
Transcranial Magnetic Stimulation of the Right Superior Parietal Lobule Modulates the Retro-Cue Benefit in Visual Short-Term Memory



{kind=link}
{kind=link}
Abstract
1. Introduction
Aims of the Present Study
2. Materials and Methods
2.1. Participants
2.2. Apparatus and Stimuli
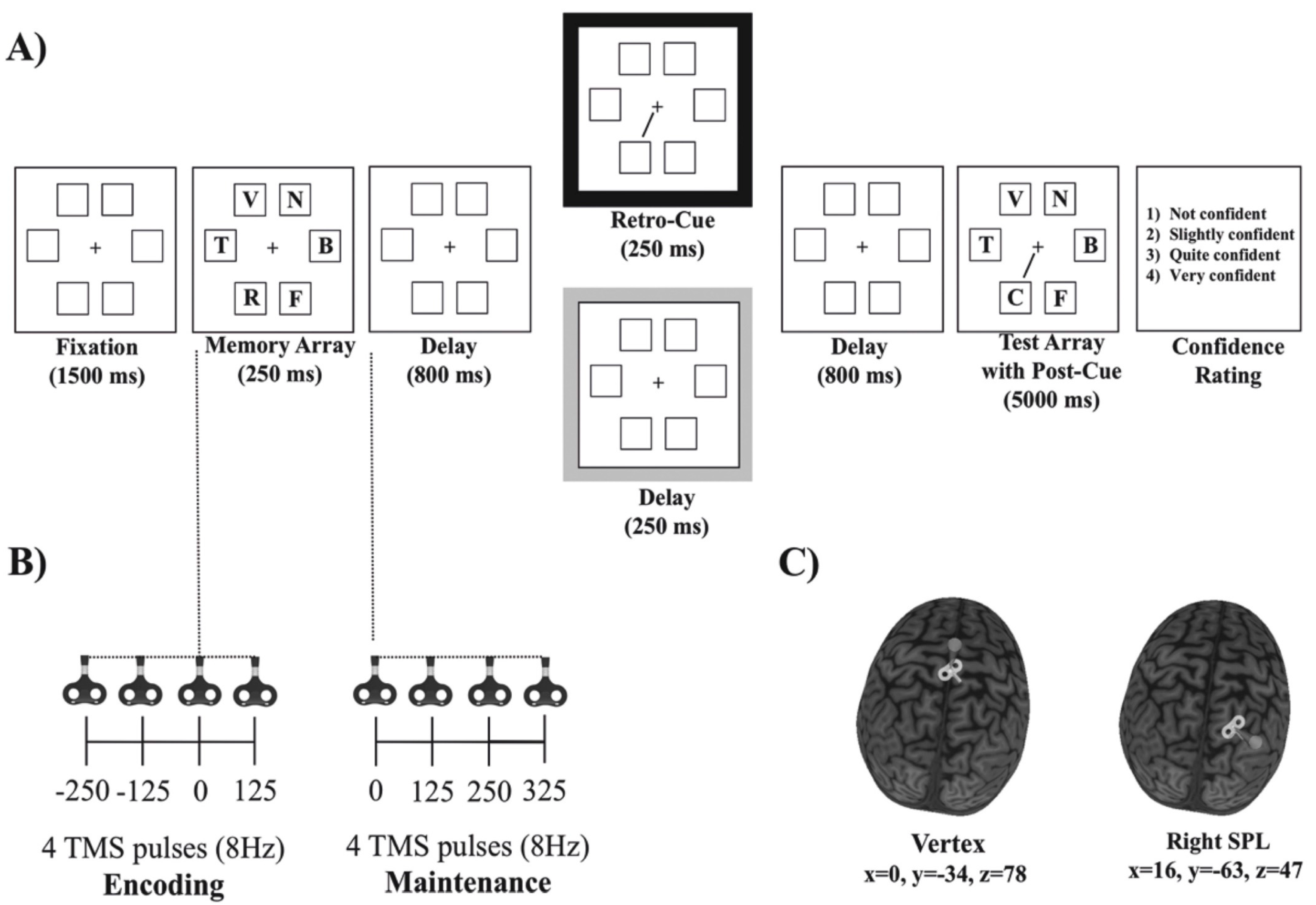
2.3. Behavioral Procedure
2.4. MRI and TMS Protocol
2.5. Diffusion Tensor Imaging (DTI) Analysis
2.6. Design and Data Analysis
3. Results
3.1. Retro-Cue vs. Post-Cue Analysis
3.2. Retro-Cue Benefit Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luck, S.J.; Vogel, E.K. The Capacity of Visual Working Memory for Features and Conjunctions. Nature 1997, 390, 279–281. [Google Scholar] [CrossRef]
- Averbach, E.; Coriell, A.S. Short-Term Memory in Vision. Bell Syst. Tech. J. 1961, 40, 309–328. [Google Scholar] [CrossRef]
- Sperling, G. The Information Available in Brief Visual Presentations. Psychol. Monogr. Gen. Appl. 1960, 74, 1–29. [Google Scholar] [CrossRef]
- Griffin, I.C.; Nobre, A.C. Orienting Attention to Locations in Internal Representations. J. Cogn. Neurosci. 2003, 15, 1176–1194. [Google Scholar] [CrossRef]
- Landman, R.; Spekreijse, H.; Lamme, V.A.F. Large Capacity Storage of Integrated Objects before Change Blindness. Vis. Res. 2003, 43, 149–164. [Google Scholar] [CrossRef]
- Sligte, I.G.; Scholte, H.S.; Lamme, V.A.F. Are There Multiple Visual Short-Term Memory Stores? PLoS ONE 2008, 3, e1699. [Google Scholar] [CrossRef] [PubMed]
- Vandenbroucke, A.R.E.; Sligte, I.G.; Lamme, V.A.F. Manipulations of Attention Dissociate Fragile Visual Short-Term Memory from Visual Working Memory. Neuropsychologia 2011, 49, 1559–1568. [Google Scholar] [CrossRef]
- Sligte, I.G.; Wokke, M.E.; Tesselaar, J.P.; Steven Scholte, H.; Lamme, V.A.F. Magnetic Stimulation of the Dorsolateral Prefrontal Cortex Dissociates Fragile Visual Short-Term Memory from Visual Working Memory. Neuropsychologia 2011, 49, 1578–1588. [Google Scholar] [CrossRef] [PubMed]
- Vandenbroucke, A.R.E.; Sligte, I.G.; Barrett, A.B.; Seth, A.K.; Fahrenfort, J.J.; Lamme, V.A.F. Accurate Metacognition for Visual Sensory Memory Representations. Psychol. Sci. 2014, 25, 861–873. [Google Scholar] [CrossRef]
- Lamme, V.A.F. Why Visual Attention and Awareness Are Different. Trends Cogn. Sci. 2003, 7, 12–18. [Google Scholar] [CrossRef]
- Lepsien, J.; Griffin, I.C.; Devlin, J.T.; Nobre, A.C. Directing Spatial Attention in Mental Representations: Interactions between Attentional Orienting and Working-Memory Load. Neuroimage 2005, 26, 733–743. [Google Scholar] [CrossRef]
- Pessoa, L.; Gutierrez, E.; Bandettini, P.A.; Ungerleider, L.G. Neural Correlates of Visual Working Memory: FMRI Amplitude Predicts Task Performance. Neuron 2002, 35, 975–987. [Google Scholar] [CrossRef]
- Sligte, I.G.; Scholte, H.S.; Lamme, V.A.F. V4 Activity Predicts the Strength of Visual Short-Term Memory Representations. J. Neurosci. 2009, 29, 7432–7438. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.S.; Oberauer, K. In Search of the Focus of Attention in Working Memory: 13 Years of the Retro-Cue Effect. Atten. Percept. Psychophys. 2016, 78, 1839–1860. [Google Scholar] [CrossRef] [PubMed]
- Makovski, T. Are Multiple Visual Short-Term Memory Storages Necessary to Explain the Retro-Cue Effect? Psychon. Bull. Rev. 2012, 19, 470–476. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Botta, F.; Martín-Arévalo, E.; Lupiáñez, J.; Bartolomeo, P. Does Spatial Attention Modulate Sensory Memory? PLoS ONE 2019, 14, e0219504. [Google Scholar] [CrossRef]
- Mack, A.; Erol, M.; Clarke, J. Iconic Memory Is Not a Case of Attention-Free Awareness. Conscious. Cogn. 2015, 33, 291–299. [Google Scholar] [CrossRef]
- Mack, A.; Erol, M.; Clarke, J.; Bert, J. No Iconic Memory without Attention. Conscious. Cogn. 2016, 40, 1–8. [Google Scholar] [CrossRef]
- Pinto, Y.; Vandenbroucke, A.R.; Otten, M.; Sligte, I.G.; Seth, A.K.; Lamme, V.A.F. Conscious Visual Memory with Minimal Attention. J. Exp. Psychol. Gen. 2017, 146, 214–226. [Google Scholar] [CrossRef]
- Awh, E.; Jonides, J. Overlapping Mechanisms of Attention and Spatial Working Memory. Trends Cogn. Sci. 2001, 5, 119–126. [Google Scholar] [CrossRef]
- Awh, E.; Vogel, E.K.; Oh, S.-H. Interactions between Attention and Working Memory. Neuroscience 2006, 139, 201–208. [Google Scholar] [CrossRef]
- Hopfinger, J.B.; Woldorff, M.G.; Fletcher, E.M.; Mangun, G.R. Dissociating Top-down Attentional Control from Selective Perception and Action. Neuropsychologia 2001, 39, 1277–1291. [Google Scholar] [CrossRef]
- LaBar, K.S.; Gitelman, D.R.; Parrish, T.B.; Mesulam, M. Neuroanatomic Overlap of Working Memory and Spatial Attention Networks: A Functional MRI Comparison within Subjects. Neuroimage 1999, 10, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Nee, D.E.; Jonides, J. Common and Distinct Neural Correlates of Perceptual and Memorial Selection. Neuroimage 2009, 45, 963–975. [Google Scholar] [CrossRef]
- Corbetta, M.; Shulman, G.L. Control of Goal-Directed and Stimulus-Driven Attention in the Brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Corbetta, M.; Patel, G.; Shulman, G.L. The Reorienting System of the Human Brain: From Environment to Theory of Mind. Neuron 2008, 58, 306–324. [Google Scholar] [CrossRef] [PubMed]
- Naghavi, H.R.; Nyberg, L. Common Fronto-Parietal Activity in Attention, Memory, and Consciousness: Shared Demands on Integration? Conscious. Cogn. 2005, 14, 390–425. [Google Scholar] [CrossRef]
- Santangelo, V.; Di Francesco, S.A.; Mastroberardino, S.; Macaluso, E. Parietal Cortex Integrates Contextual and Saliency Signals during the Encoding of Natural Scenes in Working Memory. Hum. Brain Mapp. 2015, 36, 5003–5017. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, V.; Macaluso, E. The Contribution of Working Memory to Divided Attention. Hum. Brain Mapp. 2013, 34, 158–175. [Google Scholar] [CrossRef]
- Vossel, S.; Geng, J.J.; Fink, G.R. Dorsal and Ventral Attention Systems: Distinct Neural Circuits but Collaborative Roles. Neuroscientist 2014, 20, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Thiebaut de Schotten, M.; Dell’Acqua, F.; Forkel, S.J.; Simmons, A.; Vergani, F.; Murphy, D.G.M.; Catani, M. A Lateralized Brain Network for Visuospatial Attention. Nat. Neurosci. 2011, 14, 1245–1246. [Google Scholar] [CrossRef]
- Chica, A.B.; Thiebaut de Schotten, M.; Bartolomeo, P.; Paz-Alonso, P.M. White Matter Microstructure of Attentional Networks Predicts Attention and Consciousness Functional Interactions. Brain Struct. Funct. 2018, 223, 653–668. [Google Scholar] [CrossRef]
- Umarova, R.M.; Saur, D.; Schnell, S.; Kaller, C.P.; Vry, M.-S.; Glauche, V.; Rijntjes, M.; Hennig, J.; Kiselev, V.; Weiller, C. Structural Connectivity for Visuospatial Attention: Significance of Ventral Pathways. Cereb. Cortex 2010, 20, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, A.; Chica, A.B.; Migliaccio, R.; de Schotten, M.T.; Bartolomeo, P. Cortical Control of Inhibition of Return: Evidence from Patients with Inferior Parietal Damage and Visual Neglect. Neuropsychologia 2012, 50, 800–809. [Google Scholar] [CrossRef]
- Doricchi, F.; Thiebaut de Schotten, M.; Tomaiuolo, F.; Bartolomeo, P. White Matter (Dis) Connections and Gray Matter (Dys) Functions in Visual Neglect: Gaining Insights into the Brain Networks of Spatial Awareness. Cortex 2008, 44, 983–995. [Google Scholar] [CrossRef]
- Carretié, L.; Ríos, M.; Periáñez, J.A.; Kessel, D.; Álvarez-Linera, J. The Role of Low and High Spatial Frequencies in Exogenous Attention to Biologically Salient Stimuli. PLoS ONE 2012, 7, e37082. [Google Scholar] [CrossRef]
- Martín-Arévalo, E.; Lupiáñez, J.; Narganes-Pineda, C.; Marino, G.; Colás, I.; Chica, A.B. The Causal Role of the Left Parietal Lobe in Facilitation and Inhibition of Return. Cortex 2019, 117, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Martín-Signes, M.; Pérez-Serrano, C.; Chica, A.B. Causal Contributions of the SMA to Alertness and Consciousness Interactions. Cereb. Cortex 2019, 29, 648–656. [Google Scholar] [CrossRef]
- Quentin, R.; Chanes, L.; Migliaccio, R.; Valabrègue, R.; Valero-Cabré, A. Fronto-Tectal White Matter Connectivity Mediates Facilitatory Effects of Non-Invasive Neurostimulation on Visual Detection. Neuroimage 2013, 82, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Quentin, R.; Elkin Frankston, S.; Vernet, M.; Toba, M.N.; Bartolomeo, P.; Chanes, L.; Valero-Cabré, A. Visual Contrast Sensitivity Improvement by Right Frontal High-Beta Activity Is Mediated by Contrast Gain Mechanisms and Influenced by Fronto-Parietal White Matter Microstructure. Cereb. Cortex 2016, 26, 2381–2390. [Google Scholar] [CrossRef]
- Hong, K.S.; Lee, S.K.; Kim, J.Y.; Kim, K.K.; Nam, H. Visual Working Memory Revealed by Repetitive Transcranial Magnetic Stimulation. J. Neurol. Sci. 2000, 181, 50–55. [Google Scholar] [CrossRef]
- Kessels, R.P.; d’Alfonso, A.A.; Postma, A.; de Haan, E.H. Spatial Working Memory Performance after High-Frequency Repetitive Transcranial Magnetic Stimulation of the Left and Right Posterior Parietal Cortex in Humans. Neurosci. Lett. 2000, 287, 68–70. [Google Scholar] [CrossRef]
- Prime, S.L.; Vesia, M.; Crawford, J.D. Transcranial Magnetic Stimulation over Posterior Parietal Cortex Disrupts Transsaccadic Memory of Multiple Objects. J. Neurosci. 2008, 28, 6938–6949. [Google Scholar] [CrossRef] [PubMed]
- Kastner, S.; Ungerleider, L.G. Mechanisms of Visual Attention in the Human Cortex. Annu. Rev. Neurosci. 2000, 23, 315–341. [Google Scholar] [CrossRef]
- Cavada, C.; Goldman-Rakic, P.S. Posterior Parietal Cortex in Rhesus Monkey: I. Parcellation of Areas Based on Distinctive Limbic and Sensory Corticocortical Connections. J. Comp. Neurol. 1989, 287, 393–421. [Google Scholar] [CrossRef] [PubMed]
- Cavada, C.; Goldman-Rakic, P.S. Posterior Parietal Cortex in Rhesus Monkey: II. Evidence for Segregated Corticocortical Networks Linking Sensory and Limbic Areas with the Frontal Lobe. J. Comp. Neurol. 1989, 287, 422–445. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D.; Pandya, D.N.; Wang, R.; Dai, G.; D’Arceuil, H.E.; de Crespigny, A.J.; Wedeen, V.J. Association Fibre Pathways of the Brain: Parallel Observations from Diffusion Spectrum Imaging and Autoradiography. Brain 2007, 130, 630–653. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical Power Analyses Using G*Power 3.1: Tests for Correlation and Regression Analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Safety of TMS Consensus Group Safety, Ethical Considerations, and Application Guidelines for the Use of Transcranial Magnetic Stimulation in Clinical Practice and Research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.; Eschman, A.; Zuccolotto, A. E-Prime: User’s Guide. Reference Guide. Getting Started Guide; Psychology Software Tools, Incorporated: Sharpsburg, PA, USA, 2002. [Google Scholar]
- Chen, R.; Yung, D.; Li, J.-Y. Organization of Ipsilateral Excitatory and Inhibitory Pathways in the Human Motor Cortex. J. Neurophysiol. 2003, 89, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.G.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.B.; George, M.S.; et al. Non-Invasive Electrical and Magnetic Stimulation of the Brain, Spinal Cord, Roots and Peripheral Nerves: Basic Principles and Procedures for Routine Clinical and Research Application. An Updated Report from an I.F.C.N. Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef] [PubMed]
- Triggs, W.J.; Calvanio, R.; Macdonell, R.A.; Cros, D.; Chiappa, K.H. Physiological Motor Asymmetry in Human Handedness: Evidence from Transcranial Magnetic Stimulation. Brain Res. 1994, 636, 270–276. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Restuccia, D.; Oliviero, A.; Profice, P.; Ferrara, L.; Insola, A.; Mazzone, P.; Tonali, P.; Rothwell, J.C. Magnetic Transcranial Stimulation at Intensities below Active Motor Threshold Activates Intracortical Inhibitory Circuits. Exp. Brain Res. 1998, 119, 265–268. [Google Scholar] [CrossRef]
- Köhler, S.; Paus, T.; Buckner, R.L.; Milner, B. Effects of Left Inferior Prefrontal Stimulation on Episodic Memory Formation: A Two-Stage FMRI-RTMS Study. J. Cogn. Neurosci. 2004, 16, 178–188. [Google Scholar] [CrossRef]
- Feredoes, E.; Tononi, G.; Postle, B.R. Direct Evidence for a Prefrontal Contribution to the Control of Proactive Interference in Verbal Working Memory. Proc. Natl. Acad. Sci. USA 2006, 103, 19530–19534. [Google Scholar] [CrossRef] [PubMed]
- Kincade, J.M.; Abrams, R.A.; Astafiev, S.V.; Shulman, G.L.; Corbetta, M. An Event-Related Functional Magnetic Resonance Imaging Study of Voluntary and Stimulus-Driven Orienting of Attention. J. Neurosci. 2005, 25, 4593–4604. [Google Scholar] [CrossRef] [PubMed]
- Heinen, K.; Ruff, C.C.; Bjoertomt, O.; Schenkluhn, B.; Bestmann, S.; Blankenburg, F.; Driver, J.; Chambers, C.D. Concurrent TMS-FMRI Reveals Dynamic Interhemispheric Influences of the Right Parietal Cortex during Exogenously Cued Visuospatial Attention. Eur. J. Neurosci. 2011, 33, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Harris, I.M.; Benito, C.T.; Ruzzoli, M.; Miniussi, C. Effects of Right Parietal Transcranial Magnetic Stimulation on Object Identification and Orientation Judgments. J. Cogn. Neurosci. 2008, 20, 916–926. [Google Scholar] [CrossRef][Green Version]
- Kalla, R.; Muggleton, N.G.; Cowey, A.; Walsh, V. Human Dorsolateral Prefrontal Cortex Is Involved in Visual Search for Conjunctions but Not Features: A Theta TMS Study. Cortex 2009, 45, 1085–1090. [Google Scholar] [CrossRef]
- Muggleton, N.G.; Cowey, A.; Walsh, V. The Role of the Angular Gyrus in Visual Conjunction Search Investigated Using Signal Detection Analysis and Transcranial Magnetic Stimulation. Neuropsychologia 2008, 46, 2198–2202. [Google Scholar] [CrossRef]
- Ortiz-Tudela, J.; Martín-Arévalo, E.; Chica, A.B.; Lupiáñez, J. Semantic Incongruity Attracts Attention at a Pre-Conscious Level: Evidence from a TMS Study. Cortex 2018, 102, 96–106. [Google Scholar] [CrossRef]
- Dell’Acqua, F.; Simmons, A.; Williams, S.C.R.; Catani, M. Can Spherical Deconvolution Provide More Information than Fiber Orientations? Hindrance Modulated Orientational Anisotropy, a True-Tract Specific Index to Characterize White Matter Diffusion. Hum. Brain Mapp. 2013, 34, 2464–2483. [Google Scholar] [CrossRef]
- Rojkova, K.; Volle, E.; Urbanski, M.; Humbert, F.; Dell’Acqua, F.; Thiebaut de Schotten, M. Atlasing the Frontal Lobe Connections and Their Variability due to Age and Education: A Spherical Deconvolution Tractography Study. Brain Struct. Funct. 2016, 221, 1751–1766. [Google Scholar] [CrossRef] [PubMed]
- Cowan, N. The Magical Number 4 in Short-Term Memory: A Reconsideration of Mental Storage Capacity. Behav. Brain Sci. 2001, 24, 87–114, discussion 114–185. [Google Scholar] [CrossRef]
- Field, A. Discovering Statistics Using IBM SPSS Statistics; SAGE: Newbury Park, CA, USA, 2013; ISBN 1-4462-7458-6. [Google Scholar]
- Miller, J. Reaction Time Analysis with Outlier Exclusion: Bias Varies with Sample Size. Q. J. Exp. Psychol. 1991, 43, 907–912. [Google Scholar] [CrossRef]
- Koerner, T.K.; Zhang, Y. Application of Linear Mixed-Effects Models in Human Neuroscience Research: A Comparison with Pearson Correlation in Two Auditory Electrophysiology Studies. Brain Sci. 2017, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 0-387-87458-5. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. LmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- De Rosario-Martinez, H.; Fox, J.; Team, R.C.; De Rosario-Martinez, M.H. Package ‘Phia’. CRAN Repos. Retrieved 2015, 1, 2015. [Google Scholar]
- Chica, A.B.; Bartolomeo, P.; Valero-Cabré, A. Dorsal and Ventral Parietal Contributions to Spatial Orienting in the Human Brain. J. Neurosci. 2011, 31, 8143–8149. [Google Scholar] [CrossRef]
- Tuch, D.S.; Salat, D.H.; Wisco, J.J.; Zaleta, A.K.; Hevelone, N.D.; Rosas, H.D. Choice Reaction Time Performance Correlates with Diffusion Anisotropy in White Matter Pathways Supporting Visuospatial Attention. Proc. Natl. Acad. Sci. USA 2005, 102, 12212–12217. [Google Scholar] [CrossRef]
- Walhovd, K.B.; Fjell, A.M. White Matter Volume Predicts Reaction Time Instability. Neuropsychologia 2007, 45, 2277–2284. [Google Scholar] [CrossRef] [PubMed]
- Dehaene, S.; Changeux, J.-P.; Naccache, L.; Sackur, J.; Sergent, C. Conscious, Preconscious, and Subliminal Processing: A Testable Taxonomy. Trends Cogn. Sci. 2006, 10, 204–211. [Google Scholar] [CrossRef]
- Kuo, B.-C.; Stokes, M.G.; Murray, A.M.; Nobre, A.C. Attention Biases Visual Activity in Visual Short-Term Memory. J. Cogn. Neurosci. 2014, 26, 1377–1389. [Google Scholar] [CrossRef] [PubMed]
- LaRocque, J.J.; Lewis-Peacock, J.A.; Drysdale, A.T.; Oberauer, K.; Postle, B.R. Decoding Attended Information in Short-Term Memory: An EEG Study. J. Cogn. Neurosci. 2013, 25, 127–142. [Google Scholar] [CrossRef]
- Lewis-Peacock, J.A.; Drysdale, A.T.; Oberauer, K.; Postle, B.R. Neural Evidence for a Distinction between Short-Term Memory and the Focus of Attention. J. Cogn. Neurosci. 2012, 24, 61–79. [Google Scholar] [CrossRef]
- Souza, A.S.; Rerko, L.; Lin, H.-Y.; Oberauer, K. Focused Attention Improves Working Memory: Implications for Flexible-Resource and Discrete-Capacity Models. Atten. Percept. Psychophys. 2014, 76, 2080–2102. [Google Scholar] [CrossRef] [PubMed]
- Kuo, B.-C.; Yeh, Y.-Y.; Chen, A.J.-W.; D’Esposito, M. Functional Connectivity during Top-down Modulation of Visual Short-Term Memory Representations. Neuropsychologia 2011, 49, 1589–1596. [Google Scholar] [CrossRef]
- Nobre, A.C.; Griffin, I.C.; Rao, A. Spatial Attention Can Bias Search in Visual Short-Term Memory. Front. Hum. Neurosci. 2007, 1, 4. [Google Scholar] [CrossRef]
- Rerko, L.; Oberauer, K. Focused, Unfocused, and Defocused Information in Working Memory. J. Exp. Psychol. Learn. Mem. Cogn. 2013, 39, 1075–1096. [Google Scholar] [CrossRef]
- Souza, A.S.; Rerko, L.; Oberauer, K. Getting More from Visual Working Memory: Retro-Cues Enhance Retrieval and Protect from Visual Interference. J. Exp. Psychol. Hum. Percept. Perform. 2016, 42, 890–910. [Google Scholar] [CrossRef] [PubMed]
- Bundesen, C.; Habekost, T.; Kyllingsbaek, S. A Neural Theory of Visual Attention: Bridging Cognition and Neurophysiology. Psychol Rev 2005, 112, 291–328. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, J. From Thought to Action: The Parietal Cortex as a Bridge between Perception, Action, and Cognition. Neuron 2007, 53, 9–16. [Google Scholar] [CrossRef]
- Nardo, D.; Santangelo, V.; Macaluso, E. Stimulus-Driven Orienting of Visuo-Spatial Attention in Complex Dynamic Environments. Neuron 2011, 69, 1015–1028. [Google Scholar] [CrossRef]
- Sergent, C.; Wyart, V.; Babo-Rebelo, M.; Cohen, L.; Naccache, L.; Tallon-Baudry, C. Cueing Attention after the Stimulus Is Gone Can Retrospectively Trigger Conscious Perception. Curr. Biol. 2013, 23, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Postle, B.R.; Awh, E.; Jonides, J.; Smith, E.E.; D’Esposito, M. The Where and How of Attention-Based Rehearsal in Spatial Working Memory. Cogn. Brain Res. 2004, 20, 194–205. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Botta, F.; Lupiáñez, J.; Santangelo, V.; Martín-Arévalo, E. Transcranial Magnetic Stimulation of the Right Superior Parietal Lobule Modulates the Retro-Cue Benefit in Visual Short-Term Memory. Brain Sci. 2021, 11, 252. https://doi.org/10.3390/brainsci11020252
Botta F, Lupiáñez J, Santangelo V, Martín-Arévalo E. Transcranial Magnetic Stimulation of the Right Superior Parietal Lobule Modulates the Retro-Cue Benefit in Visual Short-Term Memory. Brain Sciences. 2021; 11(2):252. https://doi.org/10.3390/brainsci11020252
Chicago/Turabian StyleBotta, Fabiano, Juan Lupiáñez, Valerio Santangelo, and Elisa Martín-Arévalo. 2021. "Transcranial Magnetic Stimulation of the Right Superior Parietal Lobule Modulates the Retro-Cue Benefit in Visual Short-Term Memory" Brain Sciences 11, no. 2: 252. https://doi.org/10.3390/brainsci11020252
APA StyleBotta, F., Lupiáñez, J., Santangelo, V., & Martín-Arévalo, E. (2021). Transcranial Magnetic Stimulation of the Right Superior Parietal Lobule Modulates the Retro-Cue Benefit in Visual Short-Term Memory. Brain Sciences, 11(2), 252. https://doi.org/10.3390/brainsci11020252