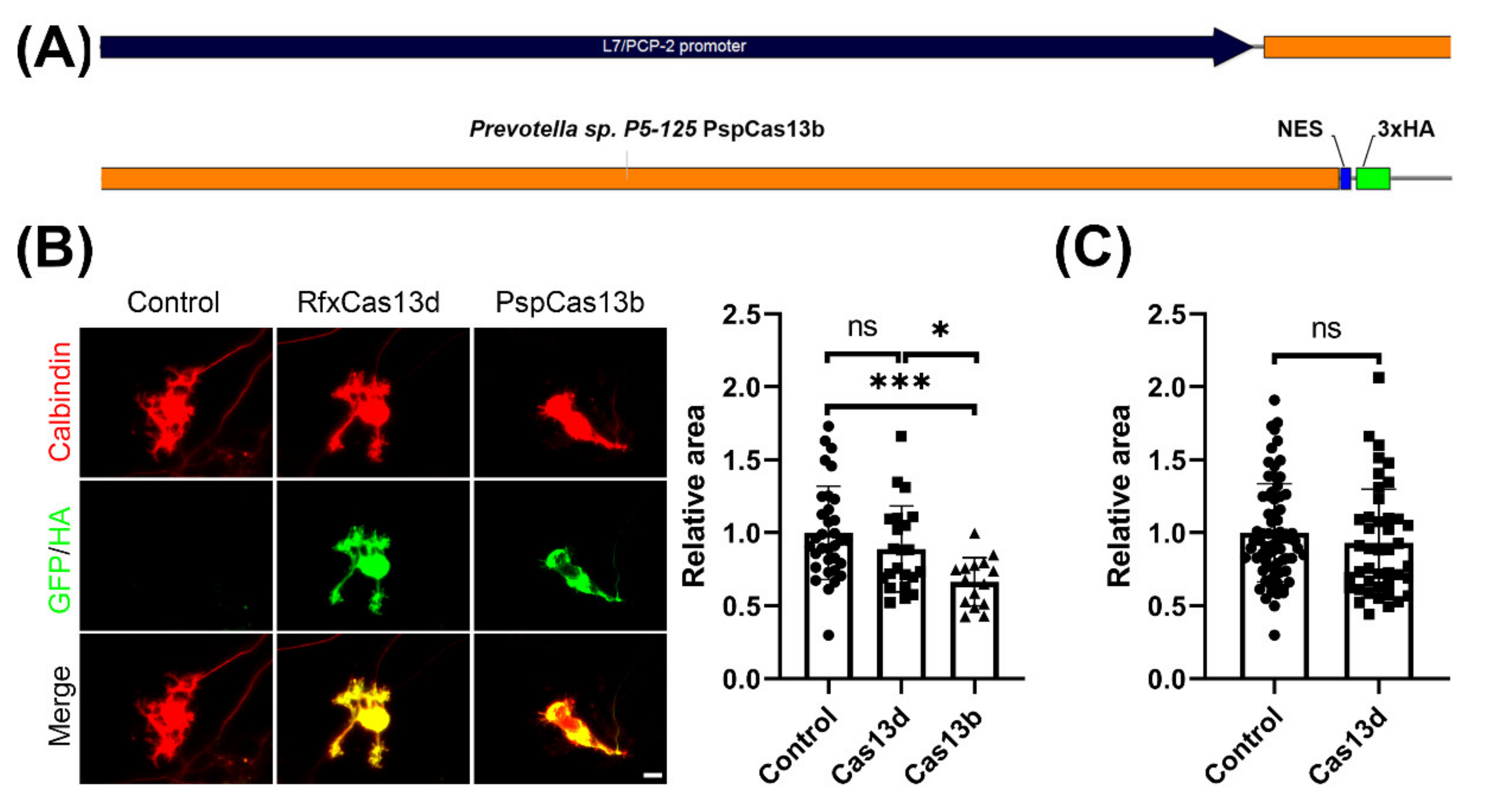
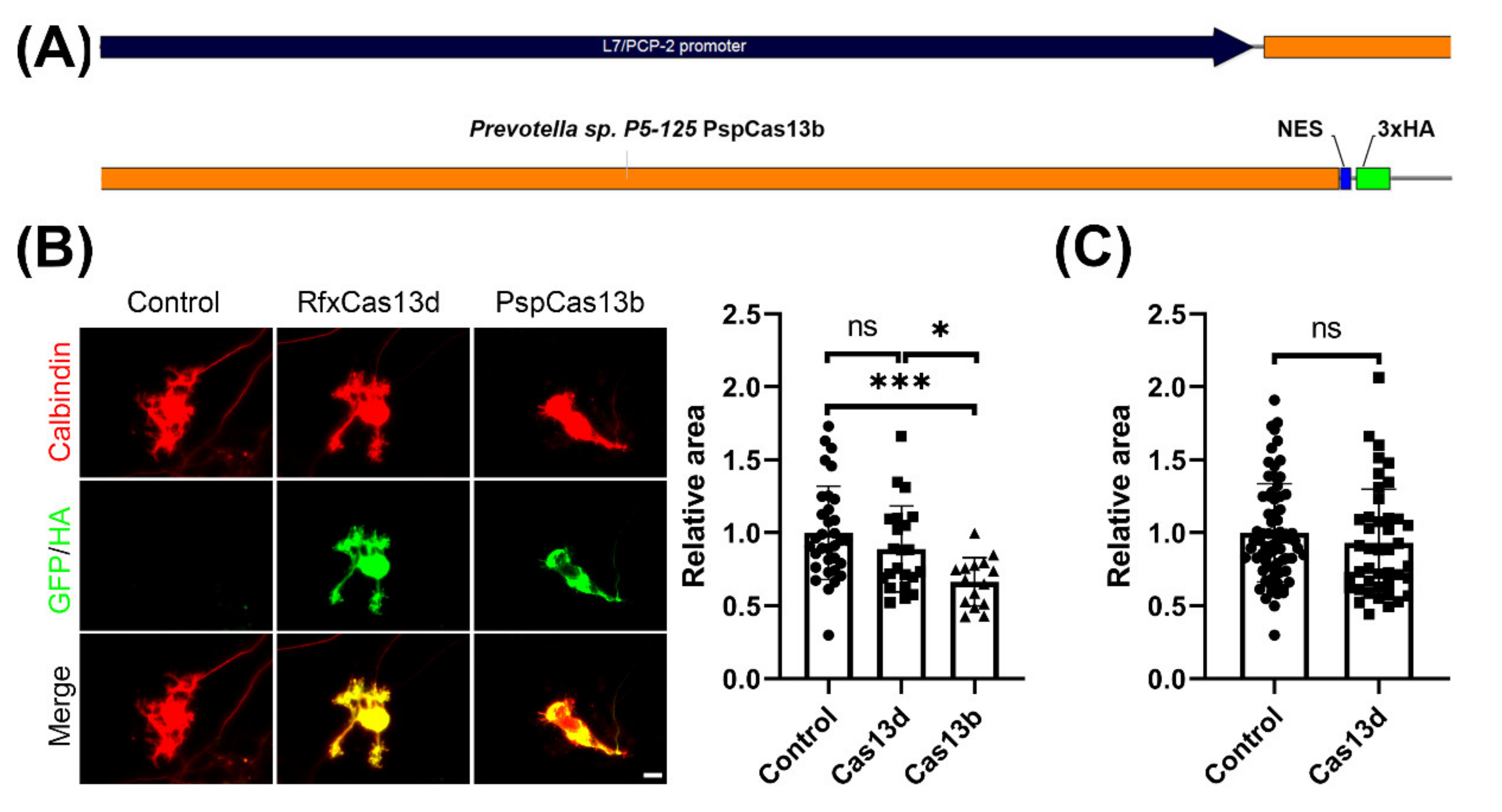
2.2. CRISPR-PspCas13b, But Not CRISPR-RfxCas13d, Impedes Development of Cultured Purkinje Neurons
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Plasmid Construction
4.3. Cultures of Cerebellar Purkinje Cells and Electroporation Procedure
4.4. Immunocytochemistry
4.5. Quantitative Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferlini, A.; Goyenvalle, A.; Muntoni, F. RNA-targeted drugs for neuromuscular diseases. Science 2021, 371, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Reardon, S. Step aside CRISPR, RNA editing is taking off. Nature 2020, 578, 24–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Roy, S. Gene-based therapies for neurodegenerative diseases. Nat. Neurosci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Su, J.; Hu, X.; Zhou, C.; Li, H.; Chen, Z.; Xiao, Q.; Wang, B.; Wu, W.; Sun, Y.; et al. Glia-to-Neuron Conversion by CRISPR-CasRx Alleviates Symptoms of Neurological Disease in Mice. Cell 2020, 181, 590–603.e16. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.B.T.; Gootenberg, J.S.; Abudayyeh, O.O.; Franklin, B.; Kellner, M.J.; Joung, J.; Zhang, F. RNA editing with CRISPR-Cas13. Science 2017, 358, 1019–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.-W.; Kapfhammer, J.P. Conditional gene silencing via a CRISPR system in cerebellar Purkinje cells. Biochimica et Biophysica Acta Gen. Subj. 2021, 1865, 129869. [Google Scholar] [CrossRef] [PubMed]
- Abudayyeh, O.O.; Gootenberg, J.S.; Franklin, B.; Koob, J.; Kellner, M.J.; Ladha, A.; Joung, J.; Kirchgatterer, P.; Cox, D.B.T.; Zhang, F. A cytosine deaminase for programmable single-base RNA editing. Science 2019, 365, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA targeting with CRISPR-Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konermann, S.; Lotfy, P.; Brideau, N.J.; Oki, J.; Shokhirev, M.N.; Hsu, P.D. Transcriptome Engineering with RNA-Targeting Type VI-D CRISPR Effectors. Cell 2018, 173, 665–676.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushawah, G.; Hernandez-Huertas, L.; Abugattas-Nuñez del Prado, J.; Martinez-Morales, J.R.; DeVore, M.L.; Hassan, H.; Moreno-Sanchez, I.; Tomas-Gallardo, L.; Diaz-Moscoso, A.; Monges, D.E.; et al. CRISPR-Cas13d Induces Efficient mRNA Knockdown in Animal Embryos. Dev. Cell 2020, 54, 805–817.e7. [Google Scholar] [CrossRef] [PubMed]
- Buchman, A.; Brogan, D.J.; Sun, R.; Yang, T.; Hsu, P.; Hsu, P.; Akbari, O.S.; Akbari, O.S. Programmable RNA Targeting Using CasRx in Flies. Cris. J. 2020, 3, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.-W.; Kapfhammer, J.P. The Bacterial Enzyme Cas13 Interferes with Neurite Outgrowth from Cultured Cortical Neurons. Toxins 2021, 13, 262. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.W.; Kapfhammer, J.P. The CRISPR-Cas13a system interferes with Purkinje cell dendritic development. Biochimica et Biophysica Acta Mol. Cell Res. 2020, 1867, 118710. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; McCroskery, S.; Hammer, J.A. An efficient method for the long-term and specific expression of exogenous cDNAs in cultured Purkinje neurons. J. Neurosci. Methods 2011, 200, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.E.; Brameld, J.M.; Hill, P.; Barrett, P.; Ebling, F.J.P.; Jethwa, P.H. The use of a viral 2A sequence for the simultaneous over-expression of both the vgf gene and enhanced green fluorescent protein (eGFP) in vitro and in vivo. J. Neurosci. Methods 2015, 256, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.-W.; Kapfhammer, J.P. Modulation of Increased mGluR1 Signaling by RGS8 Protects Purkinje Cells From Dendritic Reduction and Could Be a Common Mechanism in Diverse Forms of Spinocerebellar Ataxia. Front. Cell Dev. Biol. 2021, 8, 1912. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.-W.; Kapfhammer, J.P. The Bacterial Enzyme RfxCas13d Is Less Neurotoxic Than PspCas13b and Could Be a Promising RNA Editing and Interference Tool in the Nervous System. Brain Sci. 2021, 11, 1054. https://doi.org/10.3390/brainsci11081054
Wu Q-W, Kapfhammer JP. The Bacterial Enzyme RfxCas13d Is Less Neurotoxic Than PspCas13b and Could Be a Promising RNA Editing and Interference Tool in the Nervous System. Brain Sciences. 2021; 11(8):1054. https://doi.org/10.3390/brainsci11081054
Chicago/Turabian StyleWu, Qin-Wei, and Josef P. Kapfhammer. 2021. "The Bacterial Enzyme RfxCas13d Is Less Neurotoxic Than PspCas13b and Could Be a Promising RNA Editing and Interference Tool in the Nervous System" Brain Sciences 11, no. 8: 1054. https://doi.org/10.3390/brainsci11081054
APA StyleWu, Q.-W., & Kapfhammer, J. P. (2021). The Bacterial Enzyme RfxCas13d Is Less Neurotoxic Than PspCas13b and Could Be a Promising RNA Editing and Interference Tool in the Nervous System. Brain Sciences, 11(8), 1054. https://doi.org/10.3390/brainsci11081054