
Cathodal Transcranial Direct Current Stimulation (tDCS) Applied to the Left Premotor Cortex Interferes with Explicit Reproduction of a Motor Sequence


Abstract
1. Introduction
2. Materials and Methods
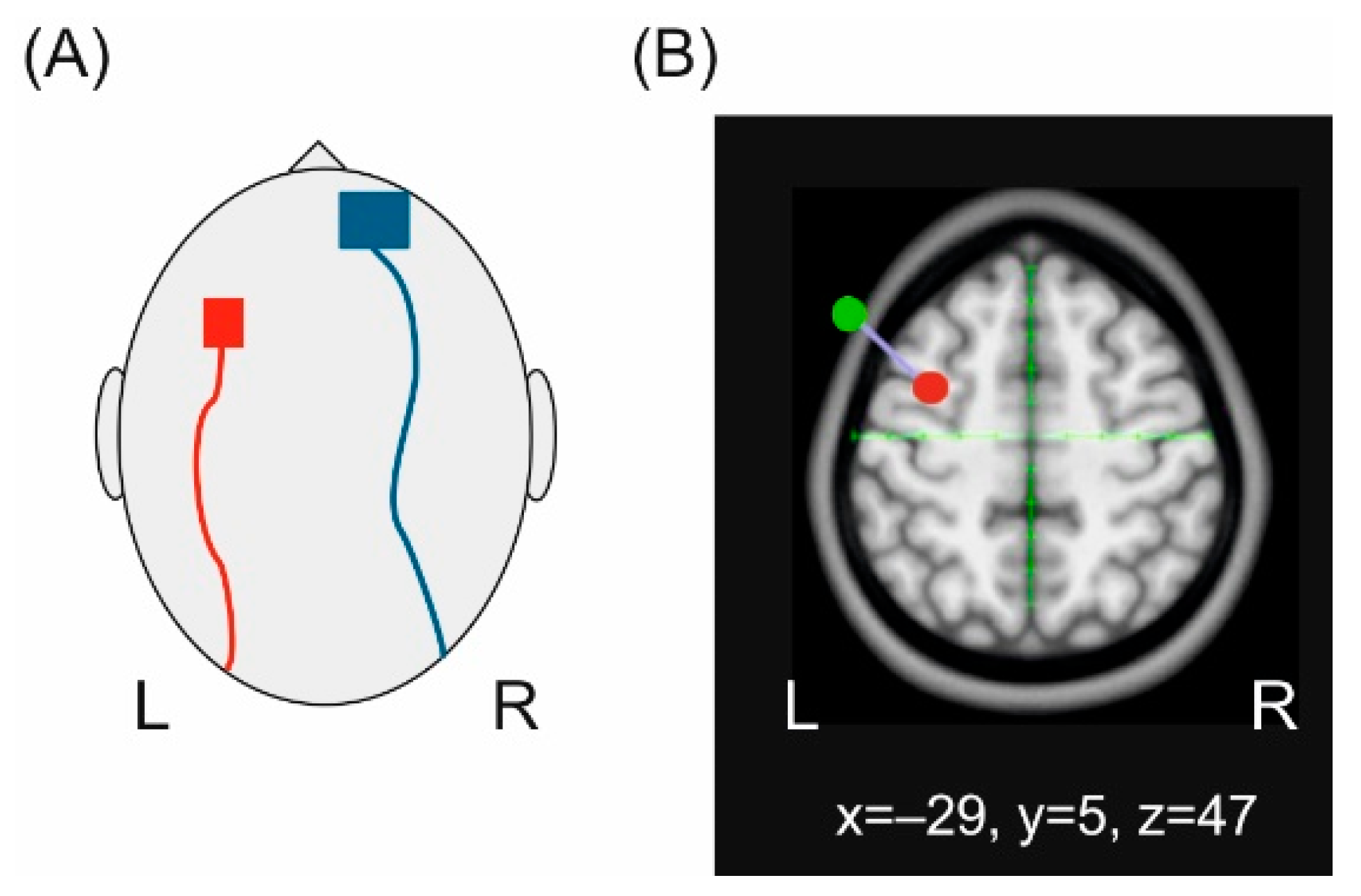
2.1. Transcranial Direct Current Stimulation (tDCS)
2.2. Explicit Motor Sequence Learning
2.3. Design
2.4. Statistics
3. Results and Discussion
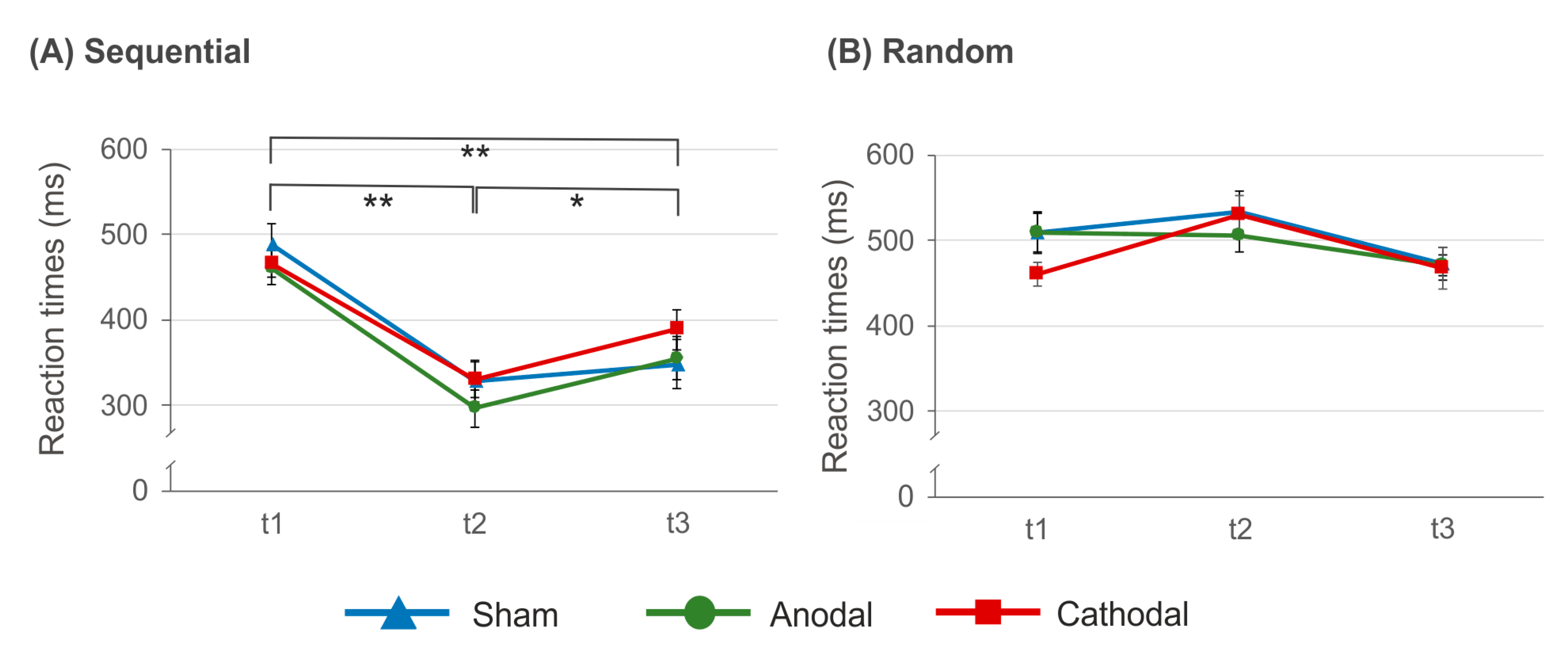
3.1. Reaction Times during SRTT Training
3.2. Reaction Times after SRTT Training
3.3. Sequence Reproduction
3.4. Consolidation during Sleep
3.5. Limitations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nissen, M.J.; Bullemer, P. Attentional Requirements of Learning: Evidence from Performance Measures. Cogn. Psychol. 1987, 19, 1–32. [Google Scholar] [CrossRef]
- Robertson, E.M. The Serial Reaction Time Task: Implicit Motor Skill Learning? J. Neurosci. 2007, 27, 10073–10075. [Google Scholar] [CrossRef]
- Penhune, V.B.; Steele, C.J. Parallel Contributions of Cerebellar, Striatal and M1 Mechanisms to Motor Sequence Learning. Behav. Brain Res. 2012, 226, 579–591. [Google Scholar] [CrossRef]
- Dayan, E.; Cohen, L.G. Neuroplasticity Subserving Motor Skill Learning. Neuron 2011, 72, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Doyon, J.; Bellec, P.; Amsel, R.; Penhune, V.; Monchi, O.; Carrier, J.; Lehéricy, S.; Benali, H. Contributions of the Basal Ganglia and Functionally Related Brain Structures to Motor Learning. Behav. Brain Res. 2009, 199, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Krakauer, J.W.; Hadjiosif, A.M.; Xu, J.; Wong, A.L.; Haith, A.M. Motor Learning. Compr. Physiol. 2019, 9, 613–663. [Google Scholar] [CrossRef] [PubMed]
- Robertson, E.M.; Pascual-Leone, A.; Miall, R.C. Current Concepts in Procedural Consolidation. Nat. Rev. Neurosci. 2004, 5, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Bönstrup, M.; Iturrate, I.; Thompson, R.; Cruciani, G.; Censor, N.; Cohen, L.G. A Rapid Form of Offline Consolidation in Skill Learning. Curr. Biol. 2019, 29, 1346–1351.e4. [Google Scholar] [CrossRef]
- Buch, E.R.; Santarnecchi, E.; Antal, A.; Born, J.; Celnik, P.A.; Classen, J.; Gerloff, C.; Hallett, M.; Hummel, F.C.; Nitsche, M.A.; et al. Effects of tDCS on Motor Learning and Memory Formation: A Consensus and Critical Position Paper. Clin. Neurophysiol. 2017, 128, 589–603. [Google Scholar] [CrossRef]
- Song, S. Consciousness and the Consolidation of Motor Learning. Behav. Brain Res. 2009, 196, 180–186. [Google Scholar] [CrossRef]
- Censor, N.; Sagi, D.; Cohen, L.G. Common Mechanisms of Human Perceptual and Motor Learning. Nat. Rev. Neurosci. 2012, 13, 658–664. [Google Scholar] [CrossRef]
- Janacsek, K.; Nemeth, D. Predicting the Future: From Implicit Learning to Consolidation. Int. J. Psychophysiol. 2012, 83, 213–221. [Google Scholar] [CrossRef]
- Nemeth, D.; Janacsek, K.; Londe, Z.; Ullman, M.T.; Howard, D.V.; Howard, J.H. Sleep Has No Critical Role in Implicit Motor Sequence Learning in Young and Old Adults. Exp. Brain Res. 2009, 201, 351–358. [Google Scholar] [CrossRef]
- Hallgató, E.; Győri-Dani, D.; Pekár, J.; Janacsek, K.; Nemeth, D. The Differential Consolidation of Perceptual and Motor Learning in Skill Acquisition. Cortex 2013, 49, 1073–1081. [Google Scholar] [CrossRef]
- Doyon, J.; Benali, H. Reorganization and Plasticity in the Adult Brain during Learning of Motor Skills. Curr. Opin. Neurobiol. 2005, 15, 161–167. [Google Scholar] [CrossRef]
- Karni, A.; Meyer, G.; Rey-Hipolito, C.; Jezzard, P.; Adams, M.M.; Turner, R.; Ungerleider, L.G. The Acquisition of Skilled Motor Performance: Fast and Slow Experience-Driven Changes in Primary Motor Cortex. Proc. Natl. Acad. Sci. USA 1998, 95, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Papale, A.E.; Hooks, B.M. Circuit Changes in Motor Cortex during Motor Skill Learning. Neuroscience 2018, 368, 283–297. [Google Scholar] [CrossRef] [PubMed]
- King, B.R.; Hoedlmoser, K.; Hirschauer, F.; Dolfen, N.; Albouy, G. Sleeping on the Motor Engram: The Multifaceted Nature of Sleep-Related Motor Memory Consolidation. Neurosci. Biobehav. Rev. 2017, 80, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Robertson, E.M.; Pascual-Leone, A.; Press, D.Z. Awareness Modifies the Skill-Learning Benefits of Sleep. Curr. Biol. 2004, 14, 208–212. [Google Scholar] [CrossRef]
- Kincses, Z.T.; Johansen-Berg, H.; Tomassini, V.; Bosnell, R.; Matthews, P.M.; Beckmann, C.F. Model-Free Characterization of Brain Functional Networks for Motor Sequence Learning Using fMRI. NeuroImage 2008, 39, 1950–1958. [Google Scholar] [CrossRef]
- Hazeltine, E.; Grafton, S.T.; Ivry, R. Attention and Stimulus Characteristics Determine the Locus of Motor—Sequence Encoding. a PET Study. Brain 1997, 120, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Tavor, I.; Botvinik-Nezer, R.; Bernstein-Eliav, M.; Tsarfaty, G.; Assaf, Y. Short-Term Plasticity Following Motor Sequence Learning Revealed by Diffusion Magnetic Resonance Imaging. Hum. Brain Mapp. 2019, 41, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Magon, S.; Pfister, A.; Laura, G.; Lüthi, M.; Papadopoulou, A.; Kappos, L.; Sprenger, T. Short Timescale Modulation of Cortical and Cerebellar Activity in the Early Phase of Motor Sequence Learning: An fMRI Study. Brain Imaging Behav. 2020, 14, 2159–2175. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.J.; Knowlton, B.J.; Chiang, M.C.; Iacoboni, M.; Udompholkul, P.; Wu, A.D. Brain–Behavior Correlates of Optimizing Learning through Interleaved Practice. NeuroImage 2011, 56, 1758–1772. [Google Scholar] [CrossRef]
- Wymbs, N.F.; Grafton, S.T. The Human Motor System Supports Sequence-Specific Representations over Multiple Training-Dependent Timescales. Cereb. Cortex 2015, 25, 4213–4225. [Google Scholar] [CrossRef]
- Grafton, S.T.; Hazeltine, E.; Ivry, R. Functional Mapping of Sequence Learning in Normal Humans. J. Cogn. Neurosci. 1995, 7, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Grafton, S.T.; Mazziotta, J.C.; Presty, S.; Friston, K.J.; Frackowiak, R.S.E.; Phelps, M. Functional Anatomy of Human Procedural Learning Determined with Regional Cerebral Blood Flow and PET. J. Neurosci. 1992, 12, 2542–2548. [Google Scholar] [CrossRef]
- Orban, P.; Peigneux, P.; Lungu, O.; Albouy, G.; Breton, E.; Laberenne, F.; Benali, H.; Maquet, P.; Doyon, J. The Multifaceted Nature of the Relationship between Performance and Brain Activity in Motor Sequence Learning. NeuroImage 2010, 49, 694–702. [Google Scholar] [CrossRef]
- Jenkins, I.; Brooks, D.; Nixon, P.; Frackowiak, R.; Passingham, R. Motor Sequence Learning: A Study with Positron Emission Tomography. J. Neurosci. 1994, 14, 3775–3790. [Google Scholar] [CrossRef]
- Hikosaka, O.; Nakamura, K.; Sakai, K.; Nakahara, H. Central Mechanisms of Motor Skill Learning. Curr. Opin. Neurobiol. 2002, 12, 217–222. [Google Scholar] [CrossRef]
- Ashe, J.; Lungu, O.V.; Basford, A.T.; Lu, X. Cortical Control of Motor Sequences. Curr. Opin. Neurobiol. 2006, 16, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, R.M.; Rottschy, C.; Miall, R.C.; Eickhoff, S.B. A Quantitative Meta-Analysis and Review of Motor Learning in the Human Brain. NeuroImage 2013, 67, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Lohse, K.R.; Wadden, K.; Boyd, L.A.; Hodges, N.J. Motor Skill Acquisition across Short and Long Time Scales: A Meta-Analysis of Neuroimaging Data. Neuropsychol 2014, 59, 130–141. [Google Scholar] [CrossRef]
- Schubotz, R.I.; Von Cramon, D. Functional–Anatomical Concepts of Human Premotor Cortex: Evidence from fMRI and PET Studies. NeuroImage 2003, 20, S120–S131. [Google Scholar] [CrossRef] [PubMed]
- Toni, I.; Krams, M.; Turner, R.; Passingham, R.E. The Time Course of Changes during Motor Sequence Learning: A Whole-Brain fMRI Study. NeuroImage 1998, 8, 50–61. [Google Scholar] [CrossRef]
- Vidoni, E.D.; Boyd, L.A. Achieving Enlightenment: What Do We Know About the Implicit Learning System and Its Interaction With Explicit Knowledge? J. Neurol. Phys. Ther. 2007, 31, 145–154. [Google Scholar] [CrossRef]
- Sami, S.; Robertson, E.M.; Miall, R.C. The Time Course of Task-Specific Memory Consolidation Effects in Resting State Networks. J. Neurosci. 2014, 34, 3982–3992. [Google Scholar] [CrossRef]
- Shadmehr, R. Neural Correlates of Motor Memory Consolidation. Science 1997, 277, 821–825. [Google Scholar] [CrossRef]
- Willingham, D.B.; Salidis, J.; Gabrieli, J.D. Direct Comparison of Neural Systems Mediating Conscious and Unconscious Skill Learning. J. Neurophysiol. 2002, 88, 1451–1460. [Google Scholar] [CrossRef]
- Honda, M.; Deiber, M.P.; Ibáñez, V.; Pascual-Leone, A.; Zhuang, P.; Hallett, M. Dynamic Cortical Involvement in Implicit and Explicit Motor Sequence Learning. A PET Study. Brain 1998, 121, 2159–2173. [Google Scholar] [CrossRef] [PubMed]
- Rauch, S.L.; Savage, C.R.; Brown, H.D.; Curran, T.; Alpert, N.M.; Kendrick, A.; Fischman, A.J.; Kosslyn, S.M. A PET Investigation of Implicit and Explicit Sequence Learning. Hum. Brain Mapp. 1995, 3, 271–286. [Google Scholar] [CrossRef]
- Stagg, C.J.; Antal, A.; Nitsche, M.A. Physiology of Transcranial Direct Current Stimulation. J. ECT 2018, 34, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-Z.; Lu, M.-K.; Antal, A.; Classen, J.; Nitsche, M.; Ziemann, U.; Ridding, M.; Hamada, M.; Ugawa, Y.; Jaberzadeh, S.; et al. Plasticity Induced by Non-invasive Transcranial Brain Stimulation: A Position Paper. Clin. Neurophysiol. 2017, 128, 2318–2329. [Google Scholar] [CrossRef] [PubMed]
- Zaghi, S.; Acar, M.; Hultgren, B.; Boggio, P.S.; Fregni, F. Noninvasive Brain Stimulation with Low-Intensity Electrical Currents: Putative Mechanisms of Action for Direct and Alternating Current Stimulation. Neuroscience 2009, 16, 285–307. [Google Scholar] [CrossRef] [PubMed]
- Stagg, C.J.; Best, J.G.; Stephenson, M.C.; O’Shea, J.; Wylezinska, M.; Kincses, Z.T.; Morris, P.G.; Matthews, P.M.; Johansen-Berg, H. Polarity-Sensitive Modulation of Cortical Neurotransmitters by Transcranial Stimulation. J. Neurosci. 2009, 29, 5202–5206. [Google Scholar] [CrossRef] [PubMed]
- Bütefisch, C.M.; Davis, B.C.; Wise, S.P.; Sawaki, L.; Kopylev, L.; Classen, J.; Cohen, L.G. Mechanisms of Use-Dependent Plasticity in the Human Motor Cortex. Proc. Natl. Acad. Sci. USA 2000, 97, 3661–3665. [Google Scholar] [CrossRef]
- Stagg, C.J.; Bachtiar, V.; Johansen-Berg, H. The Role of GABA in Human Motor Learning. Curr. Biol. 2011, 21, 480–484. [Google Scholar] [CrossRef]
- Kim, S.; Stephenson, M.C.; Morris, P.G.; Jackson, S.R. tDCS-Induced Alterations in GABA Concentration within Primary Motor Cortex Predict Motor Learning and Motor Memory: A 7T Magnetic Resonance Spectroscopy Study. NeuroImage 2014, 99, 237–243. [Google Scholar] [CrossRef]
- Bortoletto, M.; Pellicciari, M.C.; Rodella, C.; Miniussi, C. The Interaction with Task-induced Activity is More Important Than Polarization: A tDCS Study. Brain Stimul. 2015, 8, 269–276. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Schauenburg, A.; Lang, N.; Liebetanz, D.; Exner, C.; Paulus, W.; Tergau, F. Facilitation of Implicit Motor Learning by Weak Transcranial Direct Current Stimulation of the Primary Motor Cortex in the Human. J. Cogn. Neurosci. 2003, 15, 619–626. [Google Scholar] [CrossRef]
- Antal, A.A.; Nitsche, M.; Kincses, T.Z.; Kruse, W.; Hoffmann, K.-P.; Paulus, W. Facilitation of Visuo-Motor Learning by Transcranial Direct Current Stimulation of the Motor and Extrastriate Visual Areas in Humans. Eur. J. Neurosci. 2004, 19, 2888–2892. [Google Scholar] [CrossRef]
- Stagg, C.; Jayaram, G.; Pastor, D.; Kincses, Z.; Matthews, P.; Johansen-Berg, H. Polarity and Timing-Dependent Effects of Transcranial Direct Current Stimulation in Explicit Motor Learning. Neuropsychology 2011, 49, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, F.; Zappasodi, F.; Assenza, G.; Tombini, M.; Vollaro, S.; Barbati, G.; Rossini, P.M. Anodal Transcranial Direct Current Stimulation Enhances Procedural Consolidation. J. Neurophysiol. 2010, 104, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Reis, J.; Robertson, E.M.; Krakauer, J.W.; Rothwell, J.; Marshall, L.; Gerloff, C.; Wassermann, E.M.; Pascual-Leone, A.; Hummel, F.; Celnik, P.A.; et al. Consensus: Can Transcranial Direct Current Stimulation and Transcranial Magnetic Stimulation Enhance Motor Learning and Memory for-Mation? Brain Stimul. 2008, 1, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Reis, J.; Fritsch, B. Modulation of Motor Performance and Motor Learning by Transcranial Direct Current Stimulation. Curr. Opin. Neurol. 2011, 24, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.; Antal, A.; Bikson, M.; Boggio, P.; Brunoni, A.; Celnik, P.; Cohen, L.; Fregni, F.; Herrmann, C.; Kappenman, E.; et al. A Technical Guide to tDCS, and Related Non-invasive Brain Stimulation Tools. Clin. Neurophysiol. 2016, 127, 1031–1048. [Google Scholar] [CrossRef]
- Morya, E.; Monte-Silva, K.; Bikson, M.; Esmaeilpour, Z.; Biazoli, C.E.; Fonseca, A.; Bocci, T.; Farzan, F.; Chatterjee, R.; Hausdorff, J.M.; et al. Beyond the Target Area: An Integrative View of tDCS-Induced Motor Cortex Modulation in Patients and Athletes. J. Neuroeng. Rehabil. 2019, 16, 141. [Google Scholar] [CrossRef]
- Boyd, A.L.; Linsdell, A.M. Excitatory Repetitive Transcranial Magnetic Stimulation to Left Dorsal Premotor Cortex Enhances Motor Consolidation of New Skills. BMC Neurosci. 2009, 10, 72. [Google Scholar] [CrossRef]
- Meehan, S.K.; Zabukovec, J.R.; Dao, E.; Cheung, K.L.; Linsdell, M.A.; Boyd, L.A. One Hertz Repetitive Transcranial Magnetic Stimulation over Dorsal Premotor Cortex Enhances Offline Motor Memory Consoli-Dation for Sequence-Specific Implicit Learning. Eur. J. Neurosci. 2013, 38, 3071–3079. [Google Scholar] [CrossRef]
- Kantak, S.S.; Mummidisetty, C.K.; Stinear, J.W. Primary Motor and Premotor Cortex in Implicit Sequence Learning - Evidence for Competition between Implicit and Explicit Human Motor Memory Systems. Eur. J. Neurosci. 2012, 36, 2710–2715. [Google Scholar] [CrossRef] [PubMed]
- Focke, J.; Kemmet, S.; Krause, V.; Keitel, A.; Pollok, B. Cathodal Transcranial Direct Current Stimulation (tDCS) Applied to the Left Premotor Cortex (PMC) Stabilizes a Newly Learned Motor Sequence. Behav. Brain Res. 2017, 316, 87–93. [Google Scholar] [CrossRef]
- Wymbs, N.F.; Grafton, S.T. Contributions from the Left PMd and the SMA during Sequence Retrieval as Determined by Depth of Training. Exp. Brain Res. 2012, 224, 49–58. [Google Scholar] [CrossRef][Green Version]
- Kantak, S.S.; Winstein, C.J. Learning–Performance Distinction and Memory Processes for Motor Skills: A Focused Review and Perspective. Behav. Brain Res. 2012, 228, 219–231. [Google Scholar] [CrossRef]
- Oldfield, R. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychology 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Serrien, D.J.; Ivry, R.B.; Swinnen, S.P. Dynamics of Hemispheric Specialization and Integration in the Context of Motor Control. Nat. Rev. Neurosci. 2006, 7, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Paulus, W. Excitability Changes Induced in the Human Motor Cortex by Weak Transcranial Direct Current Stimulation. J. Physiol. 2000, 527, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Doemkes, S.; Karaköse, T.; Antal, A.; Liebetanz, D.; Lang, N.; Tergau, F.; Paulus, W. Shaping the Effects of Transcranial Direct Current Stimulation of the Human Motor Cortex. J. Neurophysiol. 2007, 97, 3109–3117. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Cohen, L.G.; Wassermann, E.M.; Priori, A.; Lang, N.; Antal, A.; Paulus, W.; Hummel, F.; Boggio, P.S.; Fregni, F.; et al. Transcranial Direct Current Stimulation: State of the Art 2008. Brain Stimul. 2008, 1, 206–223. [Google Scholar] [CrossRef]
- Bastani, A.; Jaberzadeh, S. a-tDCS Differential Modulation of Corticospinal Excitability: The Effects of Electrode Size. Brain Stimul. 2013, 6, 932–937. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Liebetanz, D.; Lang, N.; Antal, A.; Tergau, F.; Paulus, W. Safety Criteria for Transcranial Direct Current Stimulation (tDCS) in Humans. Clin. Neurophysiol. 2003, 114, 2220–2222. [Google Scholar] [CrossRef]
- Antal, A.; Alekseichuk, I.; Bikson, M.; Brockmöller, J.; Brunoni, A.; Chen, G.R.; Cohen, L.; Dowthwaite, G.; Ellrich, J.; Flöel, A.; et al. Low Intensity Transcranial Electric Stimulation: Safety, Ethical, Legal Regulatory and Application Guidelines. Clin. Neurophysiol. 2017, 128, 1774–1809. [Google Scholar] [CrossRef]
- Bikson, M.; Grossman, P.; Thomas, C.; Zannou, A.L.; Jiang, J.; Adnan, T.; Mourdoukoutas, A.P.; Kronberg, G.; Truong, D.; Boggio, P.; et al. Safety of Transcranial Direct Current Stimulation: Evidence Based Update 2016. Brain Stimul. 2016, 9, 641–661. [Google Scholar] [CrossRef] [PubMed]
- Saturnino, G.B.; Puonti, O.; Nielsen, J.D.; Antonenko, D.; Madsen, K.H.; Thielscher, A. SimNIBS 2.1: A Comprehensive Pipe-line for Individualized Electric Field Modelling for Transcranial Brain Stimulation. In Brain and Human Body Modeling: Compu-tational Human Modeling at EMBC 2018; Makarov, S., Horner, M., Noetscher, G., Eds.; Springer: Cham, Switzerland, 2019; pp. 3–25. [Google Scholar] [CrossRef]
- Pollok, B.; Keitel, A.; Foerster, M.; Moshiri, G.; Otto, K.; Krause, V. The Posterior Parietal Cortex Mediates Early Offline-Rather Than Online-Motor Sequence Learning. Neuropsychology 2020, 146, 107555. [Google Scholar] [CrossRef] [PubMed]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Gerschlager, W.; Siebner, H.R.; Rothwell, J.C. Decreased Corticospinal Excitability after Subthreshold 1 Hz rTMS over Lateral Premotor Cortex. Neurology 2001, 57, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.-J.; Cao, N.; Zhang, J.; Pi, Y.-L. Intermittent Theta Burst Stimulation Facilitates Functional Connectivity from the Dorsal Premotor Cortex to Primary Motor Cortex. PeerJ 2020, 8, e9253. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical Power Analyses Using G*Power 3.1: Tests for Correlation and Regression Analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, A.M.; Jakoubkova, M.; Thirugnanasambandam, N.; Schmalfuss, L.; Hullemann, S.; Sonka, K.; Paulus, W.; Trenkwalder, C.; Happe, S. Contribution of the Premotor Cortex to Consolidation of Motor Sequence Learning in Humans During Sleep. J. Neurophysiol. 2010, 104, 2603–2614. [Google Scholar] [CrossRef]
- Yokoi, A.; Arbuckle, S.A.; Diedrichsen, J. The Role of Human Primary Motor Cortex in the Production of Skilled Finger Sequences. J. Neurosci. 2018, 38, 1430–1442. [Google Scholar] [CrossRef]
- Yokoi, A.; Diedrichsen, J. Neural Organization of Hierarchical Motor Sequence Representations in the Human Neocortex. Neuron 2019, 103, 1178–1190.e7. [Google Scholar] [CrossRef]
- Kornysheva, K.; Diedrichsen, J. Human Premotor Areas Parse Sequences into Their Spatial and Temporal Features. eLife 2014, 3, e03043. [Google Scholar] [CrossRef] [PubMed]
- Rumpf, J.-J.; Wegscheider, M.; Hinselmann, K.; Fricke, C.; King, B.R.; Weise, D.; Klann, J.; Binkofski, F.; Buccino, G.; Karni, A.; et al. Enhancement of Motor Consolidation by Post-training Transcranial Direct Current Stimulation in Older People. Neurobiol. Aging 2017, 49, 1–8. [Google Scholar] [CrossRef]
- Janacsek, K.; Shattuck, K.F.; Tagarelli, K.M.; Lum, J.A.; Turkeltaub, P.E.; Ullman, M.T. Sequence Learning in the Human Brain: A Functional Neuroanatomical Meta-Analysis of Serial Reaction Time Studies. NeuroImage 2020, 207, 116387. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Penhune, V.B.; Zatorre, R.J. The Role of Auditory and Premotor Cortex in Sensorimotor Transformations. Ann. New York Acad. Sci. 2009, 1169, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Nitschke, M.F.; Melchert, U.H.; Erdmann, C.; Born, J. Motor Memory Consolidation in Sleep Shapes More Effective Neuronal Representations. J. Neurosci. 2005, 25, 11248–11255. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Hallschmid, M.; Elsner, A.L.; Born, J. Sleep forms memory for finger skills. Proc. Natl. Acad. Sci. USA 2002, 99, 11987–11991. [Google Scholar] [CrossRef] [PubMed]
- Bramham, C.R.; Srebro, B. Synaptic Plasticity in the Hippocampus Is Modulated by Behavioral State. Brain Res. 1989, 493, 74–86. [Google Scholar] [CrossRef]
- Walker, M.P.; Brakefield, T.; Morgan, A.; Hobson, J.A.; Stickgold, R. Practice with Sleep Makes Perfect. Neuron 2002, 35, 205–211. [Google Scholar] [CrossRef]
- Lauro, L.J.R.; Rosanova, M.; Mattavelli, G.; Convento, S.; Pisoni, A.; Opitz, A.; Bolognini, N.; Vallar, G. TDCS Increases Cortical Excitability: Direct Evidence From TMS–EEG. Cortex 2014, 58, 99–111. [Google Scholar] [CrossRef]
- Bolzoni, F.; Pettersson, L.-G.; Jankowska, E. Evidence for Long-Lasting Subcortical Facilitation by Transcranial Direct Current Stimulation in the Cat. J. Physiol. 2013, 591, 3381–3399. [Google Scholar] [CrossRef]
- Bolzoni, F.; Bączyk, M.; Jankowska, E. Subcortical Effects of Transcranial Direct Current Stimulation in the Rat. J. Physiol. 2013, 591, 4027–4042. [Google Scholar] [CrossRef] [PubMed]
- Bączyk, M.; Jankowska, E. Presynaptic Actions of Transcranial and Local Direct Current Stimulation in the Red Nucleus. J. Physiol. 2014, 592, 4313–4328. [Google Scholar] [CrossRef] [PubMed]
- Batsikadze, G.; Moliadze, V.; Paulus, W.; Kuo, M.-F.; Nitsche, M.A. Partially Non-linear Stimulation Intensity-Dependent Effects of Direct Current Stimulation on Motor Cortex Excitability in Humans. J. Physiol. 2013, 591, 1987–2000. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participants | Reports | ||
|---|---|---|---|
| Sham | Anodal | Cathodal | |
| 01 | Slightly increased tonicity of jaw muscles | None | Flicker sensation left eye at the end of stimulation |
| 02 | Slight tingling below electrodes | Slight tingling below electrodes | Slight itching below frontal electrode |
| 03 | None | None | None |
| 04 | None | None | None |
| 05 | None | None | None |
| 06 | None | None | None |
| 07 | Slight tingling below electrodes | None | None |
| 08 | None | None | None |
| 09 | None | Slight tingling below electrodes | Slight tingling below frontal electrode |
| 10 | Slight tingling below electrodes | Slight tingling below electrodes | None |
| 11 | Warmness below electrodes | None | None |
| 12 | None | None | Pulsation below electrodes |
| 13 | None | None | Slight tingling below electrodes |
| 14 | Slight tingling below electrodes | None | None |
| 15 | None | Slight tingling below electrodes | Slight tingling below electrodes |
| 16 | None | Slight tingling/itching below electrodes | Slight tingling below electrodes A single twitch of the leg |
| 17 | None | None | None |
| 18 | None | None | None |
| (A) Using Reproduction 1 as Criterion (Number of Correctly Reproduced items) | ||||||
| Predictor | b | beta | SE | T | p | |
| Sham | Intercept | 10.112 | 2.531 | 3.995 | 0.002 | |
| T1 (ms) | −0.001 | −0.028 | 0.006 | −0.092 | 0.928 | |
| T2 (ms) | −0.007 | −0.383 | 0.006 | −1.272 | 0.226 | |
| Anodal | Intercept | 13.478 | 3.015 | 4.470 | 0.001 | |
| T1 (ms) | −0.015 | −0.572 | 0.009 | −1.549 | 0.145 | |
| T2 (ms) | 0.002 | 0.098 | 0.008 | 0.266 | 0.794 | |
| Cathodal | Intercept | 5.733 | 7.806 | 0.734 | 0.477 | |
| T1 (ms) | 0.008 | 0.124 | 0.017 | 0.433 | 0.673 | |
| T2 (ms) | −0.008 | −0.262 | 0.009 | −0.912 | 0.380 | |
| (B) Using Reproduction 2 as Criterion (Number of Correctly Reproduced Items) | ||||||
| Predictor | b | beta | SE | T | p | |
| Sham | Intercept | 3.190 | 4.729 | 0.675 | 0.512 | |
| T1 (ms) | 0.004 | 0.112 | 0.011 | 0.343 | 0.737 | |
| T2 (ms) | 0.000 | 0.012 | 0.011 | 0.037 | 0.971 | |
| Anodal | Intercept | 5.078 | 4.349 | 1.168 | 0.264 | |
| T1 (ms) | −0.005 | −0.147 | 0.014 | −0.362 | 0.723 | |
| T2 (ms) | 0.011 | 0.397 | 0.012 | 0.975 | 0.347 | |
| Cathodal | Intercept | 16.376 | 5.314 | 3.082 | 0.010 | |
| T1 (ms) | −0.012 | −0.229 | 0.012 | −1.002 | 0.336 | |
| T2 (ms) | −0.015 | −0.547 | 0.006 | −2.396 | 0.034 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pollok, B.; Schmitz-Justen, C.; Krause, V. Cathodal Transcranial Direct Current Stimulation (tDCS) Applied to the Left Premotor Cortex Interferes with Explicit Reproduction of a Motor Sequence. Brain Sci. 2021, 11, 207. https://doi.org/10.3390/brainsci11020207
Pollok B, Schmitz-Justen C, Krause V. Cathodal Transcranial Direct Current Stimulation (tDCS) Applied to the Left Premotor Cortex Interferes with Explicit Reproduction of a Motor Sequence. Brain Sciences. 2021; 11(2):207. https://doi.org/10.3390/brainsci11020207
Chicago/Turabian StylePollok, Bettina, Claire Schmitz-Justen, and Vanessa Krause. 2021. "Cathodal Transcranial Direct Current Stimulation (tDCS) Applied to the Left Premotor Cortex Interferes with Explicit Reproduction of a Motor Sequence" Brain Sciences 11, no. 2: 207. https://doi.org/10.3390/brainsci11020207
APA StylePollok, B., Schmitz-Justen, C., & Krause, V. (2021). Cathodal Transcranial Direct Current Stimulation (tDCS) Applied to the Left Premotor Cortex Interferes with Explicit Reproduction of a Motor Sequence. Brain Sciences, 11(2), 207. https://doi.org/10.3390/brainsci11020207