
Endogenous Oxytocin Levels in Autism—A Meta-Analysis

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
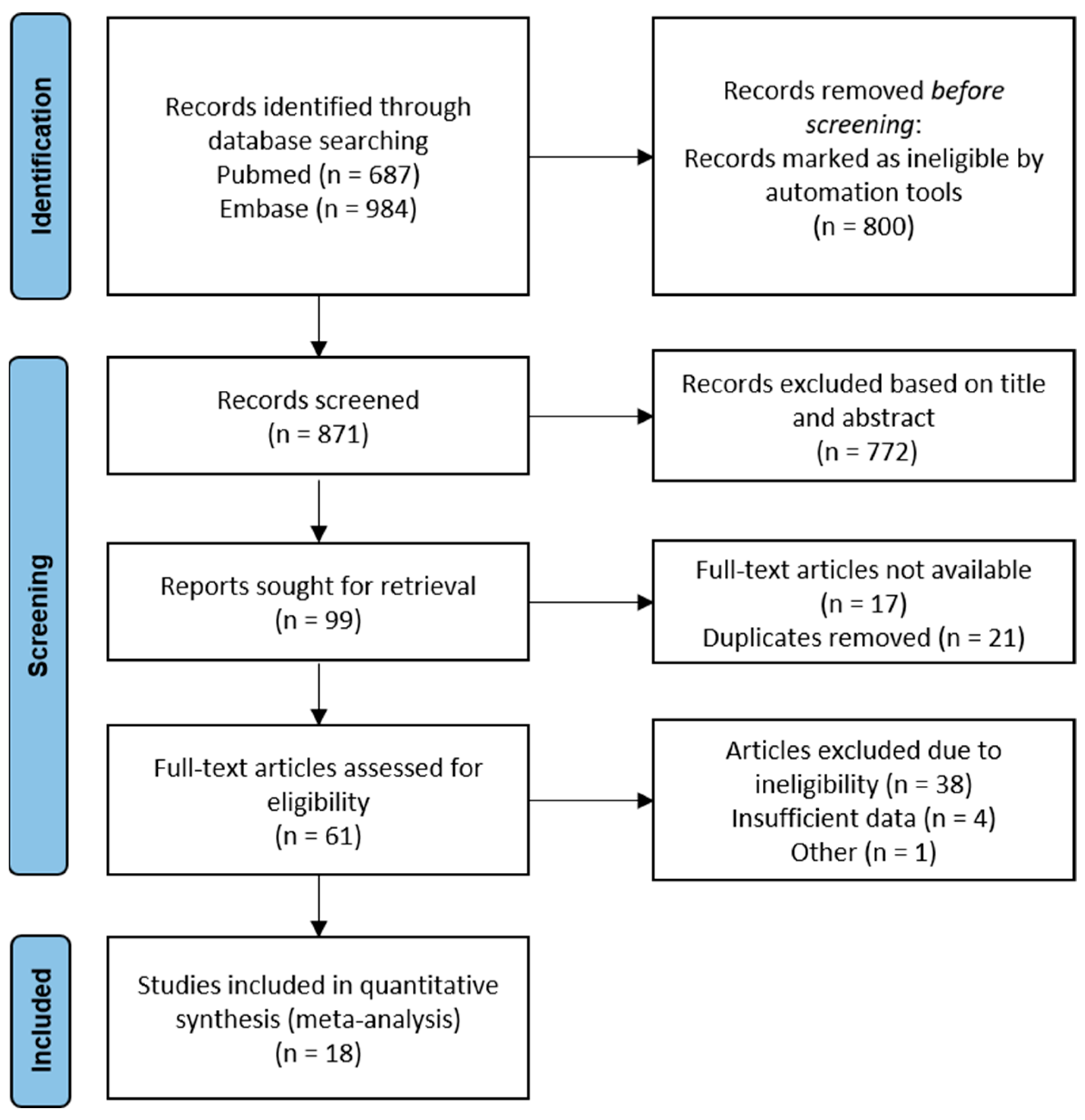
3.1. Study Selection and Characteristics
3.2. Qualitative Risk of Bias Analysis of Included Studies
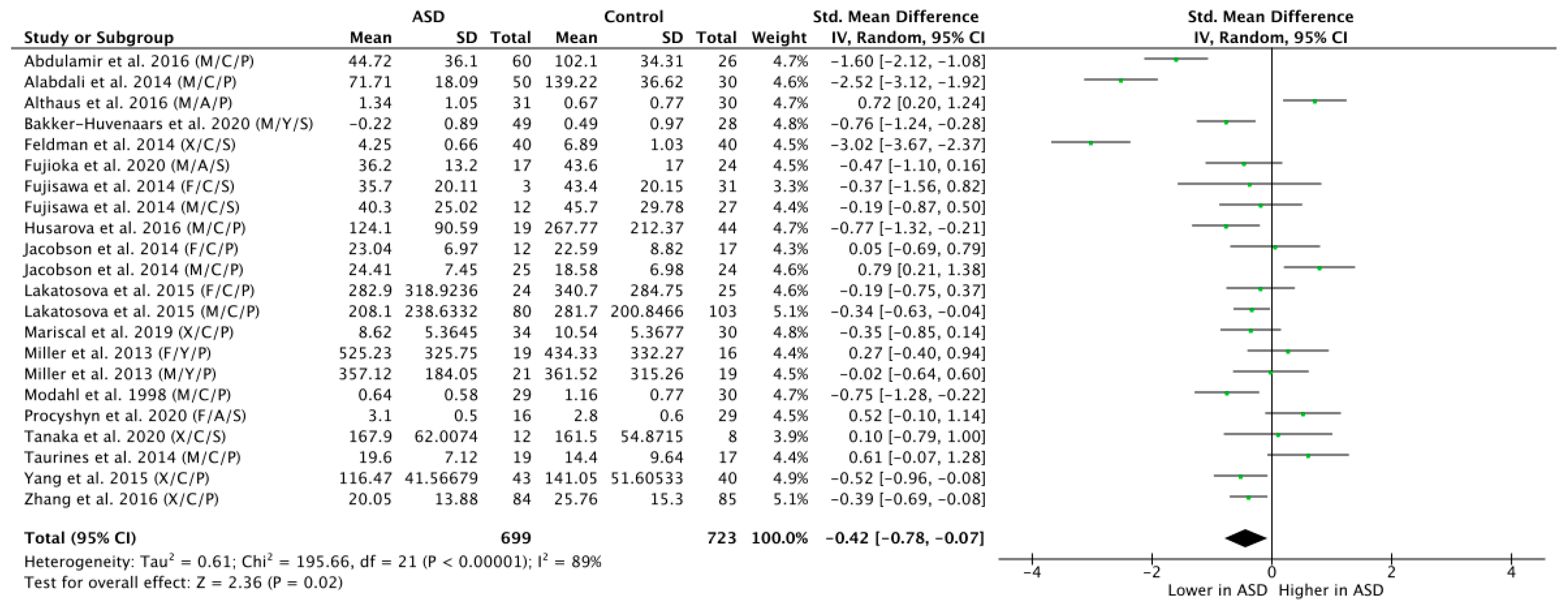
3.3. Meta-Analysis of Peripheral OT Levels in ASD vs. NT Controls
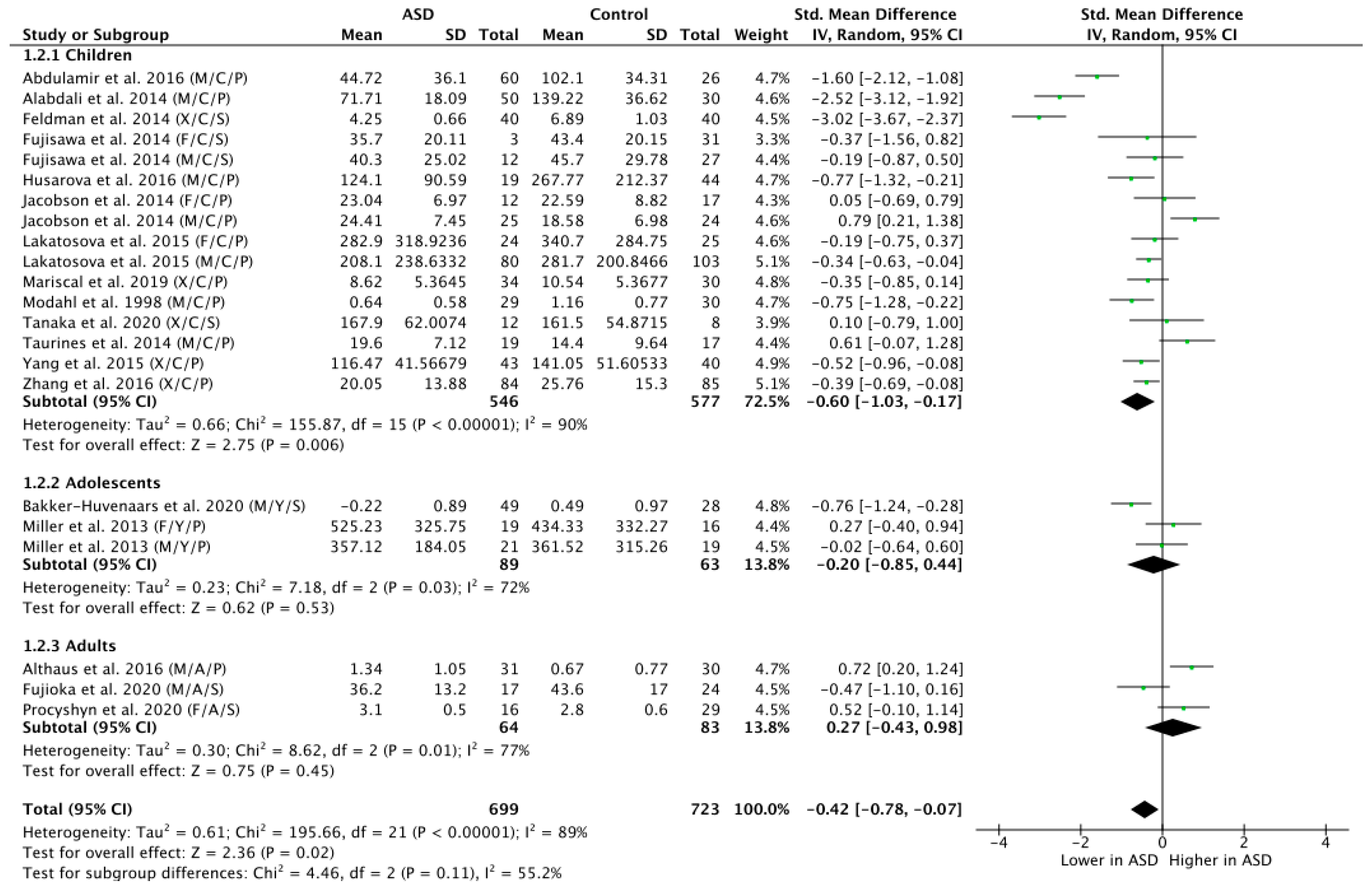
3.4. Subgroup Analysis by Age
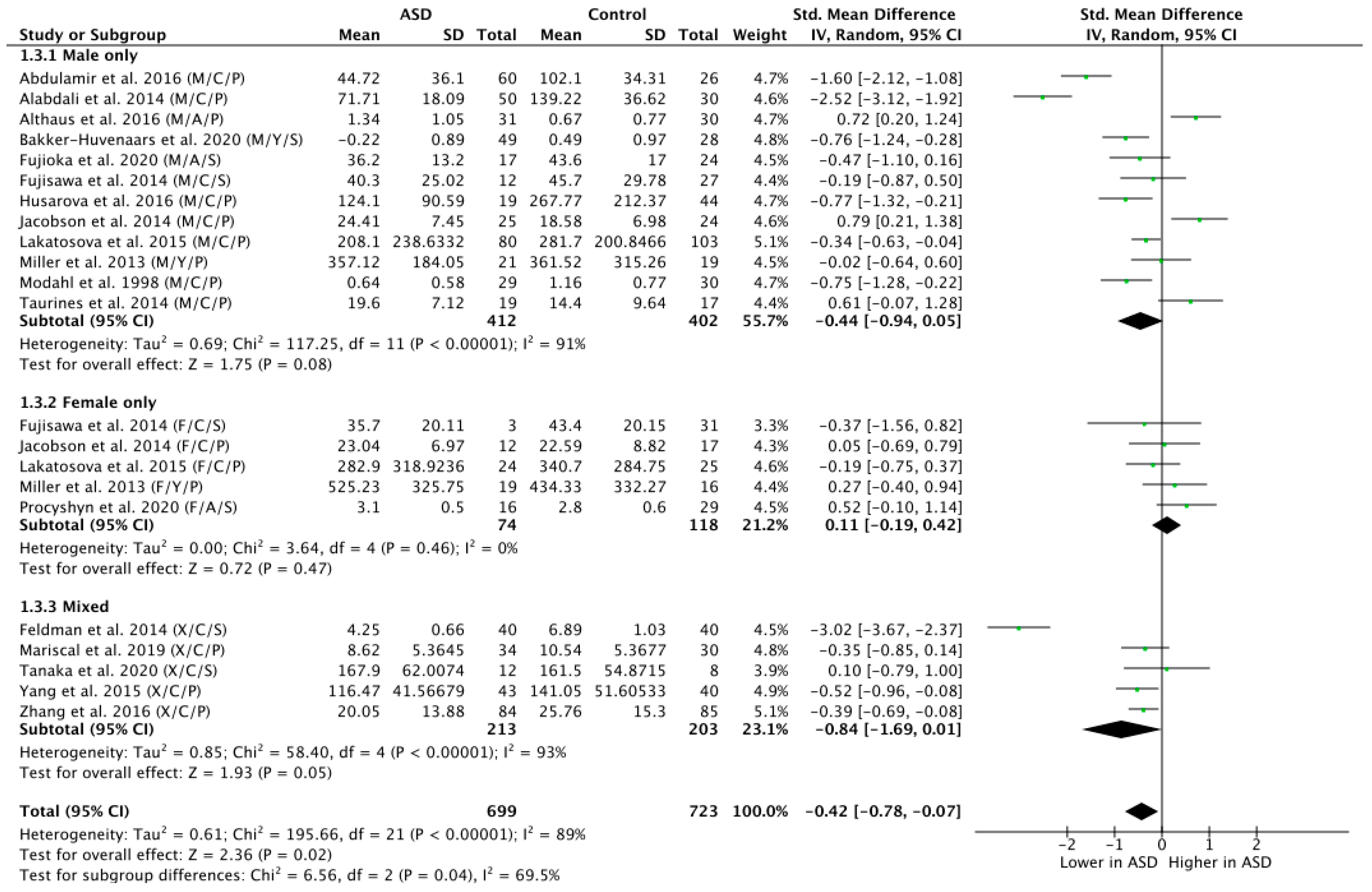
3.5. Subgroup Analysis by Sex
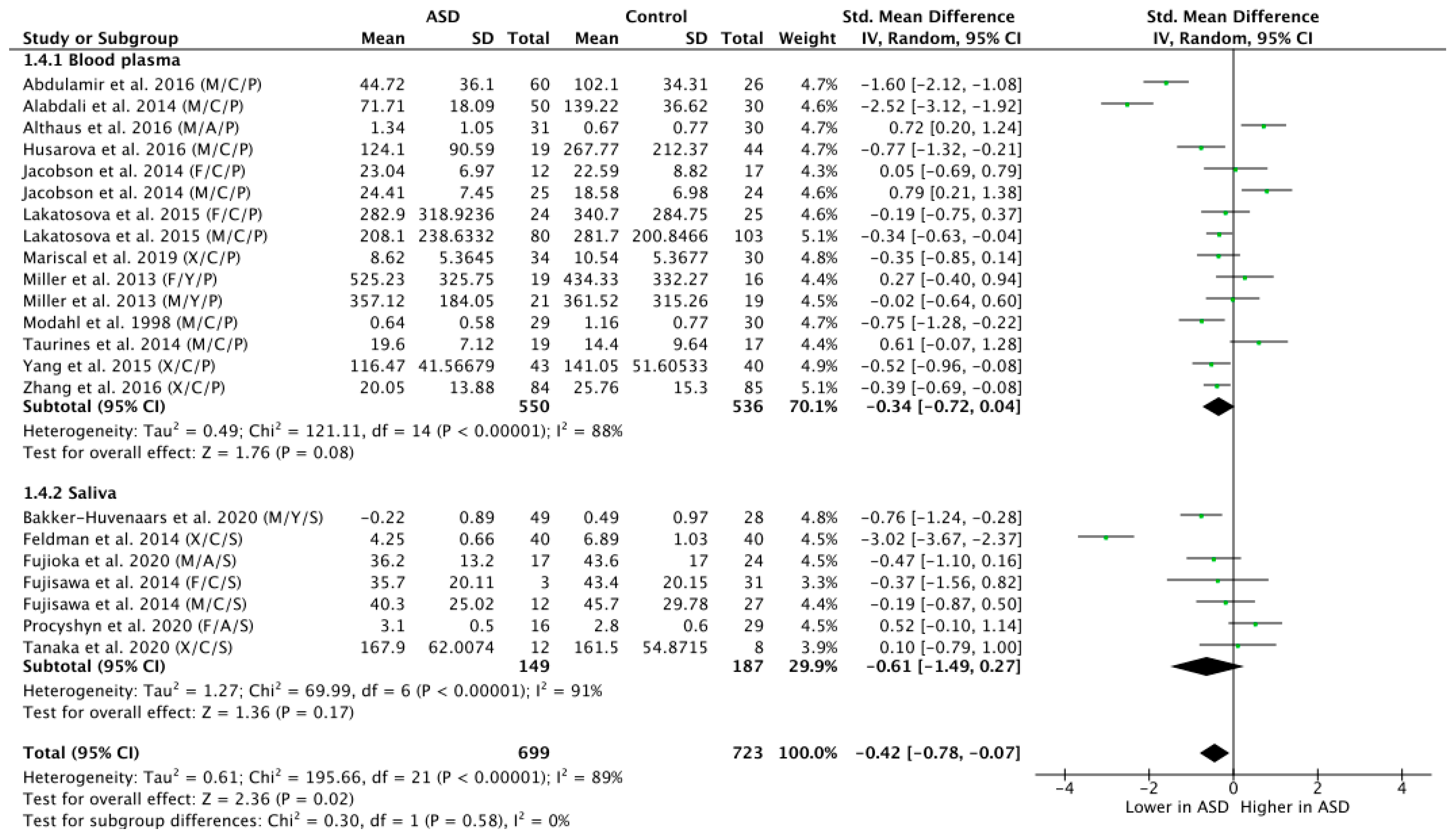
3.6. Subgroup Analysis by Tissue Sample
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Loomes, R.; Hull, L.; Mandy, W.P.L. What Is the Male-to-Female Ratio in Autism Spectrum Disorder? A Systematic Review and Meta-Analysis. J. Am. Acad. Child. Adolesc. Psychiatry 2017, 56, 466–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.-F.; Dai, Y.-C.; Wu, J.; Jia, M.-X.; Zhang, J.-S.; Shou, X.-J.; Han, S.-P.; Zhang, R.; Han, J.-S. Plasma Oxytocin and Arginine-Vasopressin Levels in Children with Autism Spectrum Disorder in China: Associations with Symptoms. Neurosci. Bull. 2016, 32, 423–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdulamir, H.A.; Abdul-Rasheed, O.F.; Abdulghani, E.A. Low oxytocin and melatonin levels and their possible role in the diagnosis and prognosis in Iraqi autistic children. Saudi Med. J. 2016, 37, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Alabdali, A.; Al-Ayadhi, L.; El-Ansary, A. Association of social and cognitive impairment and biomarkers in autism spectrum disorders. J. Neuroinflamm. 2014, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Feldman, R.; Golan, O.; Hirschler-Guttenberg, Y.; Ostfeld-Etzion, S.; Zagoory-Sharon, O. Parent-child interaction and oxytocin production in pre-schoolers with autism spectrum disorder. Br. J. Psychiatry 2014, 205, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Modahl, C.; Green, L.A.; Fein, D.; Morris, M.; Waterhouse, L.; Feinstein, C.; Levin, H. Plasma Oxytocin Levels in Autistic Children. Biol. Psychiatry 1998, 43, 270–277. [Google Scholar] [CrossRef]
- Bakker-Huvenaars, M.J.; Greven, C.U.; Herpers, P.; Wiegers, E.; Jansen, A.; van der Steen, R.; van Herwaarden, A.E.; Baanders, A.N.; Nijhof, K.S.; Scheepers, F.; et al. Saliva oxytocin, cortisol, and testosterone levels in adolescent boys with autism spectrum disorder, oppositional defiant disorder/conduct disorder and typically developing individuals. Eur. Neuropsychopharmacol. 2020, 30, 87–101. [Google Scholar] [CrossRef]
- Husarova, V.M.; Lakatosova, S.; Pivovarciova, A.; Babinska, K.; Bakos, J.; Durdiakova, J.; Kubranska, A.; Ondrejka, I.; Ostatnikova, D. Plasma oxytocin in children with autism and its correlations with behavioral parameters in children and parents. Psychiatry Investig. 2016, 13, 174–183. [Google Scholar] [CrossRef]
- Yang, C.J.; Tan, H.P.; Yang, F.Y.; Wang, H.P.; Liu, C.L.; He, H.Z.; Sang, B.; Zhu, X.M.; Du, Y.J. The cortisol, serotonin and oxytocin are associated with repetitive behavior in autism spectrum disorder. Res. Autism. Spectr. Disord. 2015, 18, 12–20. [Google Scholar] [CrossRef]
- Zingg, H.H.; Laporte, S.A. The oxytocin receptor. Trends Endocrinol. Metab. 2003, 14, 222–227. [Google Scholar] [CrossRef]
- Fineberg, S.K.; Ross, D.A. Oxytocin and the Social Brain. Biol. Psychiatry 2017, 81, e19. [Google Scholar] [CrossRef] [Green Version]
- Marlin, B.J.; Froemke, R.C. Oxytocin modulation of neural circuits for social behavior. Dev. Neurobiol. 2017, 77, 169–189. [Google Scholar] [CrossRef] [PubMed]
- Mierop, A.; Mikolajczak, M.; Stahl, C.; Béna, J.; Luminet, O.; Lane, A.; Corneille, O. How Can Intranasal Oxytocin Research Be Trusted? A Systematic Review of the Interactive Effects of Intranasal Oxytocin on Psychosocial Outcomes. Perspect. Psychol. Sci. 2020, 15, 1228–1242. [Google Scholar] [CrossRef] [PubMed]
- Bakermans-Kranenburg, M.J.; van IJzendoorn, M.H. Sniffing around oxytocin: Review and meta-analyses of trials in healthy and clinical groups with implications for pharmacotherapy. Transl. Psychiatry 2013, 3, e258. [Google Scholar] [CrossRef]
- Yatawara, C.J.; Einfeld, S.L.; Hickie, I.B.; Davenport, T.A.; Guastella, A.J. The effect of oxytocin nasal spray on social interaction deficits observed in young children with autism: A randomized clinical crossover trial. Mol. Psychiatry 2016, 21, 1225–1231. [Google Scholar] [CrossRef] [Green Version]
- Bernaerts, S.; Boets, B.; Bosmans, G.; Steyaert, J.; Alaerts, K. Behavioral effects of multiple-dose oxytocin treatment in autism: A randomized, placebo-controlled trial with long-term follow-up. Mol Autism. 2020, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Parker, K.J.; Oztan, O.; Libove, R.A.; Sumiyoshi, R.D.; Jackson, L.P.; Karhson, D.S.; Summers, J.E.; Hinman, K.E.; Motonaga, K.S.; Phillips, J.M.; et al. Intranasal oxytocin treatment for social deficits and biomarkers of response in children with autism. Proc. Natl. Acad. Sci. USA 2017, 114, 8119–8124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guastella, A.J.; Hickie, I.B. Oxytocin Treatment, Circuitry, and Autism: A Critical Review of the Literature Placing Oxytocin into the Autism Context. Biol. Psychiatry 2016, 79, 234–242. [Google Scholar] [CrossRef]
- Alaerts, K.; Steyaert, J.; Vanaudenaerde, B.; Wenderoth, N.; Bernaerts, S. Changes in endogenous oxytocin levels after intranasal oxytocin treatment in adult men with autism: An exploratory study with long-term follow-up. Eur. Neuropsychopharmacol. 2021, 43, 147–152. [Google Scholar] [CrossRef]
- Quattrocki, E.; Friston, K. Autism, oxytocin and interoception. Neurosci. Biobehav. Rev. 2014, 47, 410–430. [Google Scholar] [CrossRef] [Green Version]
- Rutigliano, G.; Rocchetti, M.; Paloyelis, Y.; Gilleen, J.; Sardella, A.; Cappucciati, M.; Palombini, E.; Dell’Osso, L.; Caverzasi, E.; Politi, P.; et al. Peripheral oxytocin and vasopressin: Biomarkers of psychiatric disorders? A comprehensive systematic review and preliminary meta-analysis. Psychiatry Res. 2016, 241, 207–220. [Google Scholar] [CrossRef]
- Torres, N.; Martins, D.; Santos, A.J.; Prata, D.; Veríssimo, M. How do hypothalamic nonapeptides shape youth’s sociality? A systematic review on oxytocin, vasopressin and human socio-emotional development. Neurosci. Biobehav. Rev. 2018, 90, 309–331. [Google Scholar] [CrossRef] [PubMed]
- Ankit Rohatgi. WebPlotDigitizer—Copyright 2010–2017 Ankit Rohatgi. 2017. Available online: https://automeris.io/WebPlotDigitizer/ (accessed on 18 November 2021).
- Valstad, M.; Alvares, G.; Egknud, M.; Matziorinis, A.M.; Andreassen, O.A.; Westlye, L.T.; Quintana, D.S. The correlation between central and peripheral oxytocin concentrations: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2017, 78, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RevMan|Cochrane Training. Available online: https://training.cochrane.org/online-learning/core-software-cochrane-reviews/revman (accessed on 18 July 2021).
- Durlak, J.A. How to Select, Calculate, and Interpret Effect Sizes. J. Pediatr. Psychol. 2009, 34, 917–928. [Google Scholar] [CrossRef] [Green Version]
- Whiting, P.F.; Rutjes, A.W.; Westwood, M.E.; Mallett, S.; Deeks, J.J.; Reitsma, J.B.; Leeflang, M.M.; Sterne, J.A.; Bossuyt, P.M.; QUADAS-2 Group. QUADAS-2: A revised tool for the quality assessment of diagnostic accuracy studies. Ann. Intern. Med. 2011, 155, 529–536. [Google Scholar] [CrossRef]
- Green, L.A.; Fein, D.; Modahl, C.; Feinstein, C.; Waterhouse, L.; Morris, M. Oxytocin and autistic disorder: Alterations in peptide forms. Biol. Psychiatry 2001, 50, 609–613. [Google Scholar] [CrossRef]
- Mariscal, M.G.; Oztan, O.; Rose, S.M.; Libove, R.A.; Jackson, L.P.; Sumiyoshi, R.D.; Trujillo, T.H.; Carson, D.S.; Phillips, J.M.; Garner, J.P.; et al. Blood oxytocin concentration positively predicts contagious yawning behavior in children with autism spectrum disorder. Autism. Res. 2019, 12, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Komagome, A.; Iguchi-Sherry, A.; Nagasaka, A.; Yuhi, T.; Higashida, H.; Rooksby, M.; Kikuchi, M.; Arai, O.; Minami, K.; et al. Participatory Art Activities Increase Salivary Oxytocin Secretion of ASD Children. Brain Sci. 2020, 10, 680. [Google Scholar] [CrossRef]
- Taurines, R.; Schwenck, C.; Lyttwin, B.; Schecklmann, M.; Jans, T.; Reefschläger, L.; Geissler, J.; Gerlach, M.; Romanos, M. Oxytocin plasma concentrations in children and adolescents with autism spectrum disorder: Correlation with autistic symptomatology. ADHD Atten. Deficit. Hyperact. Disord. 2014, 6, 231–239. [Google Scholar] [CrossRef]
- Fujisawa, T.X.; Tanaka, S.; Saito, D.N.; Kosaka, H.; Tomoda, A. Visual attention for social information and salivary oxytocin levels in preschool children with autism spectrum disorders: An eye-tracking study. Front. Neurosci. 2014, 8, 295. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, J.D.; Ellerbeck, K.A.; Kelly, K.A.; Fleming, K.K.; Jamison, T.R.; Coffey, C.W.; Smith, C.M.; Reese, R.M.; Sands, S.A. Evidence for alterations in stimulatory G proteins and oxytocin levels in children with autism. Psychoneuroendocrinology 2014, 40, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Lakatosova, S.; Bakos, J.; Pivovarciova, A.; Husarova, V.; Durdiakova, J.; Bacova, Z.; Kubranska, A.; Babinska, K.; Ostatnikova, D.; Sunbul, M.; et al. Plasma oxytocin levels are reduced in Slovak autistic boys. Bratislava Med. J. 2015, 116, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.; Bales, K.L.; Taylor, S.L.; Yoon, J.; Hostetler, C.M.; Carter, C.S.; Solomon, M. Oxytocin and Vasopressin in Children and Adolescents with Autism Spectrum Disorders: Sex Differences and Associations with Symptoms. Autism. Res. 2013, 6, 91–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Althaus, M.; Groen, Y.; Wijers, A.A.; Noltes, H.; Tucha, O.; Sweep, F.C.; Calcagnoli, F.; Hoekstra, P.J. Do blood plasma levels of oxytocin moderate the effect of nasally administered oxytocin on social orienting in high-functioning male adults with autism spectrum disorder? Psychopharmacology 2016, 233, 2737–2751. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, T.; Fujisawa, T.X.; Inohara, K.; Okamoto, Y.; Matsumura, Y.; Tsuchiya, K.; Katayama, T.; Munesue, T.; Tomoda, A.; Wada, Y.; et al. Attenuated relationship between salivary oxytocin levels and attention to social information in adolescents and adults with autism spectrum disorder: A comparative study. Ann. Gen. Psychiatry 2020, 19, 38. [Google Scholar] [CrossRef] [PubMed]
- Procyshyn, T.L.; Lombardo, M.V.; Lai, M.C.; Auyeung, B.; Crockford, S.K.; Deakin, J.; Soubramanian, S.; Sule, A.; Baron-Cohen, S.; Bethlehem, R.A.I. Effects of oxytocin administration on salivary sex hormone levels in autistic and neurotypical women. Mol. Autism. 2020, 11, 20. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, K.; Macdonald, T. The peptide that binds: A systematic review of oxytocin and its prosocial effects in humans. Harv. Rev. Psychiatry 2010, 18, 1–21. [Google Scholar] [CrossRef]
- Freeman, S.M.; Palumbo, M.C.; Lawrence, R.H.; Smith, A.L.; Goodman, M.M.; Bales, K.L. Effect of age and autism spectrum disorder on oxytocin receptor density in the human basal forebrain and midbrain. Transl. Psychiatry 2018, 8, 257. [Google Scholar] [CrossRef]
- Moerkerke, M.; Bonte, M.-L.; Daniels, N.; Chubar, V.; Alaerts, K.; Steyaert, J.; Boets, B. Oxytocin receptor gene (OXTR) DNA methylation is associated with autism and related social traits—A systematic review. Res. Autism. Spectr. Disord. 2021, 85, 101785. [Google Scholar] [CrossRef]
- Carter, C.S. Sex differences in oxytocin and vasopressin: Implications for autism spectrum disorders? Behav. Brain Res. 2007, 176, 170–186. [Google Scholar] [CrossRef]
- Kramer, K.M.; Cushing, B.S.; Carter, C.S.; Wu, J.; Ottinger, M.A. Sex and species differences in plasma oxytocin using an enzyme immunoassay. Can. J. Zool. 2004, 82, 1194–1200. [Google Scholar] [CrossRef]
- Patisaul, H.B.; Scordalakes, E.M.; Young, L.J.; Rissman, E.F. Oxytocin, But Not Oxytocin Receptor, is Regulated by Oestrogen Receptor β in the Female Mouse Hypothalamus. J. Neuroendocrinol. 2003, 15, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Witt, D.M.; Carter, C.S.; Lnsel, T.R. Oxytocin Receptor Binding in Female Prairie Voles: Endogenous and Exogenous Oestradiol Stimulation. J. Neuroendocrinol. 1991, 3, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Leng, G.; Ludwig, M. Intranasal Oxytocin: Myths and Delusions. Biol. Psychiatry 2016, 79, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Kumsta, R.; Heinrichs, M. Oxytocin, stress and social behavior: Neurogenetics of the human oxytocin system. Curr. Opin. Neurobiol. 2013, 23, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Quintana, D.S.; Alvares, G.A.; Hickie, I.B.; Guastella, A.J. Do delivery routes of intranasally administered oxytocin account for observed effects on social cognition and behavior? A two-level model. Neurosci. Biobehav. Rev. 2015, 49, 182–192. [Google Scholar] [CrossRef]
- Shamay-Tsoory, S.G.; Abu-Akel, A. The Social Salience Hypothesis of Oxytocin. Biol. Psychiatry 2016, 79, 194–202. [Google Scholar] [CrossRef]
- Engel, S.; Laufer, S.; Miller, R.; Niemeyer, H.; Knaevelsrud, C.; Schumacher, S. Demographic, sampling- and assay-related confounders of endogenous oxytocin concentrations: A systematic review and meta-analysis. Front. Neuroendocrinol. 2019, 54, 100775. [Google Scholar] [CrossRef]
- McCullough, M.E.; Churchland, P.S.; Mendez, A.J. Problems with measuring peripheral oxytocin: Can the data on oxytocin and human behavior be trusted? Neurosci. Biobehav. Rev. 2013, 37, 1485–1492. [Google Scholar] [CrossRef] [Green Version]
- Veening, J.G.; Olivier, B. Intranasal administration of oxytocin: Behavioral and clinical effects, a review. Neurosci. Biobehav. Rev. 2013, 37, 1445–1465. [Google Scholar] [CrossRef]
- Feldman, R. Bio-behavioral Synchrony: A Model for Integrating Biological and Microsocial Behavioral Processes in the Study of Parenting. Parenting 2012, 12, 154–164. [Google Scholar] [CrossRef]
- Papasteri, C.C.; Sofonea, A.; Boldasu, R.; Poalelungi, C.; Tomescu, M.I.; Pistol, C.A.D.; Vasilescu, R.I.; Nedelcea, C.; Podina, I.R.; Berceanu, A.I.; et al. Social Feedback During Sensorimotor Synchronization Changes Salivary Oxytocin and Behavioral States. Front. Psychol. 2020, 11, 2495. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Subjects | Sample | Assay | [OT] | Unit | Results | ||
|---|---|---|---|---|---|---|---|---|
| ASD | NT | ASD | NT | |||||
| OT levels in children (13) | ||||||||
| 1. Abdulamir et al. (2016) | n = 60 (60 M) Age (y): 7.28 ± 2.89 DSM-5 | n = 26 (26 M) Age (y): 6.92 ± 2.59 | Plasma | ELISA | 44.72 ± 36.1 | 102.1 ± 34.31 | μIU/mL | Lower OT in ASD (p < 0.001) |
| 2. Alabdali et al. (2014) | n = 50 (50 M) Age (y): 7.0 ± 2.34 DSM-IV; CARS, SRS | n = 30 (30 M) Age (y): 7.2 ± 2.14 | Plasma | ELISA | 71.71 ± 18.09 | 139.22 ± 36.62 | μIU/mL | Lower OT in ASD (p = 0.001) |
| 3. Feldman et al. (2014) | n = 40 (34 M/6 F) Age (m): 63.38 ± 12.35 DSM-5; ADOS-2 | n = 40 Age (m): 53.56 ± 13.83 | Saliva | ELISA | 4.25 ± 0.66 | 6.89 ± 1.03 | pg/mL | Lower OT in ASD (p < 0.05) |
| 4. Fujisawa et al. (2014) | n = 15 (12 M/3 F) Age (m): 57.9 ± 13.6 DSM-5; DQ, PARS, SDQ | n = 58 (27 M/31 F) Age (m): 48.1 ± 22.7 | Saliva | ELISA | 39.33 ± 23.52 M: 40.3 ± 23.52 F: 35.7 ± 20.11 | 44.5 ± 24.89 M: 45.7 ± 29.78 F: 43.4 ± 20.15 | pg/mL | No significant differences between groups (p = 0.449) |
| 5. Husarova et al. (2016) | n = 19 (19 M) Age (m): 56.7 ± 25.4 ICD-10; CARS, ADI | n = 44 (44 M) Age (m): 58.9 ± 23.0 | Plasma | ELISA | 124.10 ± 90.59 | 267.77 ± 212.37 | pg/mL | Lower OT in ASD (p = 0.0004) |
| 6. Jacobson et al. (2014) | n = 37 (25 M/12 F) Age (y): 4.73 ± 0.61 DSM-IV-TR; ADI-R, ADOS | n = 41 (24 M/17 F) Age (y): 4.85 ± 0.61 | Plasma | ELISA | M: 24.41 ± 7.45 F: 23.04 ± 6.97 | M: 18,58 + 6.98 F: 22.59 + 8.82 | pg/mL | Higher OT in male ASD only (p = 0.022) |
| 7. Lakatosova et al. (2015) | n = 104 (80 M/24 F) Age (y): 7 ± 5.5 DSM-IV | n = 128 (103 M/25 F) Age (y): 10.5 ± 7 | Plasma | ELISA | M: 208.1 ± 238.63F: 282.9 ± 318.92 | M: 281.7 ± 200.85F: 340.7 ± 340.70 | pg/mL | Lower OT in male ASD only (M: p = 0.0248; F: p = 0.5058) |
| 8. Mariscal et al. (2019) | n = 34 (28 M/6 F) Age (y): 9.26 ± 0.37 DSM-IV-TR/DSM-V; ADI-R, ADOS | n = 30 (21 M/9 F) Age (y): 8.80 ± 0.40 | Plasma | ELISA | 8.62 ± 5.36 | 10.54 ± 5.37 | pg/mL | No significant differences between groups (p = 0.1564) |
| 9. Modahl et al. (1998) | n = 29 (29 M) Age (y): 8.1 + 1.7 DSM-IV | n = 30 (30 M) Age (y): 8.8 + 1.8 | Plasma | RIA | 0.64 ± 0.58 | 1.16 ± 0.77 | pg/mL | Lower OT in ASD (p < 0.004) |
| 10. Tanaka et al. (2020) | n = 12 (11 M/1 F) Age (m): 135 ± 16.7 DSM-IV-TR, DSM-V; CARS, ADOS, DISCO | n = 8 (4 M/4 F) Age (m): 107 ± 6.9 | Saliva | ELISA | 167.9 ± 62.01 | 161.5 ± 54.87 | pg/mL | No significant difference between groups |
| 11. Taurines et al. (2014) | n = 19 (19 M) Age (y): 10.7 ± 3.8 ICD-10; ADI-R, ADOS | n = 17 (17 M) Age (y): 13.6 ± 2.1 | Plasma | RIA | 19.6 ± 7.1 | 14.4 ± 9.6 | pg/mL | No significant difference between groups (p = 0.132) |
| 12. Yang et al. (2015) | n = 43 (35 M/8 F) Age (y): 7.51 ± 1.47 DSM-5; CARS | n = 40 (30 M/10 F) Age (y): 7.83 ± 1.63 | Plasma | ELISA | 116.47 ± 41.57 | 141.05 ± 51.61 | pg/mL | Lower OT in ASD (p = 0.022) |
| 13. Zhang et al. (2016) | n = 84 (71 M/13 F) Age (y): 3.95 ± 1.26 DSM-IV-TR; CARS | n = 85 (71 M/14 F) Age (y): 4.80 ± 1.22 | Plasma | ELISA | 20.05 ± 13.88 | 25.76 ± 15.30 | pg/mL | Lower OT in ASD (p = 0.028) |
| OT levels in adolescents (2) | ||||||||
| 14. Bakker-Huvenaars et al. (2020) | n = 49 (49 M) Age (y): 15.0 ± 2.1 DSM-5; DISC-IV | n = 28 (28 M) Age (y): 15.9 ± 1.8 | Saliva | RIA | −0.22 ± 0.89 | 0.49 ± 0.97 | z-score | Lower OT in ASD (p = 0.002) |
| 15. Miller et al. (2013) | n = 40 (21 M/19 F) Age (y): M: 12.24 ± 3.56; F: 11.79 ± 3.43 DSM-IV-TR; ADOS | n = 35 (19 M/16 F) Age (y): M: 11.74 ± 2.49; F: 12.94 ± 3.19 | Plasma | ELISA | M: 357.12 ± 126.05F: 525.23 ± 325.75 | M: 361.52 ± 315.26F: 434.33 ± 332.27 | pg/mL | No significant differences between groups (p = 0.270) |
| OT levels in adults (3) | ||||||||
| 16. Althaus et al. (2016) | n = 31 (31 M) Age (y): 22.67 ± 4.22 DSM-IV-TR; ADOS, | n = 30 (30 M) Age (y): 22.67 ± 4.22 | Plasma | RIA | 1.34 ± 1.05 | 0.67 ± 0.77 | pmol/L | Higher OT in ASD (p = 0.006) |
| 17. Fujioka et al. (2020) | n = 17 (17 M) Age (y): 27.4 ± 7.2 DSM-IV; DISCO | n = 24 (24 M) Age (y): 29.0 ± 9.8 | Saliva | ELISA | 36.2 ± 13.2 | 43.6 ± 17.0 | pg/mL | No significant difference between groups (p = 0.154) |
| 18. Procyshyn et al. (2020) | n = 16 (16 F) Age (y): 29.9 ± 8.4 DSM-IV | n = 29 (29 F) Age (y): 27.2 ± 8.1 | Saliva | ELISA | 3.1 ± 0.5 | 2.8 ± 0.6 | pg/mL | No significant difference between groups (p = 0.064) |
| Study | Risk of Bias | Applicability Concerns | |||||
|---|---|---|---|---|---|---|---|
| Patient Selection | Index Test | Reference Test | Flow and Timing | Patient Selection | Index Test | Reference Test | |
| 1. Abdulamir et al. (2016) [4] | ? | ? | ☹ | ☺ | ☺ | ☺ | ☺ |
| 2. Alabdali et al. (2014) [5] | ☹ | ? | ☺ | ☹ | ☺ | ☺ | ☺ |
| 3. Feldman et al. (2014) [6] | ? | ? | ☺ | ☹ | ☺ | ☺ | ☺ |
| 4. Fujisawa et al., 2014) [33] | ? | ? | ☹ | ? | ☺ | ☺ | ☺ |
| 5. Husarova et al. (2016) [9] | ? | ? | ☺ | ? | ☺ | ☺ | ☺ |
| 6. Jacobson et al. (2014) [34] | ☹ | ? | ☺ | ? | ☺ | ☺ | ☺ |
| 7. Lakatosova et al. (2015) [35] | ? | ? | ☹ | ☹ | ☺ | ☺ | ☺ |
| 8. Mariscal et al. (2019) [30] | ☹ | ☺ | ☺ | ? | ☺ | ☺ | ☺ |
| 9. Modahl et al. (1998) [7] | ? | ? | ☹ | ? | ☺ | ☺ | ☺ |
| 10. Tanaka et al. (2020) [31] | ☹ | ? | ☺ | ☹ | ☺ | ☺ | ☺ |
| 11. Taurines et al., 2014) [32] | ? | ? | ☺ | ? | ? | ☺ | ☺ |
| 12. Yang et al. (2015) [10] | ☹ | ? | ☺ | ☹ | ☺ | ☺ | ☺ |
| 13. Zhang et al. (2016) [3] | ? | ? | ? | ? | ☺ | ☺ | ? |
| 14. Bakker-Huvenaars et al. (2016) [8] | ☹ | ? | ☺ | ☺ | ☺ | ☺ | ☺ |
| 15. Miller et al. (2013) [36] | ☹ | ? | ☺ | ? | ☺ | ☺ | ☺ |
| 16. Althaus et al. (2016) [37] | ? | ? | ☺ | ? | ☺ | ☺ | ☺ |
| 17. Fujioka et al. (2020) [38] | ☹ | ? | ☺ | ? | ☺ | ☺ | ☺ |
| 18. Procyshyn et al. (2020) [39] | ? | ? | ☹ | ? | ☺ | ☺ | ? |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moerkerke, M.; Peeters, M.; de Vries, L.; Daniels, N.; Steyaert, J.; Alaerts, K.; Boets, B. Endogenous Oxytocin Levels in Autism—A Meta-Analysis. Brain Sci. 2021, 11, 1545. https://doi.org/10.3390/brainsci11111545
Moerkerke M, Peeters M, de Vries L, Daniels N, Steyaert J, Alaerts K, Boets B. Endogenous Oxytocin Levels in Autism—A Meta-Analysis. Brain Sciences. 2021; 11(11):1545. https://doi.org/10.3390/brainsci11111545
Chicago/Turabian StyleMoerkerke, Matthijs, Mathieu Peeters, Lyssa de Vries, Nicky Daniels, Jean Steyaert, Kaat Alaerts, and Bart Boets. 2021. "Endogenous Oxytocin Levels in Autism—A Meta-Analysis" Brain Sciences 11, no. 11: 1545. https://doi.org/10.3390/brainsci11111545
APA StyleMoerkerke, M., Peeters, M., de Vries, L., Daniels, N., Steyaert, J., Alaerts, K., & Boets, B. (2021). Endogenous Oxytocin Levels in Autism—A Meta-Analysis. Brain Sciences, 11(11), 1545. https://doi.org/10.3390/brainsci11111545