
Comparison of Four fMRI Paradigms Probing Emotion Processing

 , , ,
, , , {kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Tasks
2.2.1. EMOBACK Task
2.2.2. FACES Task
2.2.3. IAPS Task
2.2.4. OASIS Task
2.3. Data Collection
2.4. Data Analysis
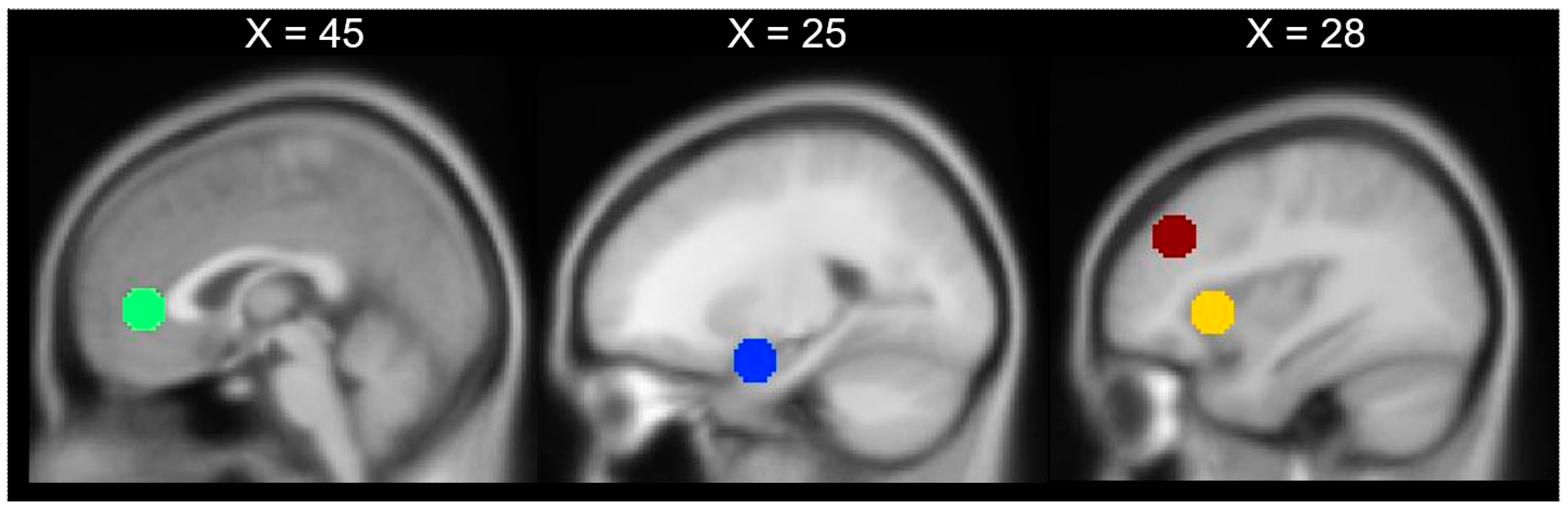
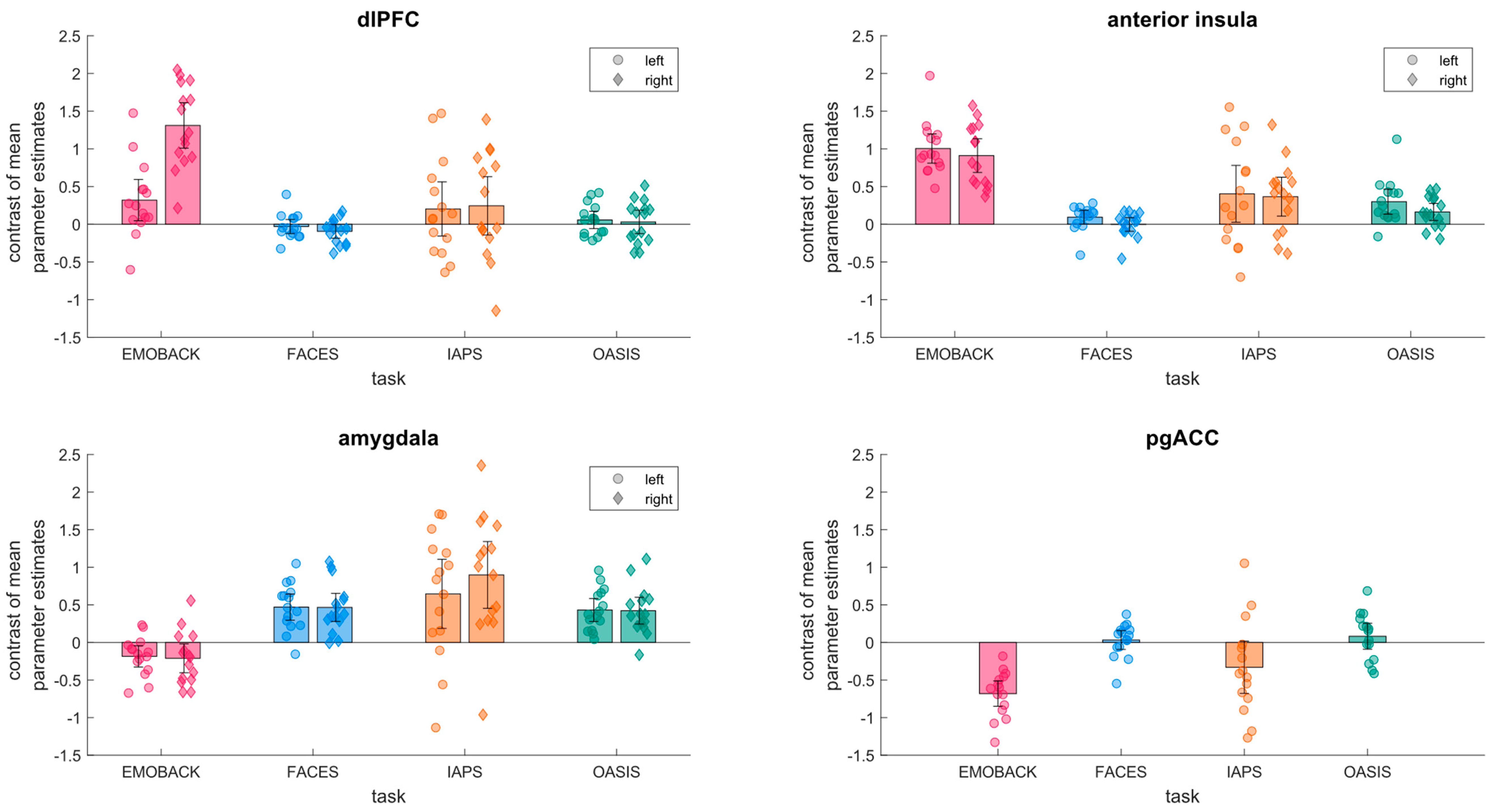
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García-García, I.; Kube, J.; Gaebler, M.; Horstmann, A.; Villringer, A.; Neumann, J. Neural Processing of Negative Emotional Stimuli and the Influence of Age, Sex and Task-Related Characteristics. Neurosci. Biobehav. Rev. 2016, 68, 773–793. [Google Scholar] [CrossRef]
- Wu, M.; Kujawa, A.; Lu, L.H.; Fitzgerald, D.A.; Klumpp, H.; Fitzgerald, K.D.; Monk, C.S.; Phan, K.L. Age-Related Changes in Amygdala-Frontal Connectivity during Emotional Face Processing from Childhood into Young Adulthood. Hum. Brain Mapp. 2016, 37, 1684–1695. [Google Scholar] [CrossRef] [PubMed]
- McTeague, L.M.; Rosenberg, B.; Lopez, J.M.; Carreon, D.; Huemer, J.; Jiang, Y.; Chick, C.; Eickhoff, S.; Etkin, A. Identification of Common Neural Circuit Disruptions in Emotional Processing Across Psychiatric Disorders. Am. J. Psychiatry 2020, 177, 411–421. [Google Scholar] [CrossRef]
- Outhred, T.; Das, P.; Felmingham, K.; Bryant, R.; Nathan, P.; Malhi, G.; Kemp, A. Impact of Acute Administration of Escitalopram on the Processing of Emotional and Neutral Images: A Randomized Crossover FMRI Study of Healthy Women. J. Psychiatry Neurosci. JPN 2014, 39, 267–275. [Google Scholar] [CrossRef]
- Enneking, V.; Dzvonyar, F.; Dück, K.; Dohm, K.; Grotegerd, D.; Förster, K.; Meinert, S.; Lemke, H.; Klug, M.; Waltemate, L.; et al. Brain Functional Effects of Electroconvulsive Therapy during Emotional Processing in Major Depressive Disorder. Brain Stimulat. 2020, 13, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Sabatinelli, D.; Fortune, E.E.; Li, Q.; Siddiqui, A.; Krafft, C.; Oliver, W.T.; Beck, S.; Jeffries, J. Emotional Perception: Meta-Analyses of Face and Natural Scene Processing. Neuroimage 2011, 54, 2524–2533. [Google Scholar] [CrossRef] [PubMed]
- Müller, V.I.; Höhner, Y.; Eickhoff, S.B. Influence of Task Instructions and Stimuli on the Neural Network of Face Processing: An ALE Meta-Analysis. Cortex 2018, 103, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Reisch, L.M.; Wegrzyn, M.; Kissler, J.; Woermann, F.G.; Bien, C.G. Negative Content Enhances Stimulus-Specific Cerebral Activity during Free Viewing of Pictures, Faces, and Words. Hum. Brain Mapp. 2020, 41, 4332–4354. [Google Scholar] [CrossRef]
- Riedel, M.C.; Yanes, J.A.; Ray, K.L.; Eickhoff, S.B.; Fox, P.T.; Sutherland, M.T.; Laird, A.R. Dissociable Meta-analytic Brain Networks Contribute to Coordinated Emotional Processing. Hum. Brain Mapp. 2018, 39, 2514–2531. [Google Scholar] [CrossRef]
- Villalta-Gil, V.; Hinton, K.E.; Landman, B.A.; Yvernault, B.C.; Perkins, S.F.; Katsantonis, A.S.; Sellani, C.L.; Lahey, B.B.; Zald, D.H. Convergent Individual Differences in Visual Cortices, but Not the Amygdala across Standard Amygdalar FMRI Probe Tasks. Neuroimage 2017, 146, 312–319. [Google Scholar] [CrossRef]
- Lindquist, K.A.; Wager, T.D.; Kober, H.; Bliss-Moreau, E.; Barrett, L.F. The Brain Basis of Emotion: A Meta-Analytic Review. Behav. Brain Sci. 2012, 35, 121–143. [Google Scholar] [CrossRef]
- Whalen, P.; Davis, F.C.; Oler, J.A.; Kim, H.; Kim, M.J.; Neta, M. Human amygdala responses to facial expressions of emotion. In The Human Amygdala; The Guilford Press: New York, NY, USA, 2009; pp. 265–288. ISBN 978-1-60623-033-6. [Google Scholar]
- Stuhrmann, A.; Suslow, T.; Dannlowski, U. Facial Emotion Processing in Major Depression: A Systematic Review of Neuroimaging Findings. Biol. Mood Anxiety Disord. 2011, 1, 10. [Google Scholar] [CrossRef] [PubMed]
- Gentili, C.; Cristea, I.A.; Angstadt, M.; Klumpp, H.; Tozzi, L.; Phan, K.L.; Pietrini, P. Beyond Emotions: A Meta-Analysis of Neural Response within Face Processing System in Social Anxiety. Exp. Biol. Med. 2016, 241, 225–237. [Google Scholar] [CrossRef]
- Craig, A.D. How Do You Feel? Interoception: The Sense of the Physiological Condition of the Body. Nat. Rev. Neurosci. 2002, 3, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Zaki, J.; Davis, J.I.; Ochsner, K.N. Overlapping Activity in Anterior Insula during Interoception and Emotional Experience. NeuroImage 2012, 62, 493–499. [Google Scholar] [CrossRef]
- Duerden, E.G.; Arsalidou, M.; Lee, M.; Taylor, M.J. Lateralization of Affective Processing in the Insula. NeuroImage 2013, 78, 159–175. [Google Scholar] [CrossRef]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable Intrinsic Connectivity Networks for Salience Processing and Executive Control. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef]
- Raichle, M.E. The Brain’s Default Mode Network. Annu. Rev. Neurosci. 2015, 38, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Palomero-Gallagher, N.; Hoffstaedter, F.; Mohlberg, H.; Eickhoff, S.B.; Amunts, K.; Zilles, K. Human Pregenual Anterior Cingulate Cortex: Structural, Functional, and Connectional Heterogeneity. Cereb. Cortex 2019, 29, 2552–2574. [Google Scholar] [CrossRef] [PubMed]
- Vogt, B.A. Submodalities of Emotion in the Context of Cingulate Subregions. Cortex J. Devoted Study Nerv. Syst. Behav. 2014, 59, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Phan, K.L.; Wager, T.; Taylor, S.F.; Liberzon, I. Functional Neuroanatomy of Emotion: A Meta-Analysis of Emotion Activation Studies in PET and FMRI. NeuroImage 2002, 16, 331–348. [Google Scholar] [CrossRef]
- Tang, W.; Jbabdi, S.; Zhu, Z.; Cottaar, M.; Grisot, G.; Lehman, J.F.; Yendiki, A.; Haber, S.N. A Connectional Hub in the Rostral Anterior Cingulate Cortex Links Areas of Emotion and Cognitive Control. eLife 2019, 8. [Google Scholar] [CrossRef]
- Ochsner, K.N.; Silvers, J.A.; Buhle, J.T. Functional Imaging Studies of Emotion Regulation: A Synthetic Review and Evolving Model of the Cognitive Control of Emotion. Ann. N. Y. Acad. Sci. 2012, 1251, E1. [Google Scholar] [CrossRef]
- Schweizer, S.; Satpute, A.B.; Atzil, S.; Field, A.P.; Hitchcock, C.; Black, M.; Barrett, L.F.; Dalgleish, T. The Impact of Affective Information on Working Memory: A Pair of Meta-Analytic Reviews of Behavioral and Neuroimaging Evidence. Psychol. Bull. 2019, 145, 566–609. [Google Scholar] [CrossRef]
- Van Dillen, L.F.; Heslenfeld, D.J.; Koole, S.L. Tuning down the Emotional Brain: An FMRI Study of the Effects of Cognitive Load on the Processing of Affective Images. NeuroImage 2009, 45, 1212–1219. [Google Scholar] [CrossRef]
- Kohn, N.; Eickhoff, S.B.; Scheller, M.; Laird, A.R.; Fox, P.T.; Habel, U. Neural Network of Cognitive Emotion Regulation—An ALE Meta-Analysis and MACM Analysis. Neuroimage 2014, 87, 345–355. [Google Scholar] [CrossRef]
- Grimm, S.; Weigand, A.; Kazzer, P.; Jacobs, A.M.; Bajbouj, M. Neural Mechanisms Underlying the Integration of Emotion and Working Memory. Neuroimage 2012, 61, 1188–1194. [Google Scholar] [CrossRef]
- Võ, M.L.H.; Conrad, M.; Kuchinke, L.; Urton, K.; Hofmann, M.J.; Jacobs, A.M. The Berlin Affective Word List Reloaded (BAWL-R). Behav. Res. Methods 2009, 41, 534–538. [Google Scholar] [CrossRef]
- Olszanowski, M.; Pochwatko, G.; Kuklinski, K.; Scibor-Rylski, M.; Lewinski, P.; Ohme, R.K. Warsaw Set of Emotional Facial Expression Pictures: A Validation Study of Facial Display Photographs. Front. Psychol. 2015, 5, 1516. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.J.; Bradley, M.M.; Cuthbert, B.N. International Affective Picture System (IAPS): Technical Manual and Affective Ratings. NIMH Cent. Study Emot. Atten. 1997, 1, 39–58. [Google Scholar]
- Kurdi, B.; Lozano, S.; Banaji, M.R. Introducing the Open Affective Standardized Image Set (OASIS). Behav. Res. Methods 2017, 49, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Price, C.J.; Veltman, D.J.; Ashburner, J.; Josephs, O.; Friston, K.J. The Critical Relationship between the Timing of Stimulus Presentation and Data Acquisition in Blocked Designs with FMRI. Neuroimage 1999, 10, 36–44. [Google Scholar] [CrossRef][Green Version]
- Friston, K.J.; Holmes, A.P.; Price, C.J.; Büchel, C.; Worsley, K.J. Multisubject FMRI Studies and Conjunction Analyses. Neuroimage 1999, 10, 385–396. [Google Scholar] [CrossRef]
- Wang, H.; He, W.; Wu, J.; Zhang, J.; Jin, Z.; Li, L. A Coordinate-Based Meta-Analysis of the n-Back Working Memory Paradigm Using Activation Likelihood Estimation. Brain Cogn. 2019, 132, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Erk, S.; Kleczar, A.; Walter, H. Valence-Specific Regulation Effects in a Working Memory Task with Emotional Context. Neuroimage 2007, 37, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Costafreda, S.G.; Brammer, M.J.; David, A.S.; Fu, C.H.Y. Predictors of Amygdala Activation during the Processing of Emotional Stimuli: A Meta-Analysis of 385 PET and FMRI Studies. Brain Res. Rev. 2008, 58, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Schlochtermeier, L.H.; Kuchinke, L.; Pehrs, C.; Urton, K.; Kappelhoff, H.; Jacobs, A.M. Emotional Picture and Word Processing: An FMRI Study on Effects of Stimulus Complexity. PLoS ONE 2013, 8, e55619. [Google Scholar] [CrossRef] [PubMed]
- Spreng, R.N. The Fallacy of a “Task-Negative” Network. Front. Psychol. 2012, 3, 145. [Google Scholar] [CrossRef]
- Lindquist, K.A.; Satpute, A.B.; Wager, T.D.; Weber, J.; Barrett, L.F. The Brain Basis of Positive and Negative Affect: Evidence from a Meta-Analysis of the Human Neuroimaging Literature. Cereb. Cortex 2016, 26, 1910–1922. [Google Scholar] [CrossRef]
- Wager, T.D.; Phan, K.L.; Liberzon, I.; Taylor, S.F. Valence, Gender, and Lateralization of Functional Brain Anatomy in Emotion: A Meta-Analysis of Findings from Neuroimaging. Neuroimage 2003, 19, 513–531. [Google Scholar] [CrossRef]
- Aldhafeeri, F.M.; Mackenzie, I.; Kay, T.; Alghamdi, J.; Sluming, V. Regional Brain Responses to Pleasant and Unpleasant IAPS Pictures: Different Networks. Neurosci. Lett. 2012, 512, 94–98. [Google Scholar] [CrossRef]
- Britton, J.C.; Taylor, S.F.; Sudheimer, K.D.; Liberzon, I. Facial Expressions and Complex IAPS Pictures: Common and Differential Networks. Neuroimage 2006, 31, 906–919. [Google Scholar] [CrossRef]
- Korgaonkar, M.S.; Grieve, S.M.; Etkin, A.; Koslow, S.H.; Williams, L.M. Using Standardized FMRI Protocols to Identify Patterns of Prefrontal Circuit Dysregulation That Are Common and Specific to Cognitive and Emotional Tasks in Major Depressive Disorder: First Wave Results from the ISPOT-D Study. Neuropsychopharmacology 2013, 38, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.L.; Chase, H.W.; Sheline, Y.I.; Etkin, A.; Almeida, J.R.; Deckersbach, T.; Trivedi, M.H. Identifying Predictors, Moderators, and Mediators of Antidepressant Response in Major Depressive Disorder: Neuroimaging Approaches. Am. J. Psychiatry 2015, 172, 124–138. [Google Scholar] [CrossRef]
- Botvinik-Nezer, R.; Holzmeister, F.; Camerer, C.F.; Dreber, A.; Huber, J.; Johannesson, M.; Kirchler, M.; Iwanir, R.; Mumford, J.A.; Adcock, R.A.; et al. Variability in the Analysis of a Single Neuroimaging Dataset by Many Teams. Nature 2020, 582, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Poldrack, R.A.; Baker, C.I.; Durnez, J.; Gorgolewski, K.J.; Matthews, P.M.; Munafò, M.R.; Nichols, T.E.; Poline, J.B.; Vul, E.; Yarkoni, T. Scanning the Horizon: Towards Transparent and Reproducible Neuroimaging Research. Nat. Rev. Neurosci. 2017, 18, 115–126. [Google Scholar] [CrossRef]
- Elliott, M.L.; Knodt, A.R.; Ireland, D.; Morris, M.L.; Poulton, R.; Ramrakha, S.; Sison, M.L.; Moffitt, T.E.; Caspi, A.; Hariri, A.R. What Is the Test-Retest Reliability of Common Task-Functional MRI Measures? New Empirical Evidence and a Meta-Analysis. Psychol. Sci. 2020, 31, 792–806. [Google Scholar] [CrossRef]
- Frohner, J.H.; Smolka, M.N.; Kroemer, N.B.; Teckentrup, V. Addressing the Reliability Fallacy in FMRI: Similar Group Effects May Arise from Unreliable Individual Effects. Neuroimage 2019, 222, 174–189. [Google Scholar] [CrossRef] [PubMed]
- Nee, D.E. FMRI Replicability Depends upon Sufficient Individual-Level Data. Commun. Biol. 2019, 2, 1–4. [Google Scholar] [CrossRef]
- Jovicich, J.; Czanner, S.; Han, X.; Salat, D.; van der Kouwe, A.; Quinn, B.; Pacheco, J.; Albert, M.; Killiany, R.; Blacker, D.; et al. MRI-Derived Measurements of Human Subcortical, Ventricular and Intracranial Brain Volumes: Reliability Effects of Scan Sessions, Acquisition Sequences, Data Analyses, Scanner Upgrade, Scanner Vendors and Field Strengths. Neuroimage 2009, 46, 177–192. [Google Scholar] [CrossRef]
- Gee, D.G.; McEwen, S.C.; Forsyth, J.K.; Haut, K.M.; Bearden, C.E.; Addington, J.; Goodyear, B.; Cadenhead, K.S.; Mirzakhanian, H.; Cornblatt, B.A.; et al. Reliability of an FMRI Paradigm for Emotional Processing in a Multisite Longitudinal Study. Hum. Brain Mapp. 2015, 36, 2558–2579. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, J.K.; McEwen, S.C.; Gee, D.G.; Bearden, C.E.; Addington, J.; Goodyear, B.; Cadenhead, K.S.; Mirzakhanian, H.; Cornblatt, B.A.; Olvet, D.M. Reliability of Functional Magnetic Resonance Imaging Activation during Working Memory in a Multi-Site Study: Analysis from the North American Prodrome Longitudinal Study. Neuroimage 2014, 97, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Esteban, O.; Markiewicz, C.J.; Blair, R.W.; Moodie, C.A.; Isik, A.I.; Erramuzpe, A.; Kent, J.D.; Goncalves, M.; DuPre, E.; Snyder, M. FMRIPrep: A Robust Preprocessing Pipeline for Functional MRI. Nat. Methods 2019, 16, 111–116. [Google Scholar] [CrossRef]
- Cacioppo, J.T.; Berntson, G.G.; Bechara, A.; Tranel, D.; Hawkley, L.C. Could an aging brain contribute to subjective well-being? The value added by a social neuroscience perspective. In Social Neuroscience: Toward Understanding the Underpinnings of the Social Mind; Oxford Series in Social Cognition and Social Neuroscience; Oxford University Press: New York, NY, USA, 2011; pp. 249–262. ISBN 978-0-19-531687-2. [Google Scholar]
- MacCormack, J.K.; Stein, A.G.; Kang, J.; Giovanello, K.S.; Satpute, A.B.; Lindquist, K.A. Affect in the Aging Brain: A Neuroimaging Meta-Analysis of Older Vs. Younger Adult Affective Experience and Perception. Affect. Sci. 2020, 1, 128–154. [Google Scholar] [CrossRef]
- Bluhm, R. New Research, Old Problems: Methodological and Ethical Issues in FMRI Research Examining Sex/Gender Differences in Emotion Processing. Neuroethics 2013, 6, 319–330. [Google Scholar] [CrossRef]
- Wrase, J.; Klein, S.; Gruesser, S.M.; Hermann, D.; Flor, H.; Mann, K.; Braus, D.F.; Heinz, A. Gender Differences in the Processing of Standardized Emotional Visual Stimuli in Humans: A Functional Magnetic Resonance Imaging Study. Neurosci. Lett. 2003, 348, 41–45. [Google Scholar] [CrossRef]
- Stevens, J.S.; Hamann, S. Sex Differences in Brain Activation to Emotional Stimuli: A Meta-Analysis of Neuroimaging Studies. Neuropsychologia 2012, 50, 1578–1593. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hartling, C.; Metz, S.; Pehrs, C.; Scheidegger, M.; Gruzman, R.; Keicher, C.; Wunder, A.; Weigand, A.; Grimm, S. Comparison of Four fMRI Paradigms Probing Emotion Processing. Brain Sci. 2021, 11, 525. https://doi.org/10.3390/brainsci11050525
Hartling C, Metz S, Pehrs C, Scheidegger M, Gruzman R, Keicher C, Wunder A, Weigand A, Grimm S. Comparison of Four fMRI Paradigms Probing Emotion Processing. Brain Sciences. 2021; 11(5):525. https://doi.org/10.3390/brainsci11050525
Chicago/Turabian StyleHartling, Corinna, Sophie Metz, Corinna Pehrs, Milan Scheidegger, Rebecca Gruzman, Christian Keicher, Andreas Wunder, Anne Weigand, and Simone Grimm. 2021. "Comparison of Four fMRI Paradigms Probing Emotion Processing" Brain Sciences 11, no. 5: 525. https://doi.org/10.3390/brainsci11050525
APA StyleHartling, C., Metz, S., Pehrs, C., Scheidegger, M., Gruzman, R., Keicher, C., Wunder, A., Weigand, A., & Grimm, S. (2021). Comparison of Four fMRI Paradigms Probing Emotion Processing. Brain Sciences, 11(5), 525. https://doi.org/10.3390/brainsci11050525