
Neural Alterations in Interpersonal Distance (IPD) Cognition and Its Correlation with IPD Behavior: A Systematic Review


Abstract
:1. Introduction
2. Materials and Methods
2.1. Eligibility Criteria
2.1.1. Included Studies
2.1.2. Included IPD Paradigms
2.1.3. Included Subjects
2.1.4. Included Results
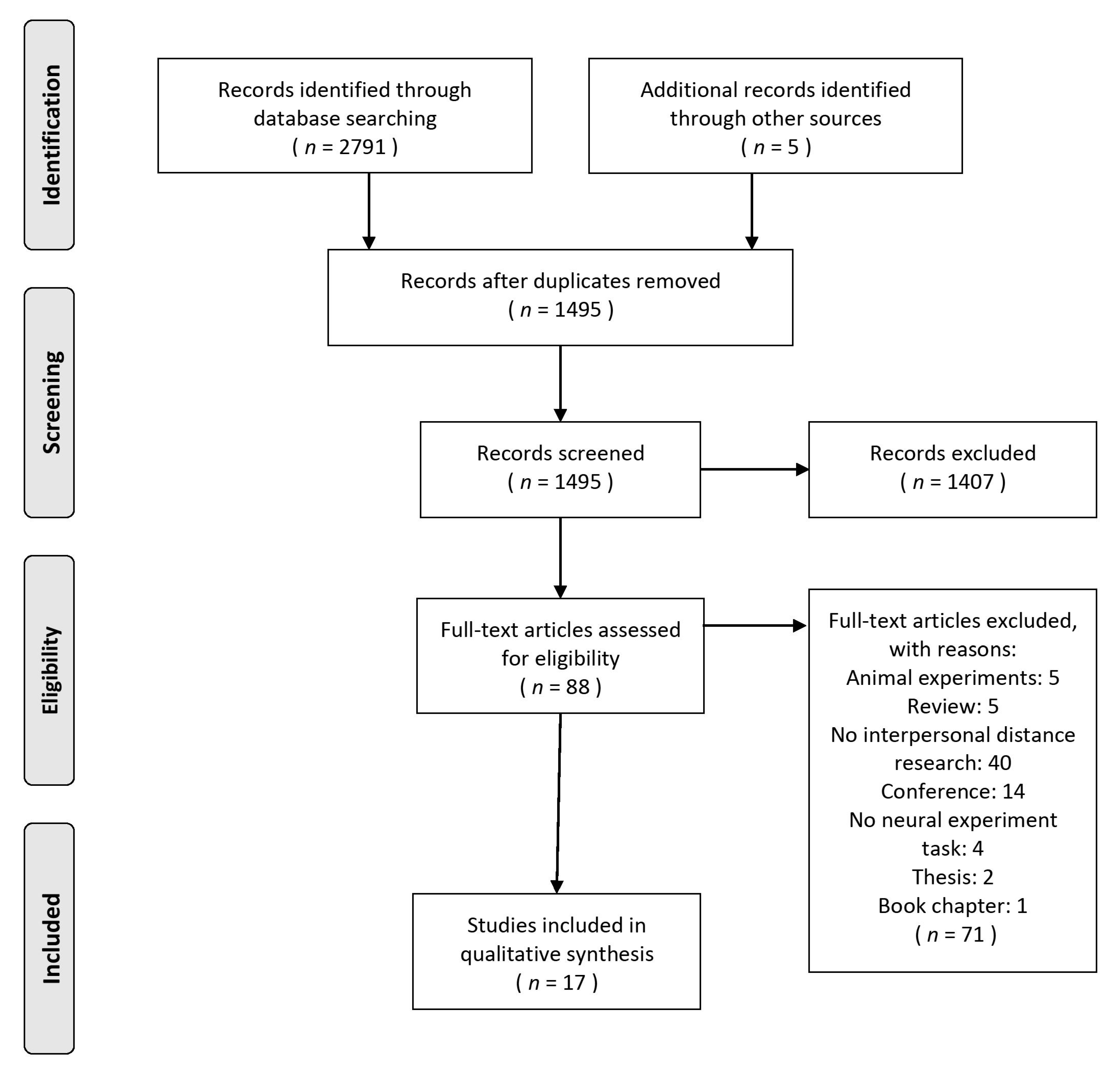
2.2. Search Methods and Study Selection
2.3. Quality Assessment of Each Study
3. Results
3.1. Quality Assessment of Each Study
3.2. Basic Information of the Studies
3.3. Study Design
3.4. Neural Activity of IPD Processing
3.4.1. Neural Activity of the fMRI Findings
3.4.2. Functional Connectivity Response to IPD Tasks
3.4.3. ERPs in Response to IPD Tasks
3.4.4. Correlation between IPD and Neural Activity
4. Discussion
5. Limitation and Future Research
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- APA Dictionary of Psychology. Available online: https://dictionary.apa.org/interpersonal-distance (accessed on 28 July 2021).
- Coello, Y.; Cartaud, A. The interrelation between peripersonal action space and interpersonal social space: Psychophysiological evidence and clinical implications. Front. Hum. Neurosci. 2021, 15, 92. [Google Scholar] [CrossRef]
- Kennedy, D.P.; Adolphs, R. Violations of Personal Space by Individuals with Autism Spectrum Disorder. PLoS ONE 2014, 9, e103369. [Google Scholar] [CrossRef]
- Ruggiero, G.; Frassinetti, F.; Coello, Y.; Rapuano, M.; Di Cola, A.S.; Iachini, T. The effect of facial expressions on peripersonal and interpersonal spaces. Psychol. Res. 2017, 81, 1232–1240. [Google Scholar] [CrossRef]
- Cartaud, A.; Ruggiero, G.; Ott, L.; Iachini, T.; Coello, Y. Physiological response to facial expressions in peripersonal space determines interpersonal distance in a social interaction context. Front. Psychol. 2018, 9, 657. [Google Scholar] [CrossRef]
- Cartaud, A.; Ott, L.; Iachini, T.; Honoré, J.; Coello, Y. The influence of facial expression at perceptual threshold on electrodermal activity and social comfort distance. Psychophysiology 2020, 57, e13600. [Google Scholar] [CrossRef]
- Candini, M.; Giuberti, V.; Manattini, A.; Grittani, S.; di Pellegrino, G.; Frassinetti, F. Personal space regulation in childhood autism: Effects of social interaction and person’s perspective. Autism Res. 2017, 10, 144–154. [Google Scholar] [CrossRef]
- Burgoon, J.K.; Jones, S.B. Toward a theory of personal space expectations and their violations. Hum. Commun. Res. 1976, 2, 131–146. [Google Scholar] [CrossRef]
- Kennedy, D.P.; Gläscher, J.; Tyszka, J.M.; Adolphs, R. Personal space regulation by the human amygdala. Nat. Neurosci. 2009, 12, 1226–1227. [Google Scholar] [CrossRef]
- Munyon, T.P. An Investigation of Interpersonal Distance and Relationship Quality at Work; The Florida State University: Tallahassee, FL, USA, 2009. [Google Scholar]
- Hall, E.T. A system for the notation of proxemic behavior 1. Am. Anthropol. 1963, 65, 1003–1026. [Google Scholar] [CrossRef]
- Sorokowska, A.; Sorokowski, P.; Hilpert, P.; Cantarero, K.; Frackowiak, T.; Ahmadi, K.; Alghraibeh, A.M.; Aryeetey, R.; Bertoni, A.; Bettache, K. Preferred interpersonal distances: A global comparison. J. Cross Cult. Psychol. 2017, 48, 577–592. [Google Scholar] [CrossRef]
- Adler, L.L.; Iverson, M.A. Interpersonal distance as a function of task difficulty, praise, status orientation, and sex of partner. Percept. Mot. Skills 1974, 39, 683–692. [Google Scholar] [CrossRef]
- Rapuano, M.; Sbordone, F.L.; Borrelli, L.O.; Ruggiero, G.; Iachini, T. The Effect of Facial Expressions on Interpersonal Space: A Gender Study in Immersive Virtual Reality. In Progresses in Artificial Intelligence and Neural Systems; Springer: Singapore, 2021; pp. 477–486. [Google Scholar]
- Iachini, T.; Coello, Y.; Frassinetti, F.; Senese, V.P.; Galante, F.; Ruggiero, G. Peripersonal and interpersonal space in virtual and real environments: Effects of gender and age. J. Environ. Psychol. 2016, 45, 154–164. [Google Scholar] [CrossRef]
- Hartnett, J.J.; Bailey, K.G.; Hartley, C.S. Body height, position, and sex as determinants of personal space. J. Psychol. 1974, 87, 129–136. [Google Scholar] [CrossRef]
- Pazhoohi, F.; Silva, C.; Lamas, J.; Mouta, S.; Santos, J.; Arantes, J. The effect of height and shoulder-to-hip ratio on interpersonal space in virtual environment. Psychol. Res. 2019, 83, 1184–1193. [Google Scholar] [CrossRef]
- Perry, A.; Nichiporuk, N.; Knight, R.T. Where does one stand: A biological account of preferred interpersonal distance. Soc. Cogn. Affect. Neurosci. 2016, 11, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Kaitz, M.; Bar-Haim, Y.; Lehrer, M.; Grossman, E. Adult attachment style and interpersonal distance. Attach. Hum. Dev. 2004, 6, 285–304. [Google Scholar] [CrossRef]
- Bogović, A.; Ivezić, E.; Filipčić, I. Personal Space of War Veterans with PTSD-Some Characteristics and Comparison with Healthy Individuals. Psychiatr. Danub. 2016, 28, 77–81. [Google Scholar]
- Nandrino, J.L.; Ducro, C.; Iachini, T.; Coello, Y. Perception of peripersonal and interpersonal space in patients with restrictive-type anorexia. Eur. Eat. Disord. Rev. 2017, 25, 179–187. [Google Scholar] [CrossRef]
- Gifford, R.; Sacilotto, P.A. Social isolation and personal space: A field study. Can. J. Behav. Sci. 1993, 25, 165. [Google Scholar] [CrossRef]
- Layden, E.A.; Cacioppo, J.T.; Cacioppo, S. Loneliness predicts a preference for larger interpersonal distance within intimate space. PLoS ONE 2018, 13, e0203491. [Google Scholar] [CrossRef] [Green Version]
- Vranic, A. Personal space in physically abused children. Environ. Behav. 2003, 35, 550–565. [Google Scholar] [CrossRef]
- Maier, A.; Gieling, C.; Heinen-Ludwig, L.; Stefan, V.; Schultz, J.; Güntürkün, O.; Becker, B.; Hurlemann, R.; Scheele, D. Association of childhood maltreatment with interpersonal distance and social touch preferences in adulthood. Am. J. Psychiatry 2020, 177, 37–46. [Google Scholar] [CrossRef]
- Di Pellegrino, G.; Làdavas, E. Peripersonal space in the brain. Neuropsychologia 2015, 66, 126–133. [Google Scholar] [CrossRef]
- Vagnoni, E.; Lewis, J.; Tajadura-Jiménez, A.; Cardini, F. Listening to a conversation with aggressive content expands the interpersonal space. PLoS ONE 2018, 13, e0192753. [Google Scholar] [CrossRef] [Green Version]
- Tajadura-Jiménez, A.; Pantelidou, G.; Rebacz, P.; Västfjäll, D.; Tsakiris, M. I-space: The effects of emotional valence and source of music on interpersonal distance. PLoS ONE 2011, 6, e26083. [Google Scholar] [CrossRef]
- Adams, L.; Zuckerman, D. The effect of lighting conditions on personal space requirements. J. Gen. Psychol. 1991, 118, 335–340. [Google Scholar] [CrossRef]
- Sawada, Y. Blood pressure and heart rate responses to an intrusion on personal space. Jpn. Psychol. Res. 2003, 45, 115–121. [Google Scholar] [CrossRef]
- Welsch, R.; Wessels, M.; Bernhard, C.; Thönes, S.; von Castell, C. Physical distancing and the perception of interpersonal distance in the COVID-19 crisis. Sci. Rep. 2021, 11, 11485. [Google Scholar] [CrossRef]
- Iachini, T.; Frassinetti, F.; Ruotolo, F.; Sbordone, F.L.; Ferrara, A.; Arioli, M.; Pazzaglia, F.; Bosco, A.; Candini, M.; Lopez, A. Social Distance during the COVID-19 Pandemic Reflects Perceived Rather Than Actual Risk. Int. J. Environ. Res. Public Health 2021, 18, 5504. [Google Scholar] [CrossRef]
- Cartaud, A.; Quesque, F.; Coello, Y. Wearing a face mask against COVID-19 results in a reduction of social distancing. PLoS ONE 2020, 15, e0243023. [Google Scholar] [CrossRef]
- Calbi, M.; Langiulli, N.; Ferroni, F.; Montalti, M.; Kolesnikov, A.; Gallese, V.; Umiltà, M.A. The consequences of COVID-19 on social interactions: An online study on face covering. Sci. Rep. 2021, 11, 2601. [Google Scholar] [CrossRef]
- Cooke, D.F.; Graziano, M.S.A. Defensive Movements Evoked by Air Puff in Monkeys. J. Neurophysiol. 2003, 90, 3317–3329. [Google Scholar]
- Graziano, M.S.; Hu, X.T.; Gross, C.G. Visuospatial Properties of Ventral Premotor Cortex. J. Neurophysiol. 1997, 77, 2268–2292. [Google Scholar]
- Lee, E.; Rhim, I.; Lee, J.W.; Ghim, J.-W.; Lee, S.; Kim, E.; Jung, M.W. Enhanced neuronal activity in the medial prefrontal cortex during social approach behavior. J. Neurosci. 2016, 36, 6926–6936. [Google Scholar] [CrossRef] [Green Version]
- Contestabile, A.; Casarotto, G.; Girard, B.; Tzanoulinou, S.; Bellone, C. Deconstructing the contribution of sensory cues in social approach. Eur. J. Neurosci. 2021, 53, 3199–3211. [Google Scholar] [CrossRef]
- Perry, A.; Lwi, S.J.; Verstaen, A.; Dewar, C.; Levenson, R.W.; Knight, R.T. The role of the orbitofrontal cortex in regulation of interpersonal space: Evidence from frontal lesion and frontotemporal dementia patients. Soc. Cogn. Affect. Neurosci. 2016, 11, 1894–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, A.; Rubinsten, O.; Peled, L.; Shamay-Tsoory, S.G. Don’t stand so close to me: A behavioral and ERP study of preferred interpersonal distance. NeuroImage 2013, 83, 761–769. [Google Scholar] [CrossRef]
- Holt, D.J.; Cassidy, B.S.; Yue, X.; Rauch, S.L.; Boeke, E.A.; Nasr, S.; Tootell, R.B.H.; Coombs, G. Neural Correlates of Personal Space Intrusion. J. Neurosci. 2014, 34, 4123–4134. [Google Scholar] [CrossRef] [Green Version]
- Holt, D.J.; Boeke, E.A.; Coombs, G.; Decross, S.N.; Cassidy, B.S.; Stufflebeam, S.; Rauch, S.L.; Tootell, R.B.H. Abnormalities in personal space and parietal-frontal function in schizophrenia. NeuroImage Clin. 2015, 9, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Schienle, A.; Wabnegger, A.; Schongassner, F.; Leutgeb, V. Effects of personal space intrusion in affective contexts: An fMRI investigation with women suffering from borderline personality disorder. Soc. Cogn. Affect. Neurosci. 2015, 10, 1424–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, D.; Perry, A.; Gilam, G.; Mayseless, N.; Gonen, T.; Hendler, T.; Shamay-Tsoory, S.G. The role of oxytocin in modulating interpersonal space: A pharmacological fMRI study. Psychoneuroendocrinology 2017, 76, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Schienle, A.; Wabnegger, A.; Leitner, M.; Leutgeb, V. Neuronal correlates of personal space intrusion in violent offenders. Brain Imaging Behav. 2017, 11, 454–460. [Google Scholar] [CrossRef] [Green Version]
- Vieira, J.B.; Tavares, T.P.; Marsh, A.A.; Mitchell, D.G.V. Emotion and personal space: Neural correlates of approach-avoidance tendencies to different facial expressions as a function of coldhearted psychopathic traits. Hum. Brain Mapp. 2017, 38, 1492–1506. [Google Scholar] [CrossRef]
- Ben Simon, E.; Walker, M.P. Sleep loss causes social withdrawal and loneliness. Nat. Commun. 2018, 9, 3146. [Google Scholar] [CrossRef]
- Cohen, D.; Perry, A.; Mayseless, N.; Kleinmintz, O.; Shamay-Tsoory, S.G. The role of oxytocin in implicit personal space regulation: An fMRI study. Psychoneuroendocrinology 2018, 91, 206–215. [Google Scholar] [CrossRef]
- Vieira, J.B.; Pierzchajlo, S.R.; Mitchell, D.G.V. Neural correlates of social and non-social personal space intrusions: Role of defensive and peripersonal space systems in interpersonal distance regulation. Soc. Neurosci. 2020, 15, 36–51. [Google Scholar] [CrossRef]
- Massaccesi, C.; Groessing, A.; Rosenberger, L.A.; Hartmann, H.; Candini, M.; di Pellegrino, G.; Frassinetti, F.; Silani, G. Neural Correlates of Interpersonal Space Permeability and Flexibility in Autism Spectrum Disorder. Cereb. Cortex 2021, 31, 2968–2979. [Google Scholar] [CrossRef]
- Nasiriavanaki, Z.; Barbour, T.; Farabaugh, A.H.; Fava, M.; Holmes, A.J.; Tootell, R.B.H.; Holt, D.J. Anxious attachment is associated with heightened responsivity of a parietofrontal cortical network that monitors peri-personal space. Neuroimage 2021, 30, 102585. [Google Scholar] [CrossRef]
- Perry, A.; Levy-Gigi, E.; Richter-Levin, G.; Shamay-Tsoory, S.G. Interpersonal distance and social anxiety in autistic spectrum disorders: A behavioral and ERP study. Soc. Neurosci. 2015, 10, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Wabnegger, A.; Leutgeb, V.; Schienle, A. Differential amygdala activation during simulated personal space intrusion by men and women. Neuroscience 2016, 330, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Rubinsten, O.; Korem, N.; Perry, A.; Goldberg, M.; Shamay-Tsoory, S. Different neural activations for an approaching friend versus stranger: Linking personal space to numerical cognition. Brain Behav. 2020, 10, e01613. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Becker, S.I.; Pegna, A.J. Attention is prioritised for proximate and approaching fearful faces. Cortex 2021, 134, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Fineberg, S.K.; Leavitt, J.; Landry, C.D.; Neustadter, E.S.; Lesser, R.E.; Stahl, D.S.; Deutsch-Link, S.; Corlett, P.R. Individuals with Borderline Personality Disorder show larger preferred social distance in live dyadic interactions. Psychiatry Res. 2018, 260, 384–390. [Google Scholar] [CrossRef]
- Hayduk, L.A. Personal space: An evaluative and orienting overview. Psychol. Bull. 1978, 85, 117. [Google Scholar] [CrossRef]
- Duke, M.P.; Nowicki, S. A new measure and social-learning model for interpersonal distance. J. Exp. Res. Personal. 1972, 6, 119–132. [Google Scholar]
- Noel, J.-P.; Serino, A.; Wallace, M.T. Increased neural strength and reliability to audiovisual stimuli at the boundary of peripersonal space. J. Cogn. Neurosci. 2019, 31, 1155–1172. [Google Scholar] [CrossRef]
- Felix-Ortiz, A.C.; Burgos-Robles, A.; Bhagat, N.D.; Leppla, C.A.; Tye, K.M. Bidirectional modulation of anxiety-related and social behaviors by amygdala projections to the medial prefrontal cortex. Neuroscience 2016, 321, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Spalding, K.N. The role of the medial prefrontal cortex in the generalization of conditioned fear. Neuropsychology 2018, 32, 1. [Google Scholar] [CrossRef]
- Hiser, J.; Koenigs, M. The multifaceted role of the ventromedial prefrontal cortex in emotion, decision making, social cognition, and psychopathology. Biol. Psychiatry 2018, 83, 638–647. [Google Scholar] [CrossRef]
- Janak, P.H.; Tye, K.M. From circuits to behaviour in the amygdala. Nature 2015, 517, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Wascher, E.; Hoffmann, S.; Sänger, J.; Grosjean, M. Visuo-spatial processing and the N1 component of the ERP. Psychophysiology 2009, 46, 1270–1277. [Google Scholar] [CrossRef]
- Berchicci, M.; Ten Brink, A.F.; Quinzi, F.; Perri, R.L.; Spinelli, D.; Di Russo, F. Electrophysiological evidence of sustained spatial attention effects over anterior cortex: Possible contribution of the anterior insula. Psychophysiology 2019, 56, e13369. [Google Scholar] [CrossRef] [PubMed]
- Fanghella, M.; Era, V.; Candidi, M. Interpersonal Motor Interactions Shape Multisensory Representations of the Peripersonal Space. Brain Sci. 2021, 11, 255. [Google Scholar] [CrossRef]
- Kwak, S.; Joo, W.-t.; Youm, Y.; Chey, J. Social brain volume is associated with in-degree social network size among older adults. Proc. R. Soc. B. Biol. Sci. 2018, 285, 1871. [Google Scholar] [CrossRef]
- Krabbe, S.; Gründemann, J.; Lüthi, A. Amygdala inhibitory circuits regulate associative fear conditioning. Biol. Psychiatry 2018, 83, 800–809. [Google Scholar] [CrossRef] [Green Version]
- Short, F.; Ward, R. Virtual limbs and body space: Critical features for the distinction between body space and near-body space. J. Exp. Psychol. Hum. Percept. Perform. 2009, 35, 1092. [Google Scholar] [CrossRef] [PubMed]
- Cléry, J.; Guipponi, O.; Wardak, C.; Hamed, S.B. Neuronal bases of peripersonal and extrapersonal spaces, their plasticity and their dynamics: Knowns and unknowns. Neuropsychologia 2015, 70, 313–326. [Google Scholar] [CrossRef]
- Canales-Johnson, A.; Billig, A.J.; Olivares, F.; Gonzalez, A.; Garcia, M.d.C.; Silva, W.; Vaucheret, E.; Ciraolo, C.; Mikulan, E.; Ibanez, A. Dissociable neural information dynamics of perceptual integration and differentiation during bistable perception. Cereb. Cortex 2020, 30, 4563–4580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vartanian, O.; Smith, I.; Lam, T.K.; King, K.; Lam, Q.; Beatty, E.L. The relationship between methods of scoring the alternate uses task and the neural correlates of divergent thinking: Evidence from voxel-based morphometry. NeuroImage 2020, 223, 117325. [Google Scholar] [CrossRef]
- Eyre, H.A.; Acevedo, B.; Yang, H.; Siddarth, P.; Van Dyk, K.; Ercoli, L.; Leaver, A.M.; Cyr, N.S.; Narr, K.; Baune, B.T. Changes in neural connectivity and memory following a yoga intervention for older adults: A pilot study. J. Alzheimers Dis. 2016, 52, 673–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riem, M.M.; Kunst, L.E.; Steenbakkers, F.D.; Kir, M.; Sluijtman, A.; Karreman, A.; Bekker, M.H. Oxytocin reduces interpersonal distance: Examining moderating effects of childrearing experiences and interpersonal context in virtual reality. Psychoneuroendocrinology 2019, 108, 102–109. [Google Scholar] [CrossRef]
- Iachini, T.; Coello, Y.; Frassinetti, F.; Ruggiero, G. Body space in social interactions: A comparison of reaching and comfort distance in immersive virtual reality. PLoS ONE 2014, 9, e111511. [Google Scholar]
- Quaresima, V.; Ferrari, M. Functional near-infrared spectroscopy (fNIRS) for assessing cerebral cortex function during human behavior in natural/social situations: A concise review. Organ. Res. Methods 2019, 22, 46–68. [Google Scholar] [CrossRef]
- Kroczek, L.O.; Pfaller, M.; Lange, B.; Müller, M.; Mühlberger, A. Interpersonal distance during real-time social interaction: Insights from subjective experience, behavior, and physiology. Front. Psychiatry 2020, 11, 561. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Author (et al.) (Year) | Participants | Age (M ± SD) | Gender (Male) | Handedness (R) | IPD Neural Task | Experiment Design | Image Technical |
|---|---|---|---|---|---|---|---|
| Anat Perry (et al.) (2013) [40] | 48 undergraduates low social anxiety (SA):22(12 female) high SA:22(10 female) (4 subjects were excluded from data analysis) | 27.5 ± 2.9 23–39 | 48 (24) | 40 | A modified computerized version of the comfortable interpersonal distance (CID) | 2 Figures (close friend/stranger) × 8 Radii (0°/45°/90°/135°/180°/225°, 270°/315°) | Event-related potential (ERP) |
| Daphne J. Holt (et al.) (2014) [41] | 8 healthy subjects 14 healthy subjects (later enrolled) | 26.4 ± 4.7 24.6 ± 4.5 | 8 (4) 14 (8) | - | Approach and Withdrawal stimuli | 2 Motions (approach/withdrawal) × 3 Stimuli (faces/cars/spheres) | Functional magnetic resonance imaging (fMRI) |
| Daphne J. Holt (et al.) (2015) [42] | 15 schizophrenia (SCZ) 14 healthy control (CON) | 30.1 ± 9.1 26.0 ± 6.5 | - | - | Approach and Withdrawal stimuli | 2 Motions (approach/withdrawal) × 2 Stimuli (faces/cars) | fMRI |
| Anat Perry (et al.) (2015) [53] | 13 typical participants 13 participants with Autistic Spectrum Disorder (ASD) | 24 ± 0.46 25 ± 1.24 | 13 (13) 13 (12) | 13 (12) 13 (13) | A modified version of the CID task | 2 Figures (Stranger/Friend) × 8 Entrances (0°/45°/90°/135°/180°/225°, 270°/315°) | ERP |
| Anne Schienle (et al.) (2015) [44] | 25 Borderline Personality Disorder (BPD) patients 25 healthy controls | 26.9 ±7.8 27.2 ±7.6 | 25 (0) 25 (0) | 20 20 | Still and approaching of facial expressions stimuli | 2 Motions (still/approaching) × 3 Facial expressions (angry/disgusted/neutral) | fMRI |
| Anat Perry (et al.) (2016) [18] | 42 undergraduate | 22.53 ± 4.29 | 42 (0) | 42 (41) | A modified version of the CID task | 3 Figures (male friend/male stranger/screen) × 4 Entrances (0°/90°/180°/270°) | Electroencephalograph (EEG) |
| Albert Wabnegger, Verena Leutgeb and Anne Schienle (2016) [54] | 30 healthy participants | 27.3± 8.1 | 30 (0) | - | Pictures of neutral facial expressions stimuli | 2 Motions (static/approaching) × 2 Faces (male/female) | fMRI |
| Daniela Cohen (et al.) (2017) [45] | 19 healthy participants | 26.05 ± 3.51 | 19 (19) | 19 | A modified version of the CID task | 2 Figures (friend/stranger) × 4 Entrances (0°/90°/180°/270°) | fMRI |
| Anne Schienle (et al.) (2017) [46] | 17 violent offenders18 nondelinquent controls | 34.82 ± 12.54 37.89 ± 9.21 | 17 (17) 18 (18) | - | Pictures of neutral facial expressions stimuli | 2 Motions (Static/approaching) × 2 Facial gender (man/women) | fMRI |
| Joana B. Vieira (et al.) (2017) [47] | 23 healthy participants | 20.96 ± 2.48 | 23 (11) | 23 | Approaching and withdrawal facial expressions stimuli | 2 Motions (approaching/withdrawal) × 5 Facial expressions (anger, fear, happiness, sadness, neutral) | fMRI |
| Eti Ben Simon, Matthew P. Walker (2018) [48] | 80 healthy adults | 20.2 ± 1.5 | 80 (71) | - | Social distance task—computerized | 2 Video stimuli (human/objects) | fMRI |
| Daniela Cohen (et al.) (2018) [49] | 24 healthy participants | 28.02 ± 2.69 | 24 (24) | 24 | Choice task of social and non-social stimuli | 2 Stimuli (social/non-social) × 3 Sizes (small/medium/larger distance) | fMRI |
| Orly Rubinsten (et al.) (2020) [55] | 11 developmental dyscalculia (DD) 12 typically developing controls (TD) | 29 ± 2 30 ± 4 | 11 (11) 12 (7) | - | A modified computerized version of the CID | 4 Figures (close friend/stranger/ball/artificial figure presented as a “griple”) × 8 Entrances (0°/45°/90°/135°/180°/225°, 270°/315°) | ERP |
| Joana B. Vieira, Stephen R. Pierzchajlo & Derek G.V. Mitchell (2020) [50] | 30 healthy volunteers | 21.90 ± 3.51 | 30 (6) | 30 | Experimental tasks: Social and non-social stimuli of varying threat levels | 2 Pictures (face/insects) × 2 Threats (high/low) × 2 Motions (approaching/withdrawal) | fMRI |
| Aimee Martin, Stefanie I. Becker and Alan J. Pegna (2021) [56] | Experiment1: 15 healthy subjects Experiment2: 30 healthy subjects | 22.73 ± 2.02 24.43 ± 9.66 | 15 (5) 30 (14) | 15 28 | Experiment 1: The task of judging letter changes with dynamic emotional faces Experiment 2: Gender judgment task | Experiment 1: 2 Face orientations (Upright, Inverted) × 2 Looming expression (Fearful, Neutral) × 2 Laterality of looming stimuli (Contralateral, Ipsilateral) Experiment 2: 2 Face expressions (Fearful, Neutral) × 4 Face gender combinations (male-male, female-female, male-female and female-male) × 2 spaces (50 cm near space,120 cm far space) | ERP |
| Claudia Massaccesi (et al.) (2021) [51] | 20 high-functioning ASD adults 20 controls | 34.25 ± 11.65 33.05 ± 12.33 | 20 (14) 20 (14) | - | Interpersonal Space task (Step 1–5 forward videos) | 5 Steps (step 1/step 2/step 3/step 4/step 5) | fMRI |
| Nasiriavanaki, Z. (et al.) (2021) [52] | 130 healthy subjects - a Reference Sample (80) - a Test Sample (50) | 19.45 ± 1.4 19.6 ± 1.4 | 80 (25) 50 (14) | - | Approach and Withdrawal stimuli | 2 Stimuli (human faces/cars) × 2 Gender (female face/male face) × 2 Motions (approaching/withdrawal) | fMRI |
| Author (et al.) (Year) | Participants | Age (M ± SD) | Gender (Male) | Handedness (R) | Data Analysis | Neural Findings | |
|---|---|---|---|---|---|---|---|
| Neural Activation | Functional Connectivity | ||||||
| Anat Perry (et al.) (2013) [40] | 48 undergraduates low SA:22 (12 female) high SA:22 (10 female) (4 subjects were excluded from data analysis) | 27.5 ± 2.9 23–39 | 48 (24) | 48 undergraduates low SA:22 (12 female) high SA:22 (10 female) (4 subjects were excluded from data analysis) | ERPs: N1, P1 Late Positive Potential (LLP) | ① High SA group > Low SA group ↓P1 ↓N1 ② Stranger condition > Friend condition ↑N1 ③ R Hemisphere > L Hemisphere (low SA group only) ↑N1 | - |
| Daphne J. Holt (et al.) (2014) [41] | 8 healthy subjects 14 healthy subjects (later enrolled) | 26.4 ± 4.7 24.6 ± 4.5 | 8 (4) 14 (8) | 8 healthy subjects 14 healthy subjects (later enrolled) | Region of interest (ROI): dorsal intraparietal sulcus (DIPS), ventral premotor cortex, precuneus (PMv) Whole-brain analysis | ① Approach faces > Withdrawal faces ↑DIPS, PMv, mid-cingulate gyrus, dorsal precentral, middle frontal gyri, middle occipital and inferior temporal gyri, and ventral superior parietal gyrus ② Faces > cars, Approach > Withdrawal ↑DIPS and PMv | Significant connectivity with the DIPS and PMv seed regions (Approaching > Withdrawing face stimuli) (n = 17) |
| Daphne J. Holt (et al.) (2015) [42] | 15 schizophrenia (SCZ) 14 healthy control (CON) | 30.1 ± 9.1 26.0 ± 6.5 | - | 15 schizophrenia (SCZ) 14 healthy control (CON) | ROI: DIPS, PMv Whole-brain analysis | ① Controls and schizophrenic patients: approaching faces > withdrawing faces: ↑DIPS and PMv ② Schizophrenic patients > Controls: ↑left DIPS, ↑left lateral frontal cortex,↑right middle temporal gyrus | DIPS-PMv connectivity (CON and SCZ) DIPS (Approaching > Withdrawing Faces) |
| Anat Perry (et al.) (2015) [53] | 13 typical participants 13 participants with ASD | 24 ± 0.46 25 ± 1.24 | 13 (13) 13 (12) | 13 typical participants 13 participants with ASD | ERP Analysis: N1, P1 | ① R hemisphere > L hemisphere ↑P1 (within all subjects) ② Stranger > friend ↑N1 | - |
| Anne Schienle (et al.) (2015) [44] | 25 BPD patients 25 healthy controls | 26.9 ± 7.8 27.2 ± 7.6 | 25 (0) 25 (0) | 25 BPD patients 25 healthy controls | ROI: amygdala, the insula, the premotor cortex, the putamen, and parietal regions Whole-brain voxel intensity tests | Approaching > Still:↑right amygdala, ↑several parietal regions (primary somatosensory cortex, inferior parietal region, intraparietal sulcus) | - |
| Anat Perry (et al.) (2016) [18] | 42 undergraduate | 22.53 ± 4.29 | 42 (0) | 42 undergraduate | Electroencephalograph (EEG) analysis: Alpha suppression, Hemispheres | ① High sensory sensitivity group > Low sensory sensitivity group ↑Alpha suppression,↑electrode O2 ② High sensory sensitivity group > low sensory sensitivity group, occipital sites > central sites > frontal sites ↑ Alpha suppression ③ Friend > computer screen ↑ Alpha suppression ④ Far distance > middle distance > near distance ↑Alpha suppression (occipital and central sites) | - |
| Albert Wabnegger, Verena Leutgeb and Anne Schienle (2016) [54] | 30 healthy participants | 27.3 ± 8.1 | 30 (0) | 30 healthy participants | ROI: amygdala, putamen, and parietal regions Whole-brain voxel intensity tests | ① Approaching stimuli > static stimuli:↑bilateral inferior, ↑superior parietal cortices,↑intraparietal sulci,↑left primary somatosensory cortex (SI), ↑occipital areas ② Approaching male stimuli > Approaching female stimuli: ↑R amygdala | - |
| Daniela Cohen (et al.) (2017) [45] | 19 healthy participants | 26.05 ± 3.51 | 19 (19) | 19 healthy participants | Whole-brain analysis | ① Friend > stranger: ↑R temporal lobe/occipital lobe/middle temporal gyrus,↑ L superior frontal gyrus/premotor/subthalamic nucleus/cingulate gyrus/inferior frontal gyrus/parahippocampal gyrus/middle occipital gyrus/inferior parietal lobule ② Oxytocin friend > Oxytocin stranger: ↑L medial prefrontal cortex, ↑R anterior cingulate, ↑R posterior-anterior cingulate, ↓R parahippocampal gyrus ③ Placebo friend > Placebo stranger: ↓L medial prefrontal cortex, ↓R anterior cingulate, ↓R posterior-anterior cingulate, ↑R parahippocampal gyrus | - |
| Anne Schienle (et al.) (2017) [46] | 17 violent offenders 18 nondelinquent controls | 34.82 ± 12.54 37.89 ± 9.21 | 17 (17) 18 (18) | 17 violent offenders 18 nondelinquent controls | ROI: amygdala, the insula, the premotor cortex, and parietal regions Whole brain voxel intensity tests | ① controls > offenders:↑inferior parietal ② Approaching > Static: ↑fronto-parietal regions (premotor cortex, SI, dorsolateral prefrontal cortex (DLPFC), superior/inferior parietal region),↑insula activation ③ Female faces > male faces: ↑Orbitofrontal cortex(OFC)activation ④ Offenders > Controls: Approaching > Static: ↑ insula activation ⑤ Offender > Controls, Male > Female: Approaching > Static:↑R insula activation ⑥ Male > Female, Approaching > Static: ↑L amygdala,↑L inferior parietal region | - |
| Joana B. Vieira (et al.) (2017) [47] | 23 healthy participants | 20.96 ± 2.48 | 23 (11) | 23 healthy participants | ROI: amygdala Whole-brain analysis | ① Approaching > Withdrawal: ↑bilateral visual cortex,↑fusiform gyrus, ↑R inferior parietal lobule (IPL),↑superior parietal lobules (SPL),↑R amygdala, ↑bilateral anterior insula (AI), DLPFC ② Happiness, Angry > fear, sadness, neutral:↑L dorsomedial prefrontal cortex (dmPFC),↑R OFC, R Inferior frontal gyrus (IFG),↑R Inferior parietal lobule (IPL) ③ Approaching > Withdrawal, Happiness, Angry > fear, sadness, neutral: ↑insula (bilaterally),↑L IFG ④ Approaching > Withdrawal, Sadness > happiness, angry, fear, neutral:↓insula (bilaterally) ⑤ Withdrawal > Approaching, sadness > happiness, angry, fear, neutral ↑ventrolateral prefrontal cortex (vlPFC) (bilaterally) ⑥ Approaching > Withdrawing, Happiness > sadness, angry, fear, neutral ↑vlPFC (bilaterally) | - |
| Eti Ben Simon, Matthew P. Walker (2018) [48] | 80 healthy adults | 20.2 ± 1.5 | 80 (71) | 80 healthy adults | ROI: Near Space network Theory of mind (ToM) network | ① Human approach > object approach, sleep-deprivation > Sleep rested:↑Near Space network (dorsal intraparietal sulcus and ventral premotor cortex) ② Human approach > object approach, Sleep rested > sleep-deprivation:↑Theory-of-Mind network (temporal–parietal junction and precuneus) | - |
| Daniela Cohen (et al.) (2018) [49] | 24 healthy participants | 28.02 ± 2.69 | 24 (24) | 24 healthy participants | ROI: Right dorsal striatum, dmPFC whole-brain analysis | ① social stimulus > non-social stimulus:↑R medial frontal gyrus ② Oxytocin > Placebo:↓L anterior cingulate (ACC),↓R culmen ③ Oxytocin > Placebo, social stimulus > non-social stimulus:↑R dorsal striatum | A connectivity Psychophysiological interaction (PPI) analysis dorsal striatum as a seed: Placebo condition: R Occipital lobe, L thalamus, R parietal lobe, L superior frontal gyrus, L occipital lobe Oxytocin condition: R occipital lobe, R putamen, L occipital lobe, L putamen |
| Orly Rubinsten (et al.) (2020) [55] | 11 developmental dyscalculia (DD) 12 typically developing controls (TD) | 29 ± 2 30 ± 4 | 11 (11) 12 (7) | 11 developmental dyscalculia (DD) 12 typically developing controls | ERP analysis: N1 | DD > TD:↑Latencies N1,↑N1 | - |
| Joana B. Vieira, Stephen R. Pierzchajlo & Derek G.V. Mitchell (2020) [50] | 30 healthy volunteers | 21.90 ± 3.51 | 30 (6) | 30 healthy volunteers | Whole-brain analysis | ① Social stimuli > Non-social stimuli (approach/withdrawal event): ↑R face fusiform area (FFA),↑bilateral temporoparietal junction (TPJ), and↑L medial prefrontal cortex (MPFC) ② First static image of each trail social stimuli > non-social stimuli:↑R TPJ, ↑bilateral FFA, ↑Ventromedial prefrontal cortex (vmPFC) ③ Approach > Withdrawal:↑R midbrain periaqueductal gray (PAG),↑R insula,↑R PMv extending to the dorsolateral prefrontal cortex,↑bilateral superior parietal lobule ④ Near > Far:↑midbrain (PAG) | Approach Social stimuli > Approach Non-social stimuli A midbrain PAG seed: bilateral premotor cortex and R dorsolateral prefrontal cortex |
| Aimee Martin, Stefanie I. Becker and Alan J. Pegna (2021) [56] | Experiment1: 15 healthy subjects Experiment2: 30 healthy subjects | 22.73 ± 2.02 24.43 ± 9.66 | 15 (5) 30 (14) | Experiment1: 15 healthy subjects Experiment2: 30 healthy subjects | ERPs analysis: N170, N2 posterior contralateral (N2pc) | Experiment 1: ① Looming fearful upright face contralateral amplitudes > ipsilateral amplitudes:↓l-N170,↓N2pc ② Upright looming fearful face > inverted neutral looming face: ↓l-N170 ③ Upright looming fearful face > inverted looming fearful face:↓N2pc Experiment 2: ④ Close faces > far faces:↑l-N170 ⑤ Contralateral amplitudes > ipsilateral amplitudes (fearful face):↓l-N170,↓N2pc (close distance) ⑥ fearful faces > neutral face (close/far distance):↓l-N170 | - |
| Claudia Massaccesi (et al.) (2021) [51] | 20 high-functioning ASD adults 20 controls | 34.25 ± 11.65 33.05 ± 12.33 | 20 (14) 20 (14) | 20 high-functioning ASD adults 20 controls | Task-based univariate fMRI analysis | CTR > ASD: bilateral dIPS, R human middle temporal visual area (hMT+/V5),L Fusiform Gyrus (FFA) | ASDs > CTRs:↑AMY- dIPS, AMY-FFA,↓FFA- dIPS,↑dIPS – AMY, ↓FFA -AMY |
| Nasiriavanaki, Z. (et al.) (2021) [52] | 130 healthy subjects - a Reference Sample (80) - a Test Sample (50) | 19.45 ± 1.4 19.6 ± 1.4 | 80 (25) 50 (14) | 130 healthy subjects - a Reference Sample (80) - a Test Sample (50) | ROIs: peripersonal space (PPS) network Whole-brain analysis | ① Peripersonal space (PPS)network responses (Face Approach > Withdrawal) ↑R and L superior frontal cortex (SFC), ↑R and L medial parietal cortex (MPC), ↑R and L superior parietal cortex (SPC) ② Inside > outside the personal space boundary:↑PPS network ③ Faces approach > cars approach: ↑PPS network | No significant correlations were found |
| Author (et al.) (Year) | Participants | Age (M ± SD) | Gender (Male) | Handedness (R) | IPD Task | IPD Index (Correlation) | Details of the Relationship between IPD and Neural Outcomes | r/z |
|---|---|---|---|---|---|---|---|---|
| Daphne J. Holt (et al.) (2014) [41] | 8 healthy subjects 14 healthy subjects (later enrolled) | 26.4 ± 4.7 24.6 ± 4.5 | 8 (4) 14 (8) | - | Stop-Distance paradigm | personal space size (pps) (−) personal space permeability (psp) (+) | ①↑DIPS-PMv(dorsal intraparietal sulcus-ventral premotor cortex) functional coupling,↓personal space size ②↑DIPS-PMv functional coupling,↑ personal space permeability | Pearson correlation: pps r = −0.55 * Voxelwise regression analyses: pps z1 = 3.4 ** (−) psp z2 = 3.6 ** (+) |
| Daphne J. Holt (et al.) (2015) [42] | 15 schizophrenia (SCZ) 14 healthy control (CON) | 30.1 ± 9.1 26.0 ± 6.5 | - | - | Stop-Distance paradigm | personal space size (+/−) personal space permeability | ①↑DIPS activation (approaching > withdrawing faces), ↑personal space size ②↑DIPS–PMv connectivity,↓personal space size | Pearson correlation: r (left DIPS) = 0.62 * (CON) r (right DIPS) = 0.56 * (SCZ) Voxelwise regression analyses: pps z (CON) = 3.53 ** (−) pps z (SCZ) = 3.47 ** (−) |
| Anat Perry (et al.) (2015) [53] | 13 typical participants 13 participants with ASD | 24 ± 0.46 25 ± 1.24 | 13 (13) 13 (12) | 13 (12) 13 (13) | Stop distance paradigm | Average preferred distances (+) | ASD group: ↑N1 ERP amplitude, ↑Average preferred distances | r = 0.62 * |
| Daniela Cohen (et al.) (2017) [45] | 19 healthy participants | 26.05 ± 3.51 | 19 (19) | 19 | A modified version of the comfortable interpersonal distance (CID) task | Distance index score -friend -stranger (+) -Oxytocin—Placebo (friend) -Oxytocin—Placebo (stranger) (−) | ①↓dmPFC,↓distance index score (stranger) ②↓dmPFC,↑Oxytocin - Placebo distance index score (stranger) | Not report |
| Joana B. Vieira (et al.) (2017) [47] | 23 healthy participants | 20.96 ± 2.48 | 23 (11) | 23 | Computerized Interpersonal Distance Task | Computerized desired distance -sadness (+) -happiness -fear (+) -angry (+) -neutral | ↑R amygdala ↑distance to angry,↑distance to sad,↑distance to fearful | r (angry) = 0.61 ** r (sad) = 0.527 ** r (fearful) = 0.504 * |
| Eti Ben Simon, Matthew P. Walker (2018) [48] | 80 healthy adults | 20.2 ± 1.5 | 80 (71) | - | Social distance task—computerized | sleep deprivation (SD) distance - sleep rested (SR) distance (+) | ↑Near Space network (human > object approach),↑social distance (Sleep deprivation-Sleep rest) | r = 0.53 * |
| Daniela Cohen (et al.) (2018) [49] | 24 healthy participants | 28.02 ± 2.69 | 24 (24) | 24 | Choice task of social and non-social stimuli | Distance behavioral scores Placebo condition (−) Oxytocin condition (+) | ↑Right dorsal striatum -↓distance scores (Placebo condition) -↑distance scores (Oxytocin condition) | r = −0.1 (Placebo) r = 0.3 (Oxytocin) |
| Joana B. Vieira, Stephen R. Pierzchajlo & Derek G.V. Mitchell (2020) [50] | 30 healthy volunteers | 21.90 ± 3.51 | 30 (6) | 30 | Computerized distance task “Stop-distance” task | percentage of stimulus (−) physical distance (−) | ↑connectivity strength between the midbrain and the left premotor cortex ↓percentage of stimulus ↓physical distance | r1 = −0.409 * r2 = −0.374 * |
| Claudia Massaccesi (et al.) (2021) [51] | 20 high-functioning ASD adults 20 controls | 34.25 ± 11.65 33.05 ± 12.33 | 20 (14) 20 (14) | - | Interpersonal Space Task | Averaged comfort ratings(+/−) | ↑L fusiform face area (FFA),↑Averaged comfort rating (all participants) ↓comfort,↑connectivity from FFA to the amygdala | r = 0.455 * connection parameters: 0.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Izumi, S.-I. Neural Alterations in Interpersonal Distance (IPD) Cognition and Its Correlation with IPD Behavior: A Systematic Review. Brain Sci. 2021, 11, 1015. https://doi.org/10.3390/brainsci11081015
Huang X, Izumi S-I. Neural Alterations in Interpersonal Distance (IPD) Cognition and Its Correlation with IPD Behavior: A Systematic Review. Brain Sciences. 2021; 11(8):1015. https://doi.org/10.3390/brainsci11081015
Chicago/Turabian StyleHuang, Xinxin, and Shin-Ichi Izumi. 2021. "Neural Alterations in Interpersonal Distance (IPD) Cognition and Its Correlation with IPD Behavior: A Systematic Review" Brain Sciences 11, no. 8: 1015. https://doi.org/10.3390/brainsci11081015
APA StyleHuang, X., & Izumi, S.-I. (2021). Neural Alterations in Interpersonal Distance (IPD) Cognition and Its Correlation with IPD Behavior: A Systematic Review. Brain Sciences, 11(8), 1015. https://doi.org/10.3390/brainsci11081015