
Contribution of TMS and TMS-EEG to the Understanding of Mechanisms Underlying Physiological Brain Aging


 and
and 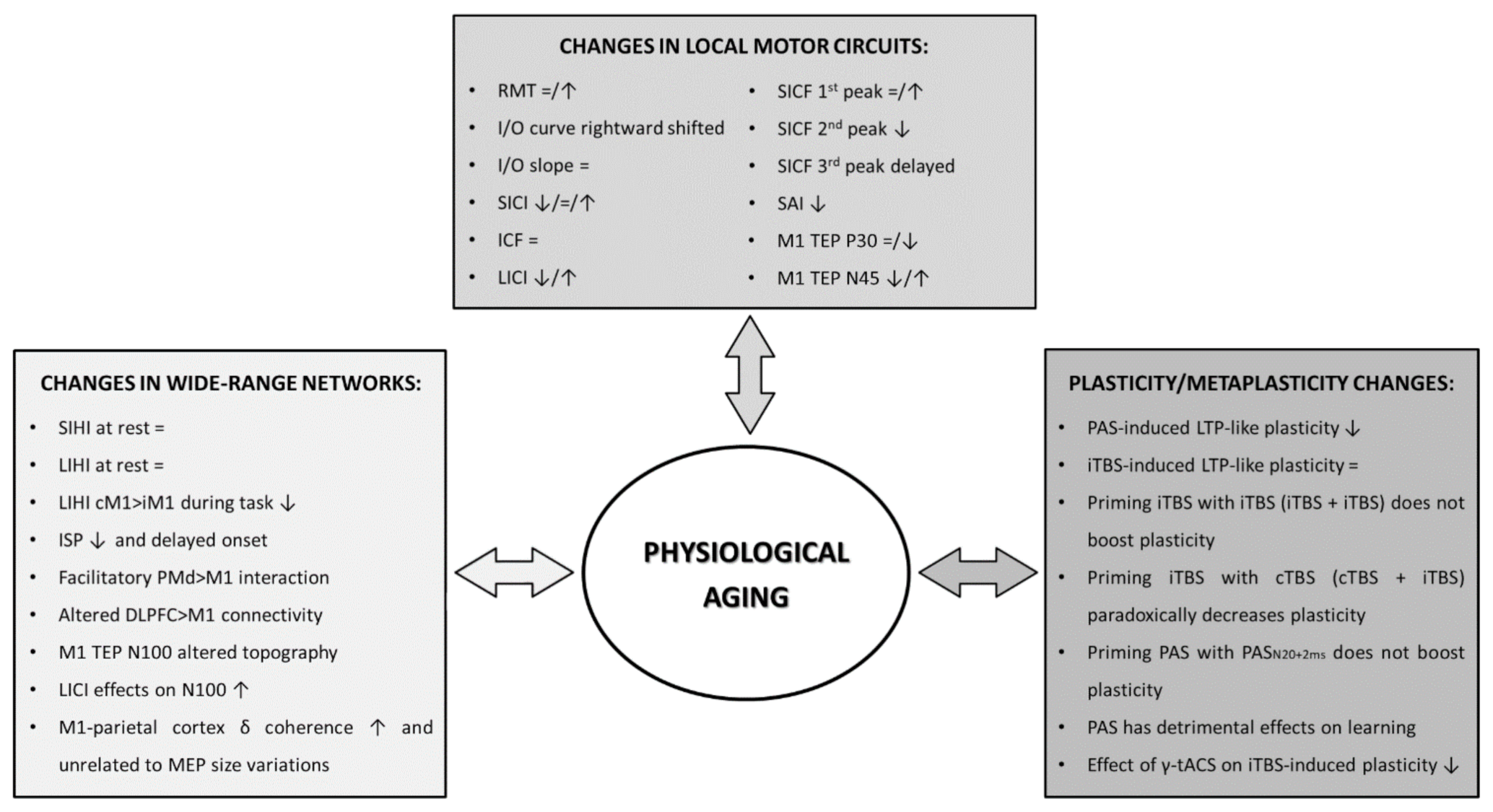{kind=link}
Abstract
1. Introduction
2. Neurophysiological Changes in Local Motor Circuits during Aging
2.1. TMS Studies
2.2. TMS-EEG Studies
3. Neurophysiological Changes in Wide-Range Networks during Aging
3.1. TMS Studies
3.2. TMS-EEG Studies
4. Neurophysiological Changes in Plasticity and Metaplasticity Processes during Aging
5. Limitations, Perspectives and Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Burke, S.N.; Barnes, C.A. Neural plasticity in the ageing brain. Nat. Rev. Neurosci. 2006, 7, 30–40. [Google Scholar] [CrossRef]
- Yeoman, M.; Scutt, G.; Faragher, R. Insights into CNS ageing from animal models of senescence. Nat. Rev. Neurosci. 2012, 13, 435–445. [Google Scholar] [CrossRef]
- Enzinger, C.; Fazekas, F.; Matthews, P.M.; Ropele, S.; Schmidt, H.; Smith, S.; Schmidt, R. Risk factors for progression of brain atrophy in aging: Six-year follow-up of normal subjects. Neurology 2005, 64, 1704–1711. [Google Scholar] [CrossRef]
- Rossini, P.M.; Ferilli, M.A.N.; Rossini, L.; Ferreri, F. Clinical neurophysiology of brain plasticity in aging brain. Curr. Pharm. Des. 2013, 19, 6426–6439. [Google Scholar] [CrossRef] [PubMed]
- Doherty, T.J. Invited review: Aging and sarcopenia. J. Appl. Physiol. (1985) 2003, 95, 1717–1727. [Google Scholar] [CrossRef] [PubMed]
- Semmler, J.G.; Kornatz, K.W.; Meyer, F.G.; Enoka, R.M. Diminished task-related adjustments of common inputs to hand muscle motor neurons in older adults. Exp. Brain Res. 2006, 172, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, J.A.; Larkin, L.M.; Claflin, D.R.; Brooks, S.V. Age-related changes in the structure and function of skeletal muscles. Clin. Exp. Pharmacol. Physiol. 2007, 34, 1091–1096. [Google Scholar] [CrossRef]
- Bhandari, A.; Radhu, N.; Farzan, F.; Mulsant, B.H.; Rajji, T.K.; Daskalakis, Z.J.; Blumberger, D.M. A meta-analysis of the effects of aging on motor cortex neurophysiology assessed by transcranial magnetic stimulation. Clin. Neurophysiol. 2016, 127, 2834–2845. [Google Scholar] [CrossRef] [PubMed]
- Damoiseaux, J.S. Effects of aging on functional and structural brain connectivity. Neuroimage 2017, 160, 32–40. [Google Scholar] [CrossRef]
- Ferreri, F.; Pauri, F.; Pasqualetti, P.; Fini, R.; Dal Forno, G.; Rossini, P.M. Motor cortex excitability in Alzheimer’s disease: A transcranial magnetic stimulation study. Ann. Neurol. 2003, 53, 102–108. [Google Scholar] [CrossRef]
- Ferreri, F.; Pasqualetti, P.; Määttä, S.; Ponzo, D.; Guerra, A.; Bressi, F.; Chiovenda, P.; Del Duca, M.; Giambattistelli, F.; Ursini, F.; et al. Motor cortex excitability in Alzheimer’s disease: A transcranial magnetic stimulation follow-up study. Neurosci. Lett. 2011, 492, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Vecchio, F.; Vollero, L.; Guerra, A.; Petrichella, S.; Ponzo, D.; Määtta, S.; Mervaala, E.; Könönen, M.; Ursini, F.; et al. Sensorimotor cortex excitability and connectivity in Alzheimer’s disease: A TMS-EEG Co-registration study. Hum. Brain Mapp. 2016, 37, 2083–2096. [Google Scholar] [CrossRef]
- Bologna, M.; Guerra, A.; Paparella, G.; Giordo, L.; Alunni Fegatelli, D.; Vestri, A.R.; Rothwell, J.C.; Berardelli, A. Neurophysiological correlates of bradykinesia in Parkinson’s disease. Brain 2018, 141, 2432–2444. [Google Scholar] [CrossRef] [PubMed]
- Bologna, M.; Guerra, A.; Colella, D.; Cioffi, E.; Paparella, G.; Di Vita, A.; D’Antonio, F.; Trebbastoni, A.; Berardelli, A. Bradykinesia in Alzheimer’s disease and its neurophysiological substrates. Clin. Neurophysiol. 2020, 131, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.; Costantini, E.M.; Maatta, S.; Ponzo, D.; Ferreri, F. Disorders of consciousness and electrophysiological treatment strategies: A review of the literature and new perspectives. Curr. Pharm. Des. 2014, 20, 4248–4267. [Google Scholar] [CrossRef]
- Guerra, A.; Petrichella, S.; Vollero, L.; Ponzo, D.; Pasqualetti, P.; Määttä, S.; Mervaala, E.; Könönen, M.; Bressi, F.; Iannello, G.; et al. Neurophysiological features of motor cortex excitability and plasticity in Subcortical Ischemic Vascular Dementia: A TMS mapping study. Clin. Neurophysiol. 2015, 126, 906–913. [Google Scholar] [CrossRef]
- Guerra, A.; Suppa, A.; D’Onofrio, V.; Di Stasio, F.; Asci, F.; Fabbrini, G.; Berardelli, A. Abnormal cortical facilitation and L-dopa-induced dyskinesia in Parkinson’s disease. Brain Stimul. 2019, 12, 1517–1525. [Google Scholar] [CrossRef]
- Guerra, A.; Asci, F.; D’Onofrio, V.; Sveva, V.; Bologna, M.; Fabbrini, G.; Berardelli, A.; Suppa, A. Enhancing Gamma Oscillations Restores Primary Motor Cortex Plasticity in Parkinson’s Disease. J. Neurosci. 2020, 40, 4788–4796. [Google Scholar] [CrossRef]
- Benussi, A.; Alberici, A.; Ferrari, C.; Cantoni, V.; Dell’Era, V.; Turrone, R.; Cotelli, M.S.; Binetti, G.; Paghera, B.; Koch, G.; et al. The impact of transcranial magnetic stimulation on diagnostic confidence in patients with Alzheimer disease. Alzheimer’s Res. Ther. 2018, 10, 94. [Google Scholar] [CrossRef]
- Colella, D.; Guerra, A.; Paparella, G.; Cioffi, E.; Di Vita, A.; Trebbastoni, A.; Berardelli, A.; Bologna, M. Motor dysfunction in mild cognitive impairment as tested by kinematic analysis and transcranial magnetic stimulation. Clin. Neurophysiol. 2021, 132, 315–322. [Google Scholar] [CrossRef]
- Hill, A.T.; Rogasch, N.C.; Fitzgerald, P.B.; Hoy, K.E. TMS-EEG: A window into the neurophysiological effects of transcranial electrical stimulation in non-motor brain regions. Neurosci. Biobehav. Rev. 2016, 64, 175–184. [Google Scholar] [CrossRef]
- Rawji, V.; Latorre, A.; Sharma, N.; Rothwell, J.C.; Rocchi, L. On the Use of TMS to Investigate the Pathophysiology of Neurodegenerative Diseases. Front. Neurol. 2020, 11, 584664. [Google Scholar] [CrossRef] [PubMed]
- Cardin, J.A.; Abel, T. Memory suppressor genes: Enhancing the relationship between synaptic plasticity and memory storage. J. Neurosci. Res. 1999, 58, 10–23. [Google Scholar] [CrossRef]
- Rosenzweig, E.S.; Barnes, C.A. Impact of aging on hippocampal function: Plasticity, network dynamics, and cognition. Prog. Neurobiol. 2003, 69, 143–179. [Google Scholar] [CrossRef]
- Disterhoft, J.F.; Oh, M.M. Learning, aging and intrinsic neuronal plasticity. Trends Neurosci. 2006, 29, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, L.; Chalavi, S.; Swinnen, S.P. Aging and brain plasticity. Aging 2018, 10, 1789–1790. [Google Scholar] [CrossRef] [PubMed]
- Abraham, W.C.; Richter-Levin, G. From Synaptic Metaplasticity to Behavioral Metaplasticity. Neurobiol. Learn. Mem. 2018, 154, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Dunn, A.R.; Kaczorowski, C.C. Regulation of intrinsic excitability: Roles for learning and memory, aging and Alzheimer’s disease, and genetic diversity. Neurobiol. Learn. Mem. 2019, 164, 107069. [Google Scholar] [CrossRef]
- Amassian, V.E.; Stewart, M.; Quirk, G.J.; Rosenthal, J.L. Physiological basis of motor effects of a transient stimulus to cerebral cortex. Neurosurgery 1987, 20, 74–93. [Google Scholar] [CrossRef] [PubMed]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.G.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.B.; George, M.S.; et al. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application. An updated report from an I.F.C.N. Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, U. I-waves in motor cortex revisited. Exp. Brain Res. 2020, 238, 1601–1610. [Google Scholar] [CrossRef]
- Guerra, A.; Ranieri, F.; Falato, E.; Musumeci, G.; Di Santo, A.; Asci, F.; Di Pino, G.; Suppa, A.; Berardelli, A.; Di Lazzaro, V. Detecting cortical circuits resonant to high-frequency oscillations in the human primary motor cortex: A TMS-tACS study. Sci. Rep. 2020, 10, 7695. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Rothwell, J.; Capogna, M. Noninvasive Stimulation of the Human Brain: Activation of Multiple Cortical Circuits. Neuroscientist 2017, 24, 246–260. [Google Scholar] [CrossRef]
- Brown, K.E.; Lohse, K.R.; Mayer, I.M.S.; Strigaro, G.; Desikan, M.; Casula, E.P.; Meunier, S.; Popa, T.; Lamy, J.-C.; Odish, O.; et al. The reliability of commonly used electrophysiology measures. Brain Stimul. 2017, 10, 1102–1111. [Google Scholar] [CrossRef]
- Davila-Pérez, P.; Jannati, A.; Fried, P.J.; Cudeiro Mazaira, J.; Pascual-Leone, A. The Effects of Waveform and Current Direction on the Efficacy and Test-Retest Reliability of Transcranial Magnetic Stimulation. Neuroscience 2018, 393, 97–109. [Google Scholar] [CrossRef]
- Gomes-Osman, J.; Indahlastari, A.; Fried, P.J.; Cabral, D.L.F.; Rice, J.; Nissim, N.R.; Aksu, S.; McLaren, M.E.; Woods, A.J. Non-invasive Brain Stimulation: Probing Intracortical Circuits and Improving Cognition in the Aging Brain. Front. Aging Neurosci. 2018, 10, 177. [Google Scholar] [CrossRef] [PubMed]
- Kozel, F.A.; Nahas, Z.; deBrux, C.; Molloy, M.; Lorberbaum, J.P.; Bohning, D.; Risch, S.C.; George, M.S. How coil-cortex distance relates to age, motor threshold, and antidepressant response to repetitive transcranial magnetic stimulation. J. Neuropsychiatry Clin. Neurosci. 2000, 12, 376–384. [Google Scholar] [CrossRef]
- List, J.; Kübke, J.C.; Lindenberg, R.; Külzow, N.; Kerti, L.; Witte, V.; Flöel, A. Relationship between excitability, plasticity and thickness of the motor cortex in older adults. Neuroimage 2013, 83, 809–816. [Google Scholar] [CrossRef] [PubMed]
- McGinley, M.; Hoffman, R.L.; Russ, D.W.; Thomas, J.S.; Clark, B.C. Older adults exhibit more intracortical inhibition and less intracortical facilitation than young adults. Exp. Gerontol. 2010, 45, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Young-Bernier, M.; Davidson, P.S.R.; Tremblay, F. Paired-pulse afferent modulation of TMS responses reveals a selective decrease in short latency afferent inhibition with age. Neurobiol. Aging 2012, 33, 835.e1–835.e11. [Google Scholar] [CrossRef]
- Peinemann, A.; Lehner, C.; Conrad, B.; Siebner, H.R. Age-related decrease in paired-pulse intracortical inhibition in the human primary motor cortex. Neurosci Lett. 2001, 313, 33–36. [Google Scholar] [CrossRef]
- Oliviero, A.; Profice, P.; Tonali, P.A.; Pilato, F.; Saturno, E.; Dileone, M.; Ranieri, F.; Di Lazzaro, V. Effects of aging on motor cortex excitability. Neurosci. Res. 2006, 55, 74–77. [Google Scholar] [CrossRef]
- Opie, G.M.; Semmler, J.G. Age-related differences in short- and long-interval intracortical inhibition in a human hand muscle. Brain Stimul. 2014, 7, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, K.; Park, S.B.; Geevasinga, N.; Huynh, W.; Simon, N.G.; Menon, P.; Howells, J.; Vucic, S.; Kiernan, M.C. Threshold tracking transcranial magnetic stimulation: Effects of age and gender on motor cortical function. Clin. Neurophysiol. 2016, 127, 2355–2361. [Google Scholar] [CrossRef] [PubMed]
- Hess, C.W.; Mills, K.R.; Murray, N.M. Responses in small hand muscles from magnetic stimulation of the human brain. J. Physiol. 1987, 388, 397–419. [Google Scholar] [CrossRef]
- Bologna, M.; Rocchi, L.; Paparella, G.; Nardella, A.; Li Voti, P.; Conte, A.; Kojovic, M.; Rothwell, J.C.; Berardelli, A. Reversal of Practice-related Effects on Corticospinal Excitability has no Immediate Effect on Behavioral Outcome. Brain Stimul. 2015, 8, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, J.B.; Ogston, K.M.; Miles, T.S. Age and sex differences in human motor cortex input-output characteristics. J. Physiol. 2003, 546, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, J.B.; Doeltgen, S.H.; Goldsworthy, M.R.; Schneider, L.A.; Vallence, A.-M.; Smith, A.E.; Semmler, J.G.; McDonnell, M.N.; Ridding, M.C. A comparison of two methods for estimating 50% of the maximal motor evoked potential. Clin. Neurophysiol. 2015, 126, 2337–2341. [Google Scholar] [CrossRef] [PubMed]
- Kujirai, T.; Caramia, M.D.; Rothwell, J.C.; Day, B.L.; Thompson, P.D.; Ferbert, A.; Wroe, S.; Asselman, P.; Marsden, C.D. Corticocortical inhibition in human motor cortex. J. Physiol. 1993, 471, 501–519. [Google Scholar] [CrossRef]
- Ganos, C.; Rocchi, L.; Latorre, A.; Hockey, L.; Palmer, C.; Joyce, E.M.; Bhatia, K.P.; Haggard, P.; Rothwell, J. Motor cortical excitability during voluntary inhibition of involuntary tic movements. Mov. Disord. 2018, 33, 1804–1809. [Google Scholar] [CrossRef] [PubMed]
- Tokimura, H.; Di Lazzaro, V.; Tokimura, Y.; Oliviero, A.; Profice, P.; Insola, A.; Mazzone, P.; Tonali, P.; Rothwell, J.C. Short latency inhibition of human hand motor cortex by somatosensory input from the hand. The Journal of Physiology 2000, 523, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, J.; Hannah, R.; Rocchi, L.; Rothwell, J.C. Premovement Suppression of Corticospinal Excitability may be a Necessary Part of Movement Preparation. Cereb. Cortex 2020, 30, 2910–2923. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, U.; Reis, J.; Schwenkreis, P.; Rosanova, M.; Strafella, A.; Badawy, R.; Müller-Dahlhaus, F. TMS and drugs revisited 2014. Clin. Neurophysiol. 2015, 126, 1847–1868. [Google Scholar] [CrossRef]
- Rocchi, L.; Latorre, A.; Ibanez Pereda, J.; Spampinato, D.; Brown, K.E.; Rothwell, J.; Bhatia, K. A case of congenital hypoplasia of the left cerebellar hemisphere and ipsilateral cortical myoclonus. Mov. Disord. 2019, 34, 1745–1747. [Google Scholar] [CrossRef]
- Paparella, G.; Rocchi, L.; Bologna, M.; Berardelli, A.; Rothwell, J. Differential effects of motor skill acquisition on the primary motor and sensory cortices in healthy humans. J. Physiol. 2020, 598, 4031–4045. [Google Scholar] [CrossRef]
- Marneweck, M.; Loftus, A.; Hammond, G. Short-interval intracortical inhibition and manual dexterity in healthy aging. Neurosci. Res. 2011, 70, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Heise, K.-F.; Zimerman, M.; Hoppe, J.; Gerloff, C.; Wegscheider, K.; Hummel, F.C. The aging motor system as a model for plastic changes of GABA-mediated intracortical inhibition and their behavioral relevance. J. Neurosci. 2013, 33, 9039–9049. [Google Scholar] [CrossRef]
- Hermans, L.; Levin, O.; Maes, C.; van Ruitenbeek, P.; Heise, K.-F.; Edden, R.A.E.; Puts, N.A.J.; Peeters, R.; King, B.R.; Meesen, R.L.J.; et al. GABA levels and measures of intracortical and interhemispheric excitability in healthy young and older adults: An MRS-TMS study. Neurobiol. Aging 2018, 65, 168–177. [Google Scholar] [CrossRef]
- Cuypers, K.; Verstraelen, S.; Maes, C.; Hermans, L.; Hehl, M.; Heise, K.-F.; Chalavi, S.; Mikkelsen, M.; Edden, R.; Levin, O.; et al. Task-related measures of short-interval intracortical inhibition and GABA levels in healthy young and older adults: A multimodal TMS-MRS study. Neuroimage 2020, 208, 116470. [Google Scholar] [CrossRef]
- Rogasch, N.C.; Dartnall, T.J.; Cirillo, J.; Nordstrom, M.A.; Semmler, J.G. Corticomotor plasticity and learning of a ballistic thumb training task are diminished in older adults. J. Appl. Physiol. (1985) 2009, 107, 1874–1883. [Google Scholar] [CrossRef]
- Bashir, S.; Perez, J.M.; Horvath, J.C.; Pena-Gomez, C.; Vernet, M.; Capia, A.; Alonso-Alonso, M.; Pascual-Leone, A. Differential effects of motor cortical excitability and plasticity in young and old individuals: A Transcranial Magnetic Stimulation (TMS) study. Front. Aging Neurosci. 2014, 6, 111. [Google Scholar] [CrossRef]
- Smith, A.E.; Ridding, M.C.; Higgins, R.D.; Wittert, G.A.; Pitcher, J.B. Age-related changes in short-latency motor cortex inhibition. Exp. Brain Res. 2009, 198, 489–500. [Google Scholar] [CrossRef]
- Nakamura, H.; Kitagawa, H.; Kawaguchi, Y.; Tsuji, H. Intracortical facilitation and inhibition after transcranial magnetic stimulation in conscious humans. J. Physiol. 1997, 498 Pt 3, 817–823. [Google Scholar] [CrossRef]
- Erro, R.; Rocchi, L.; Antelmi, E.; Liguori, R.; Tinazzi, M.; Berardelli, A.; Rothwell, J.; Bhatia, K.P. High frequency somatosensory stimulation in dystonia: Evidence fordefective inhibitory plasticity. Mov. Disord. 2018, 33, 1902–1909. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, U.; Tergau, F.; Wassermann, E.M.; Wischer, S.; Hildebrandt, J.; Paulus, W. Demonstration of facilitatory I wave interaction in the human motor cortex by paired transcranial magnetic stimulation. J. Physiol. 1998, 511 Pt 1, 181–190. [Google Scholar] [CrossRef]
- Hannah, R.; Rocchi, L.; Tremblay, S.; Wilson, E.; Rothwell, J.C. Pulse width biases the balance of excitation and inhibition recruited by transcranial magnetic stimulation. Brain Stimul. 2020, 13, 536–538. [Google Scholar] [CrossRef] [PubMed]
- Ortu, E.; Deriu, F.; Suppa, A.; Tolu, E.; Rothwell, J.C. Effects of volitional contraction on intracortical inhibition and facilitation in the human motor cortex. J. Physiol. 2008, 586, 5147–5159. [Google Scholar] [CrossRef] [PubMed]
- Shirota, Y.; Hamada, M.; Terao, Y.; Matsumoto, H.; Ohminami, S.; Furubayashi, T.; Nakatani-Enomoto, S.; Ugawa, Y.; Hanajima, R. Influence of short-interval intracortical inhibition on short-interval intracortical facilitation in human primary motor cortex. J. Neurophysiol. 2010, 104, 1382–1391. [Google Scholar] [CrossRef]
- Clark, J.; Loftus, A.; Hammond, G. Age-related changes in short-interval intracortical facilitation and dexterity. Neuroreport 2011, 22, 499–503. [Google Scholar] [CrossRef]
- Opie, G.M.; Cirillo, J.; Semmler, J.G. Age-related changes in late I-waves influence motor cortex plasticity induction in older adults. J. Physiol. 2018, 596, 2597–2609. [Google Scholar] [CrossRef]
- Opie, G.M.; Hand, B.J.; Semmler, J.G. Age-related changes in late synaptic inputs to corticospinal neurons and their functional significance: A paired-pulse TMS study. Brain Stimul. 2020, 13, 239–246. [Google Scholar] [CrossRef]
- Peurala, S.H.; Müller-Dahlhaus, J.F.M.; Arai, N.; Ziemann, U. Interference of short-interval intracortical inhibition (SICI) and short-interval intracortical facilitation (SICF). Clin. Neurophysiol. 2008, 119, 2291–2297. [Google Scholar] [CrossRef]
- Erro, R.; Antelmi, E.; Bhatia, K.P.; Latorre, A.; Tinazzi, M.; Berardelli, A.; Rothwell, J.C.; Rocchi, L. Reversal of Temporal Discrimination in Cervical Dystonia after Low-Frequency Sensory Stimulation. Mov. Disord. 2020. [Google Scholar] [CrossRef] [PubMed]
- Gövert, F.; Becktepe, J.; Balint, B.; Rocchi, L.; Brugger, F.; Garrido, A.; Walter, T.; Hannah, R.; Rothwell, J.; Elble, R.; et al. Temporal discrimination is altered in patients with isolated asymmetric and jerky upper limb tremor. Mov. Disord. 2020, 35, 306–315. [Google Scholar] [CrossRef]
- Latorre, A.; Cocco, A.; Bhatia, K.P.; Erro, R.; Antelmi, E.; Conte, A.; Rothwell, J.C.; Rocchi, L. Defective Somatosensory Inhibition and Plasticity Are Not Required to Develop Dystonia. Mov. Disord. 2020. [Google Scholar] [CrossRef]
- Hanajima, R.; Okabe, S.; Terao, Y.; Furubayashi, T.; Arai, N.; Inomata-Terada, S.; Hamada, M.; Yugeta, A.; Ugawa, Y. Difference in intracortical inhibition of the motor cortex between cortical myoclonus and focal hand dystonia. Clin. Neurophysiol. 2008, 119, 1400–1407. [Google Scholar] [CrossRef] [PubMed]
- Sale, M.V.; Lavender, A.P.; Opie, G.M.; Nordstrom, M.A.; Semmler, J.G. Increased intracortical inhibition in elderly adults with anterior-posterior current flow: A TMS study. Clin. Neurophysiol. 2016, 127, 635–640. [Google Scholar] [CrossRef]
- Ibáñez, J.; Spampinato, D.A.; Paraneetharan, V.; Rothwell, J.C. SICI during changing brain states: Differences in methodology can lead to different conclusions. Brain Stimul. 2020, 13, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Neubert, F.-X.; Mars, R.B.; Olivier, E.; Rushworth, M.F.S. Modulation of short intra-cortical inhibition during action reprogramming. Exp. Brain Res. 2011, 211, 265–276. [Google Scholar] [CrossRef]
- Hermans, L.; Maes, C.; Pauwels, L.; Cuypers, K.; Heise, K.-F.; Swinnen, S.P.; Leunissen, I. Age-related alterations in the modulation of intracortical inhibition during stopping of actions. Aging 2019, 11, 371–385. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Pilato, F.; Dileone, M.; Tonali, P.A.; Ziemann, U. Dissociated effects of diazepam and lorazepam on short-latency afferent inhibition. J. Physiol. 2005, 569, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Ponzo, D.; Hukkanen, T.; Mervaala, E.; Könönen, M.; Pasqualetti, P.; Vecchio, F.; Rossini, P.M.; Määttä, S. Human brain cortical correlates of short-latency afferent inhibition: A combined EEG-TMS study. J. Neurophysiol. 2012, 108, 314–323. [Google Scholar] [CrossRef]
- Hwang, Y.T.; Rocchi, L.; Hammond, P.; Hardy, C.J.; Warren, J.D.; Ridha, B.H.; Rothwell, J.; Rossor, M.N. Effect of Donepezil on Transcranial Magnetic Stimulation Parameters in Alzheimer’s Disease. Available online: https://pubmed.ncbi.nlm.nih.gov/29560413/ (accessed on 26 January 2021).
- Young-Bernier, M.; Kamil, Y.; Tremblay, F.; Davidson, P.S.R. Associations between a neurophysiological marker of central cholinergic activity and cognitive functions in young and older adults. Behav. Brain Funct. 2012, 8, 17. [Google Scholar] [CrossRef]
- Young-Bernier, M.; Tanguay, A.N.; Davidson, P.S.R.; Tremblay, F. Short-latency afferent inhibition is a poor predictor of individual susceptibility to rTMS-induced plasticity in the motor cortex of young and older adults. Front. Aging Neurosci. 2014, 6, 182. [Google Scholar] [CrossRef]
- Brown, K.E.; Neva, J.L.; Feldman, S.J.; Staines, W.R.; Boyd, L.A. Sensorimotor integration in healthy aging: Baseline differences and response to sensory training. Exp. Gerontol. 2018, 112, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, F.; Ponzo, V.; Bonnì, S.; Motta, C.; Negrão Serra, P.C.; Bozzali, M.; Caltagirone, C.; Martorana, A.; Koch, G. Long-term potentiation-like cortical plasticity is disrupted in Alzheimer’s disease patients independently from age of onset: Cortical Plasticity in Alzheimer’s Disease. Ann. Neurol. 2016, 80, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, U. Transcranial magnetic stimulation at the interface with other techniques: A powerful tool for studying the human cortex. Neuroscientist 2011, 17, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Rossini, P.M. TMS and TMS-EEG techniques in the study of the excitability, connectivity, and plasticity of the human motor cortex. Rev. Neurosci. 2013, 24, 431–442. [Google Scholar] [CrossRef]
- Ferreri, F.; Ponzo, D.; Vollero, L.; Guerra, A.; Di Pino, G.; Petrichella, S.; Benvenuto, A.; Tombini, M.; Rossini, L.; Denaro, L.; et al. Does an intraneural interface short-term implant for robotic hand control modulate sensorimotor cortical integration? An EEG-TMS co-registration study on a human amputee. Restor. Neurol. Neurosci. 2014, 32, 281–292. [Google Scholar] [CrossRef]
- Hannah, R.; Rocchi, L.; Tremblay, S.; Rothwell, J. Controllable Pulse Parameter TMS and TMS-EEG As Novel Approaches to Improve Neural Targeting with rTMS in Human Cerebral Cortex. Available online: https://pubmed.ncbi.nlm.nih.gov/27965543/ (accessed on 26 January 2021).
- Tremblay, S.; Rogasch, N.C.; Premoli, I.; Blumberger, D.M.; Casarotto, S.; Chen, R.; Di Lazzaro, V.; Farzan, F.; Ferrarelli, F.; Fitzgerald, P.B.; et al. Clinical utility and prospective of TMS-EEG. Clin. Neurophysiol. 2019, 130, 802–844. [Google Scholar] [CrossRef]
- Bonato, C.; Miniussi, C.; Rossini, P.M. Transcranial magnetic stimulation and cortical evoked potentials: A TMS/EEG co-registration study. Clin. Neurophysiol. 2006, 117, 1699–1707. [Google Scholar] [CrossRef]
- Ilmoniemi, R.J.; Kicić, D. Methodology for combined TMS and EEG. Brain Topogr. 2010, 22, 233–248. [Google Scholar] [CrossRef]
- Ilmoniemi, R.J.; Virtanen, J.; Ruohonen, J.; Karhu, J.; Aronen, H.J.; Näätänen, R.; Katila, T. Neuronal responses to magnetic stimulation reveal cortical reactivity and connectivity. Neuroreport 1997, 8, 3537–3540. [Google Scholar] [CrossRef] [PubMed]
- Gosseries, O.; Sarasso, S.; Casarotto, S.; Boly, M.; Schnakers, C.; Napolitani, M.; Bruno, M.-A.; Ledoux, D.; Tshibanda, J.-F.; Massimini, M.; et al. On the cerebral origin of EEG responses to TMS: Insights from severe cortical lesions. Brain Stimul. 2015, 8, 142–149. [Google Scholar] [CrossRef]
- Mäki, H.; Ilmoniemi, R.J. Projecting out muscle artifacts from TMS-evoked EEG. Neuroimage 2011, 54, 2706–2710. [Google Scholar] [CrossRef]
- Ilmoniemi, R.J.; Hernandez-Pavon, J.C.; Makela, N.N.; Metsomaa, J.; Mutanen, T.P.; Stenroos, M.; Sarvas, J. Dealing with artifacts in TMS-evoked EEG. In Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25–29 August 2015; Volume 2015, pp. 230–233. [Google Scholar]
- Rocchi, L.; Di Santo, A.; Brown, K.; Ibáñez, J.; Casula, E.; Rawji, V.; Di Lazzaro, V.; Koch, G.; Rothwell, J. Disentangling EEG responses to TMS due to cortical and peripheral activations. Brain Stimul. 2020, 14, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Pasqualetti, P.; Määttä, S.; Ponzo, D.; Ferrarelli, F.; Tononi, G.; Mervaala, E.; Miniussi, C.; Rossini, P.M. Human brain connectivity during single and paired pulse transcranial magnetic stimulation. Neuroimage 2011, 54, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Rossini, P.M.; Desiato, M.T.; Caramia, M.D. Age-related changes of motor evoked potentials in healthy humans: Non-invasive evaluation of central and peripheral motor tracts excitability and conductivity. Brain Res. 1992, 593, 14–19. [Google Scholar] [CrossRef]
- Ferreri, F.; Guerra, A.; Vollero, L.; Ponzo, D.; Maatta, S.; Mervaala, E.; Iannello, G.; Di Lazzaro, V. Age-related changes of cortical excitability and connectivity in healthy humans: Non-invasive evaluation of sensorimotor network by means of TMS-EEG. Neuroscience 2017, 357, 255–263. [Google Scholar] [CrossRef][Green Version]
- Lehmann, D.; Skrandies, W. Reference-free identification of components of checkerboard-evoked multichannel potential fields. Electroencephalogr. Clin. Neurophysiol. 1980, 48, 609–621. [Google Scholar] [CrossRef]
- Casula, E.P.; Rocchi, L.; Hannah, R.; Rothwell, J.C. Effects of pulse width, waveform and current direction in the cortex: A combined cTMS-EEG study. Brain Stimul. 2018, 11, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Opie, G.M.; Sidhu, S.K.; Rogasch, N.C.; Ridding, M.C.; Semmler, J.G. Cortical inhibition assessed using paired-pulse TMS-EEG is increased in older adults. Brain Stimul. 2018, 11, 545–557. [Google Scholar] [CrossRef]
- Hashimoto, T.; Arion, D.; Unger, T.; Maldonado-Avilés, J.G.; Morris, H.M.; Volk, D.W.; Mirnics, K.; Lewis, D.A. Alterations in GABA-related transcriptome in the dorsolateral prefrontal cortex of subjects with schizophrenia. Mol. Psychiatry 2008, 13, 147–161. [Google Scholar] [CrossRef]
- Fillman, S.G.; Duncan, C.E.; Webster, M.J.; Elashoff, M.; Weickert, C.S. Developmental co-regulation of the beta and gamma GABAA receptor subunits with distinct alpha subunits in the human dorsolateral prefrontal cortex. Int. J. Dev. Neurosci. 2010, 28, 513–519. [Google Scholar] [CrossRef]
- Duncan, C.E.; Webster, M.J.; Rothmond, D.A.; Bahn, S.; Elashoff, M.; Shannon Weickert, C. Prefrontal GABA(A) receptor alpha-subunit expression in normal postnatal human development and schizophrenia. J. Psychiatr. Res. 2010, 44, 673–681. [Google Scholar] [CrossRef]
- Matsumura, M.; Sawaguchi, T.; Oishi, T.; Ueki, K.; Kubota, K. Behavioral deficits induced by local injection of bicuculline and muscimol into the primate motor and premotor cortex. J. Neurophysiol. 1991, 65, 1542–1553. [Google Scholar] [CrossRef]
- Matsumura, M.; Sawaguchi, T.; Kubota, K. GABAergic inhibition of neuronal activity in the primate motor and premotor cortex during voluntary movement. J. Neurophysiol. 1992, 68, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Edden, R.A.E.; Li, M.; Puts, N.A.J.; Wang, G.; Liu, C.; Zhao, B.; Wang, H.; Bai, X.; Zhao, C.; et al. Edited magnetic resonance spectroscopy detects an age-related decline in brain GABA levels. Neuroimage 2013, 78, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Premoli, I.; Castellanos, N.; Rivolta, D.; Belardinelli, P.; Bajo, R.; Zipser, C.; Espenhahn, S.; Heidegger, T.; Müller-Dahlhaus, F.; Ziemann, U. TMS-EEG signatures of GABAergic neurotransmission in the human cortex. J. Neurosci. 2014, 34, 5603–5612. [Google Scholar] [CrossRef] [PubMed]
- Premoli, I.; Rivolta, D.; Espenhahn, S.; Castellanos, N.; Belardinelli, P.; Ziemann, U.; Müller-Dahlhaus, F. Characterization of GABAB-receptor mediated neurotransmission in the human cortex by paired-pulse TMS-EEG. Neuroimage 2014, 103, 152–162. [Google Scholar] [CrossRef]
- Noda, Y.; Zomorrodi, R.; Cash, R.F.H.; Barr, M.S.; Farzan, F.; Rajji, T.K.; Chen, R.; Daskalakis, Z.J.; Blumberger, D.M. Characterization of the influence of age on GABAA and glutamatergic mediated functions in the dorsolateral prefrontal cortex using paired-pulse TMS-EEG. Aging 2017, 9, 556–572. [Google Scholar] [CrossRef] [PubMed]
- Irlbacher, K.; Brocke, J.; Mechow, J.V.; Brandt, S.A. Effects of GABA(A) and GABA(B) agonists on interhemispheric inhibition in man. Clin. Neurophysiol. 2007, 118, 308–316. [Google Scholar] [CrossRef]
- Ferbert, A.; Priori, A.; Rothwell, J.C.; Day, B.L.; Colebatch, J.G.; Marsden, C.D. Interhemispheric inhibition of the human motor cortex. J. Physiol. 1992, 453, 525–546. [Google Scholar] [CrossRef]
- Wahl, M.; Lauterbach-Soon, B.; Hattingen, E.; Jung, P.; Singer, O.; Volz, S.; Klein, J.C.; Steinmetz, H.; Ziemann, U. Human motor corpus callosum: Topography, somatotopy, and link between microstructure and function. J. Neurosci. 2007, 27, 12132–12138. [Google Scholar] [CrossRef]
- Ni, Z.; Gunraj, C.; Nelson, A.J.; Yeh, I.-J.; Castillo, G.; Hoque, T.; Chen, R. Two phases of interhemispheric inhibition between motor related cortical areas and the primary motor cortex in human. Cereb. Cortex 2009, 19, 1654–1665. [Google Scholar] [CrossRef] [PubMed]
- Plow, E.B.; Cunningham, D.A.; Bonnett, C.; Gohar, D.; Bayram, M.; Wyant, A.; Varnerin, N.; Mamone, B.; Siemionow, V.; Hou, J.; et al. Neurophysiological correlates of aging-related muscle weakness. J. Neurophysiol. 2013, 110, 2563–2573. [Google Scholar] [CrossRef]
- Mooney, R.A.; Cirillo, J.; Byblow, W.D. Adaptive threshold hunting reveals differences in interhemispheric inhibition between young and older adults. Eur. J. Neurosci. 2018, 48, 2247–2258. [Google Scholar] [CrossRef]
- Talelli, P.; Waddingham, W.; Ewas, A.; Rothwell, J.C.; Ward, N.S. The effect of age on task-related modulation of interhemispheric balance. Exp. Brain Res. 2008, 186, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Hinder, M.R.; Schmidt, M.W.; Garry, M.I.; Summers, J.J. Unilateral contractions modulate interhemispheric inhibition most strongly and most adaptively in the homologous muscle of the contralateral limb. Exp. Brain Res. 2010, 205, 423–433. [Google Scholar] [CrossRef]
- Talelli, P.; Ewas, A.; Waddingham, W.; Rothwell, J.C.; Ward, N.S. Neural correlates of age-related changes in cortical neurophysiology. Neuroimage 2008, 40, 1772–1781. [Google Scholar] [CrossRef]
- Hinder, M.R.; Fujiyama, H.; Summers, J.J. Premotor-motor interhemispheric inhibition is released during movement initiation in older but not young adults. PLoS ONE 2012, 7, e52573. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fujiyama, H.; Van Soom, J.; Rens, G.; Gooijers, J.; Leunissen, I.; Levin, O.; Swinnen, S.P. Age-Related Changes in Frontal Network Structural and Functional Connectivity in Relation to Bimanual Movement Control. J. Neurosci. 2016, 36, 1808–1822. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R. Cognitive neuroscience of aging: Contributions of functional neuroimaging. Scand. J. Psychol. 2001, 42, 277–286. [Google Scholar] [CrossRef]
- Ward, N.S.; Swayne, O.B.C.; Newton, J.M. Age-dependent changes in the neural correlates of force modulation: An fMRI study. Neurobiol. Aging 2008, 29, 1434–1446. [Google Scholar] [CrossRef]
- Boudrias, M.-H.; Gonçalves, C.S.; Penny, W.D.; Park, C.; Rossiter, H.E.; Talelli, P.; Ward, N.S. Age-related changes in causal interactions between cortical motor regions during hand grip. Neuroimage 2012, 59, 3398–3405. [Google Scholar] [CrossRef]
- Fujiyama, H.; Hinder, M.R.; Summers, J.J. Functional role of left PMd and left M1 during preparation and execution of left hand movements in older adults. J. Neurophysiol. 2013, 110, 1062–1069. [Google Scholar] [CrossRef]
- Chen, R.; Yung, D.; Li, J.-Y. Organization of ipsilateral excitatory and inhibitory pathways in the human motor cortex. J. Neurophysiol. 2003, 89, 1256–1264. [Google Scholar] [CrossRef]
- Davidson, T.; Tremblay, F. Age and hemispheric differences in transcallosal inhibition between motor cortices: An ispsilateral silent period study. BMC Neurosci. 2013, 14, 62. [Google Scholar] [CrossRef]
- Petitjean, M.; Ko, J.Y.L. An age-related change in the ipsilateral silent period of a small hand muscle. Clin. Neurophysiol. 2013, 124, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Coppi, E.; Houdayer, E.; Chieffo, R.; Spagnolo, F.; Inuggi, A.; Straffi, L.; Comi, G.; Leocani, L. Age-related changes in motor cortical representation and interhemispheric interactions: A transcranial magnetic stimulation study. Front. Aging Neurosci. 2014, 6, 209. [Google Scholar] [CrossRef]
- Strauss, S.; Lotze, M.; Flöel, A.; Domin, M.; Grothe, M. Changes in Interhemispheric Motor Connectivity Across the Lifespan: A Combined TMS and DTI Study. Front. Aging Neurosci. 2019, 11, 12. [Google Scholar] [CrossRef]
- Ward, N.S. Compensatory mechanisms in the aging motor system. Ageing Res. Rev. 2006, 5, 239–254. [Google Scholar] [CrossRef]
- Seidler, R.D.; Bernard, J.A.; Burutolu, T.B.; Fling, B.W.; Gordon, M.T.; Gwin, J.T.; Kwak, Y.; Lipps, D.B. Motor control and aging: Links to age-related brain structural, functional, and biochemical effects. Neurosci. Biobehav. Rev. 2010, 34, 721–733. [Google Scholar] [CrossRef]
- Lee, L.; Harrison, L.M.; Mechelli, A. A report of the functional connectivity workshop, Dusseldorf 2002. Neuroimage 2003, 19, 457–465. [Google Scholar] [CrossRef]
- Komssi, S.; Kähkönen, S. The novelty value of the combined use of electroencephalography and transcranial magnetic stimulation for neuroscience research. Brain Res. Rev. 2006, 52, 183–192. [Google Scholar] [CrossRef]
- Casula, E.P.; Maiella, M.; Pellicciari, M.C.; Porrazzini, F.; D’Acunto, A.; Rocchi, L.; Koch, G. Novel TMS-EEG indexes to investigate interhemispheric dynamics in humans. Clin. Neurophysiol. 2020, 131, 70–77. [Google Scholar] [CrossRef]
- Casula, E.P.; Pellicciari, M.C.; Bonnì, S.; Spanò, B.; Ponzo, V.; Salsano, I.; Giulietti, G.; Martino Cinnera, A.; Maiella, M.; Borghi, I.; et al. Evidence for interhemispheric imbalance in stroke patients as revealed by combining transcranial magnetic stimulation and electroencephalography. Hum. Brain Mapp. 2021, 42, 1343–1358. [Google Scholar] [CrossRef]
- Massimini, M.; Ferrarelli, F.; Huber, R.; Esser, S.K.; Singh, H.; Tononi, G. Breakdown of cortical effective connectivity during sleep. Science 2005, 309, 2228–2232. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Vecchio, F.; Ponzo, D.; Pasqualetti, P.; Rossini, P.M. Time-varying coupling of EEG oscillations predicts excitability fluctuations in the primary motor cortex as reflected by motor evoked potentials amplitude: An EEG-TMS study. Hum. Brain Mapp. 2014, 35, 1969–1980. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Rocchi, L.; Hannah, R.; Xu, G.; Rothwell, J.C.; Ibáñez, J. Corticospinal excitability modulation by pairing peripheral nerve stimulation with cortical states of movement initiation. J. Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Desideri, D.; Zrenner, C.; Ziemann, U.; Belardinelli, P. Phase of sensorimotor μ-oscillation modulates cortical responses to transcranial magnetic stimulation of the human motor cortex. J. Physiol. 2019, 597, 5671–5686. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Vecchio, F.; Guerra, A.; Miraglia, F.; Ponzo, D.; Vollero, L.; Iannello, G.; Maatta, S.; Mervaala, E.; Rossini, P.M.; et al. Age related differences in functional synchronization of EEG activity as evaluated by means of TMS-EEG coregistrations. Neurosci. Lett. 2017, 647, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Kiers, L.; Cros, D.; Chiappa, K.H.; Fang, J. Variability of motor potentials evoked by transcranial magnetic stimulation. Electroencephalogr. Clin. Neurophysiol. 1993, 89, 415–423. [Google Scholar] [CrossRef]
- Thickbroom, G.W.; Byrnes, M.L.; Mastaglia, F.L. A model of the effect of MEP amplitude variation on the accuracy of TMS mapping. Clin. Neurophysiol. 1999, 110, 941–943. [Google Scholar] [CrossRef]
- Schulz, H.; Ubelacker, T.; Keil, J.; Müller, N.; Weisz, N. Now I am ready-now i am not: The influence of pre-TMS oscillations and corticomuscular coherence on motor-evoked potentials. Cereb. Cortex 2014, 24, 1708–1719. [Google Scholar] [CrossRef]
- Gloor, P.; Ball, G.; Schaul, N. Brain lesions that produce delta waves in the EEG. Neurology 1977, 27, 326–333. [Google Scholar] [CrossRef]
- Giovanni, A.; Capone, F.; di Biase, L.; Ferreri, F.; Florio, L.; Guerra, A.; Marano, M.; Paolucci, M.; Ranieri, F.; Salomone, G.; et al. Oscillatory Activities in Neurological Disorders of Elderly: Biomarkers to Target for Neuromodulation. Front. Aging Neurosci. 2017, 9, 189. [Google Scholar]
- Hebb, D. The Organization of Behavior; Wiley: New York, NY, USA, 1949. [Google Scholar]
- Dan, Y.; Poo, M.-M. Spike timing-dependent plasticity: From synapse to perception. Physiol. Rev. 2006, 86, 1033–1048. [Google Scholar] [CrossRef]
- Caporale, N.; Dan, Y. Spike timing-dependent plasticity: A Hebbian learning rule. Annu. Rev. Neurosci. 2008, 31, 25–46. [Google Scholar] [CrossRef]
- Feldman, D.E. The spike-timing dependence of plasticity. Neuron 2012, 75, 556–571. [Google Scholar] [CrossRef]
- Suppa, A.; Asci, F.; Guerra, A. TMS as a Tool to Induce and Explore Plasticity in Humans. Handb. Clin. Neurol. 2021. [Google Scholar]
- Stefan, K.; Kunesch, E.; Cohen, L.G.; Benecke, R.; Classen, J. Induction of plasticity in the human motor cortex by paired associative stimulation. Brain 2000, 123 Pt 3, 572–584. [Google Scholar] [CrossRef]
- Suppa, A.; Quartarone, A.; Siebner, H.; Chen, R.; Di Lazzaro, V.; Del Giudice, P.; Paulus, W.; Rothwell, J.C.; Ziemann, U.; Classen, J. The associative brain at work: Evidence from paired associative stimulation studies in humans. Clin. Neurophysiol. 2017, 128, 2140–2164. [Google Scholar] [CrossRef]
- Müller-Dahlhaus, F.; Ziemann, U.; Classen, J. Plasticity resembling spike-timing dependent synaptic plasticity: The evidence in human cortex. Front. Synaptic. Neurosci. 2010, 2, 34. [Google Scholar] [CrossRef]
- Larson, J.; Lynch, G. Theta pattern stimulation and the induction of LTP: The sequence in which synapses are stimulated determines the degree to which they potentiate. Brain Res. 1989, 489, 49–58. [Google Scholar] [CrossRef]
- Bliss, T.V.; Lomo, T. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol. 1973, 232, 331–356. [Google Scholar] [CrossRef] [PubMed]
- Cooke, S.F.; Bliss, T.V.P. Plasticity in the human central nervous system. Brain 2006, 129, 1659–1673. [Google Scholar] [CrossRef]
- Huang, Y.-Z.; Edwards, M.J.; Rounis, E.; Bhatia, K.P.; Rothwell, J.C. Theta burst stimulation of the human motor cortex. Neuron 2005, 45, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Suppa, A.; Huang, Y.-Z.; Funke, K.; Ridding, M.C.; Cheeran, B.; Di Lazzaro, V.; Ziemann, U.; Rothwell, J.C. Ten Years of Theta Burst Stimulation in Humans: Established Knowledge, Unknowns and Prospects. Brain Stimul. 2016, 9, 323–335. [Google Scholar] [CrossRef]
- Georgiev, D.; Rocchi, L.; Tocco, P.; Speekenbrink, M.; Rothwell, J.C.; Jahanshahi, M. Continuous Theta Burst Stimulation Over the Dorsolateral Prefrontal Cortex and the Pre-SMA Alter Drift Rate and Response Thresholds Respectively During Perceptual Decision-Making. Brain Stimul. 2016, 9, 601–608. [Google Scholar] [CrossRef]
- Méndez, J.C.; Rocchi, L.; Jahanshahi, M.; Rothwell, J.; Merchant, H. Probing the timing network: A continuous theta burst stimulation study of temporal categorization. Neuroscience 2017, 356, 167–175. [Google Scholar] [CrossRef]
- Dumitru, A.; Rocchi, L.; Saini, F.; Rothwell, J.C.; Roiser, J.P.; David, A.S.; Richieri, R.M.; Lewis, G.; Lewis, G. Influence of theta-burst transcranial magnetic stimulation over the dorsolateral prefrontal cortex on emotion processing in healthy volunteers. Cogn. Affect. Behav. Neurosci. 2020, 20, 1278–1293. [Google Scholar] [CrossRef]
- Müller-Dahlhaus, J.F.M.; Orekhov, Y.; Liu, Y.; Ziemann, U. Interindividual variability and age-dependency of motor cortical plasticity induced by paired associative stimulation. Exp. Brain Res. 2008, 187, 467–475. [Google Scholar] [CrossRef]
- Tecchio, F.; Zappasodi, F.; Pasqualetti, P.; Gennaro, L.D.; Pellicciari, M.C.; Ercolani, M.; Squitti, R.; Rossini, P.M. Age dependence of primary motor cortex plasticity induced by paired associative stimulation. Clin. Neurophysiol. 2008, 119, 675–682. [Google Scholar] [CrossRef]
- Polimanti, R.; Simonelli, I.; Zappasodi, F.; Ventriglia, M.; Pellicciari, M.C.; Benussi, L.; Squitti, R.; Rossini, P.M.; Tecchio, F. Biological factors and age-dependence of primary motor cortex experimental plasticity. Neurol. Sci. 2016, 37, 211–218. [Google Scholar] [CrossRef]
- Fathi, D.; Ueki, Y.; Mima, T.; Koganemaru, S.; Nagamine, T.; Tawfik, A.; Fukuyama, H. Effects of aging on the human motor cortical plasticity studied by paired associative stimulation. Clin. Neurophysiol. 2010, 121, 90–93. [Google Scholar] [CrossRef]
- Kishore, A.; Popa, T.; James, P.; Yahia-Cherif, L.; Backer, F.; Varughese Chacko, L.; Govind, P.; Pradeep, S.; Meunier, S. Age-related decline in the responsiveness of motor cortex to plastic forces reverses with levodopa or cerebellar stimulation. Neurobiol. Aging 2014, 35, 2541–2551. [Google Scholar] [CrossRef]
- Dickins, D.S.E.; Sale, M.V.; Kamke, M.R. Plasticity Induced by Intermittent Theta Burst Stimulation in Bilateral Motor Cortices Is Not Altered in Older Adults. Neural Plast. 2015, 2015, 323409. [Google Scholar] [CrossRef] [PubMed]
- Opie, G.M.; Vosnakis, E.; Ridding, M.C.; Ziemann, U.; Semmler, J.G. Priming theta burst stimulation enhances motor cortex plasticity in young but not old adults. Brain Stimul. 2017, 10, 298–304. [Google Scholar] [CrossRef]
- Guerra, A.; Asci, F.; Zampogna, A.; D’Onofrio, V.; Berardelli, A.; Suppa, A. The effect of gamma oscillations in boosting primary motor cortex plasticity is greater in young than older adults. Clin. Neurophysiol. 2021. [Google Scholar] [CrossRef]
- Guerra, A.; López-Alonso, V.; Cheeran, B.; Suppa, A. Variability in non-invasive brain stimulation studies: Reasons and results. Neurosci. Lett. 2020, 719, 133330. [Google Scholar] [CrossRef]
- Huang, Y.-Z.; Lu, M.-K.; Antal, A.; Classen, J.; Nitsche, M.; Ziemann, U.; Ridding, M.; Hamada, M.; Ugawa, Y.; Jaberzadeh, S.; et al. Plasticity induced by non-invasive transcranial brain stimulation: A position paper. Clin. Neurophysiol. 2017, 128, 2318–2329. [Google Scholar] [CrossRef]
- Suppa, A.; Li Voti, P.; Rocchi, L.; Papazachariadis, O.; Berardelli, A. Early visuomotor integration processes induce LTP/LTD-like plasticity in the human motor cortex. Cereb. Cortex 2015, 25, 703–712. [Google Scholar] [CrossRef]
- Suppa, A.; Rocchi, L.; Li Voti, P.; Papazachariadis, O.; Casciato, S.; Di Bonaventura, C.; Giallonardo, A.T.; Berardelli, A. The Photoparoxysmal Response Reflects Abnormal Early Visuomotor Integration in the Human Motor Cortex. Brain Stimul. 2015, 8, 1151–1161. [Google Scholar] [CrossRef]
- Abraham, W.C. Metaplasticity: Tuning synapses and networks for plasticity. Nat. Rev. Neurosci. 2008, 9, 387. [Google Scholar] [CrossRef]
- Müller-Dahlhaus, F.; Ziemann, U. Metaplasticity in human cortex. Neuroscientist 2015, 21, 185–202. [Google Scholar] [CrossRef]
- Hulme, S.R.; Jones, O.D.; Abraham, W.C. Emerging roles of metaplasticity in behaviour and disease. Trends Neurosci. 2013, 36, 353–362. [Google Scholar] [CrossRef]
- Opie, G.M.; Post, A.K.; Ridding, M.C.; Ziemann, U.; Semmler, J.G. Modulating motor cortical neuroplasticity with priming paired associative stimulation in young and old adults. Clin. Neurophysiol. 2017, 128, 763–769. [Google Scholar] [CrossRef]
- Opie, G.M.; Hand, B.J.; Coxon, J.P.; Ridding, M.C.; Ziemann, U.; Semmler, J.G. Visuomotor task acquisition is reduced by priming paired associative stimulation in older adults. Neurobiol. Aging 2019, 81, 67–76. [Google Scholar] [CrossRef]
- Guerra, A.; Suppa, A.; Bologna, M.; D’Onofrio, V.; Bianchini, E.; Brown, P.; Di Lazzaro, V.; Berardelli, A. Boosting the LTP-like plasticity effect of intermittent theta-burst stimulation using gamma transcranial alternating current stimulation. Brain Stimul. 2018, 11, 734–742. [Google Scholar] [CrossRef]
- Guerra, A.; Suppa, A.; Asci, F.; De Marco, G.; D’Onofrio, V.; Bologna, M.; Di Lazzaro, V.; Berardelli, A. LTD-like plasticity of the human primary motor cortex can be reversed by γ-tACS. Brain Stimul. 2019, 12, 1490–1499. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.; Asci, F.; Zampogna, A.; D’Onofrio, V.; Petrucci, S.; Ginevrino, M.; Berardelli, A.; Suppa, A. Gamma-transcranial alternating current stimulation and theta-burst stimulation: Inter-subject variability and the role of BDNF. Clin. Neurophysiol. 2020, 131, 2691–2699. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.; López-Alonso, V.; Cheeran, B.; Suppa, A. Solutions for managing variability in non-invasive brain stimulation studies. Neurosci. Lett. 2020, 719, 133332. [Google Scholar] [CrossRef]
- Stinear, C.M.; Byblow, W.D. Elevated threshold for intracortical inhibition in focal hand dystonia. Mov. Disord. 2004, 19, 1312–1317. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Profice, P.; Ranieri, F.; Capone, F.; Dileone, M.; Oliviero, A.; Pilato, F. I-wave origin and modulation. Brain Stimul. 2012, 5, 512–525. [Google Scholar] [CrossRef]
- Opie, G.M.; Semmler, J.G. Preferential Activation of Unique Motor Cortical Networks With Transcranial Magnetic Stimulation: A Review of the Physiological, Functional, and Clinical Evidence. Neuromodulation 2020. [Google Scholar] [CrossRef]
- Van den Bos, M.A.J.; Higashihara, M.; Geevasinga, N.; Menon, P.; Kiernan, M.C.; Vucic, S. Imbalance of cortical facilitatory and inhibitory circuits underlies hyperexcitability in ALS. Neurology 2018, 91, e1669–e1676. [Google Scholar] [CrossRef]
- Ni, Z.; Bahl, N.; Gunraj, C.A.; Mazzella, F.; Chen, R. Increased motor cortical facilitation and decreased inhibition in Parkinson disease. Neurology 2013, 80, 1746–1753. [Google Scholar] [CrossRef]
- Shirota, Y.; Ohminami, S.; Tsutsumi, R.; Terao, Y.; Ugawa, Y.; Tsuji, S.; Hanajima, R. Increased facilitation of the primary motor cortex in de novo Parkinson’s disease. Parkinsonism. Relat. Disord. 2019, 66, 125–129. [Google Scholar] [CrossRef]
- Ammann, C.; Dileone, M.; Pagge, C.; Catanzaro, V.; Mata-Marín, D.; Hernández-Fernández, F.; Monje, M.H.G.; Sánchez-Ferro, Á.; Fernández-Rodríguez, B.; Gasca-Salas, C.; et al. Cortical disinhibition in Parkinson’s disease. Brain 2020, 143, 3408–3421. [Google Scholar] [CrossRef]
- Rocchi, L.; Ibáñez, J.; Benussi, A.; Hannah, R.; Rawji, V.; Casula, E.; Rothwell, J. Variability and Predictors of Response to Continuous Theta Burst Stimulation: A TMS-EEG Study. Front. Neurosci. 2018, 12, 400. [Google Scholar] [CrossRef] [PubMed]
- Premoli, I.; Király, J.; Müller-Dahlhaus, F.; Zipser, C.M.; Rossini, P.; Zrenner, C.; Ziemann, U.; Belardinelli, P. Short-interval and long-interval intracortical inhibition of TMS-evoked EEG potentials. Brain Stimul. 2018, 11, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Rawji, V.; Kaczmarczyk, I.; Rocchi, L.; Rothwell, J.C.; Sharma, N. Short interval intracortical inhibition as measured by TMS-EEG. Sci. Rep. 2021. [Google Scholar] [CrossRef]
- Rossini, P.M.; Di Iorio, R.; Bentivoglio, M.; Bertini, G.; Ferreri, F.; Gerloff, C.; Ilmoniemi, R.J.; Miraglia, F.; Nitsche, M.A.; Pestilli, F.; et al. Methods for analysis of brain connectivity: An IFCN-sponsored review. Clin. Neurophysiol. 2019, 130, 1833–1858. [Google Scholar] [CrossRef]
- Belardinelli, P.; Biabani, M.; Blumberger, D.M.; Bortoletto, M.; Casarotto, S.; David, O.; Desideri, D.; Etkin, A.; Ferrarelli, F.; Fitzgerald, P.B.; et al. Reproducibility in TMS-EEG studies: A call for data sharing, standard procedures and effective experimental control. Brain Stimul. 2019, 12, 787–790. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerra, A.; Rocchi, L.; Grego, A.; Berardi, F.; Luisi, C.; Ferreri, F. Contribution of TMS and TMS-EEG to the Understanding of Mechanisms Underlying Physiological Brain Aging. Brain Sci. 2021, 11, 405. https://doi.org/10.3390/brainsci11030405
Guerra A, Rocchi L, Grego A, Berardi F, Luisi C, Ferreri F. Contribution of TMS and TMS-EEG to the Understanding of Mechanisms Underlying Physiological Brain Aging. Brain Sciences. 2021; 11(3):405. https://doi.org/10.3390/brainsci11030405
Chicago/Turabian StyleGuerra, Andrea, Lorenzo Rocchi, Alberto Grego, Francesca Berardi, Concetta Luisi, and Florinda Ferreri. 2021. "Contribution of TMS and TMS-EEG to the Understanding of Mechanisms Underlying Physiological Brain Aging" Brain Sciences 11, no. 3: 405. https://doi.org/10.3390/brainsci11030405
APA StyleGuerra, A., Rocchi, L., Grego, A., Berardi, F., Luisi, C., & Ferreri, F. (2021). Contribution of TMS and TMS-EEG to the Understanding of Mechanisms Underlying Physiological Brain Aging. Brain Sciences, 11(3), 405. https://doi.org/10.3390/brainsci11030405