
An Exploratory Study on Resting-State Functional Connectivity in Individuals with Disorganized Attachment: Evidence for Key Regions in Amygdala and Hippocampus

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Image Acquisition and Analysis
3. Results
4. Discussion
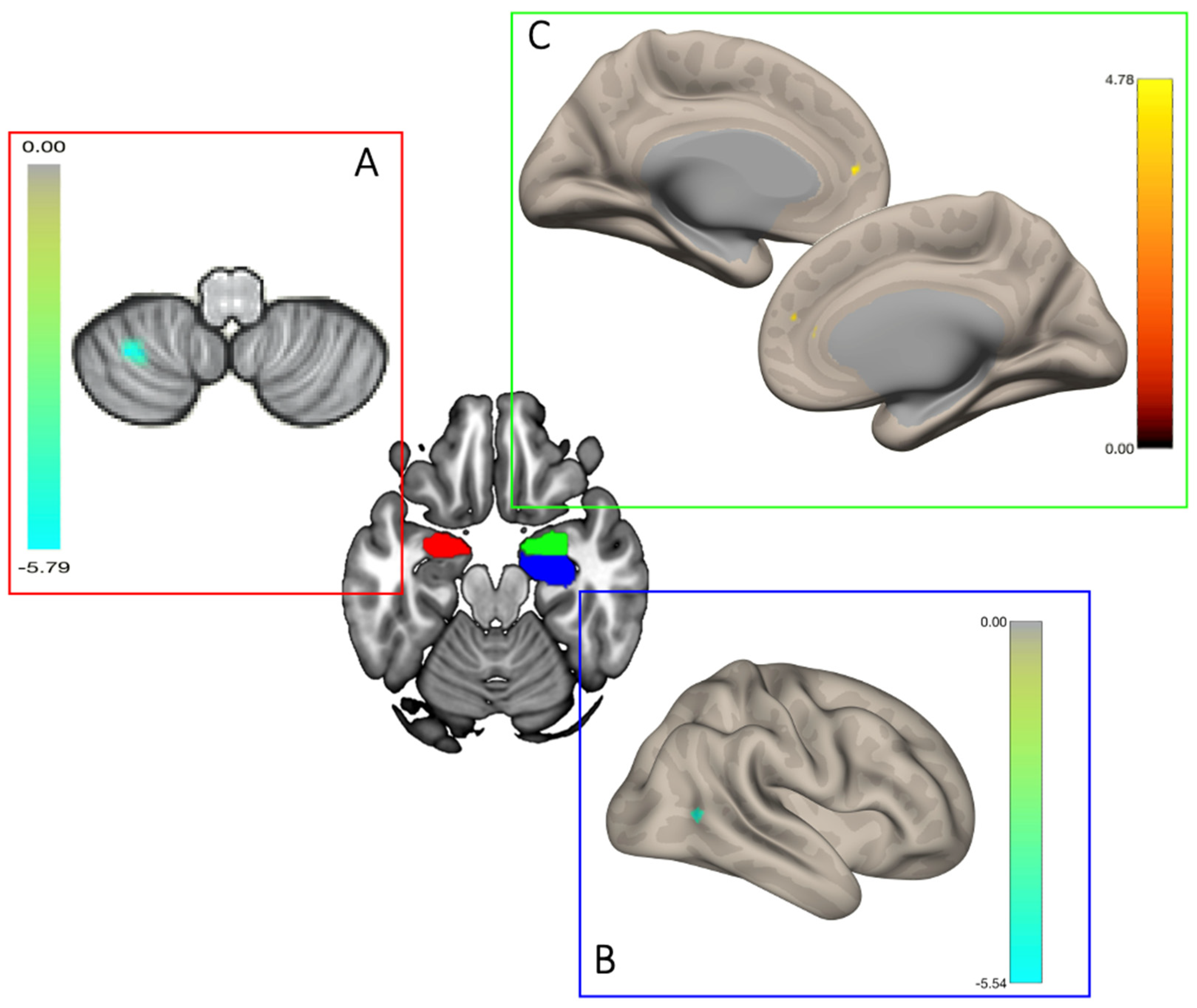
4.1. The Role of the Amygdala: Altered Functional Connectivity of the Bilateral Amygdala in UD Compared to O Participants
4.2. Higher Functional Connectivity between the Right Amygdala and the ACC in UD
4.3. Lower Functional Connectivity between the Left Amygdala and the Left Cerebellum in UD
4.4. Lower Functional Connectivity between the Right Hippocampus and the Right Middle Temporal Gyrus in UD
5. Limitations and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bowlby, J. Attachment and Loss; Basic Books Classics: New York, NY, USA, 1982; Volume 1. [Google Scholar]
- Bretherton, I.; Munholland, K.A. The internal working model construct in light of contemporary neuroimaging research. In Handbook of Attachment: Theory, Research, and Clinical Applications, 3rd ed.; Cassidy, J., Shaver, P.R., Eds.; Guilford Publications: New York, NY, USA, 2016; pp. 63–88. [Google Scholar]
- Cassidy, J. Emotion regulation: Influences of attachment relationships. Monogr. Soc. Res. Child 1994, 59, 228–249. [Google Scholar] [CrossRef]
- Main, M.; Kaplan, N.; Cassidy, J. Security in infancy, childhood, and adulthood: A move to the level of representation. Monogr. Soc. Res. Child 1985, 50, 66–104. [Google Scholar] [CrossRef]
- Diamond, L.M.; Hicks, A.M. Attachment style, current relationship security, and negative emotions: The mediating role of physiological regulation. J. Soc. Pers. Relat. 2005, 22, 499–518. [Google Scholar] [CrossRef]
- Fox, N.A. The Development of Emotion Regulation: Biological and Behavioral Considerations; University of Chicago Press: Chicago, IL, USA, 1994. [Google Scholar]
- Bakermans-Kranenburg, M.J.; van IJzendoorn, M.H. The first 10,000 Adult Attachment Interviews: Distributions of adult attachment representations in clinical and non-clinical groups. Attach. Hum. Dev. 2009, 11, 223–263. [Google Scholar] [CrossRef] [PubMed]
- Liotti, G. Trauma, dissociation, and disorganized attachment: Three strands of a single braid. Psychother. Theor. Res. 2004, 41, 472–486. [Google Scholar] [CrossRef] [Green Version]
- Lyons-Ruth, K.; Jacobvitz, D. Attachment disorganization: Unresolved loss, relational violence, and lapses in behavioral and attentional strategies. In Handbook of Attachment: Theory, Research, and Clinical Applications; Cassidy, J., Shaver, P.R., Eds.; Guilford Press: New York, NY, USA, 1999; pp. 520–554. [Google Scholar]
- Main, M.; Goldwyn, R.; Hesse, E. Adult Attachment Scoring and Classification Systems; Version 7.1. Unpublished manuscript; University of California: Berkeley, CA, USA, 2002. [Google Scholar]
- Hesse, E. The Adult Attachment Interview: Protocol, method of analysis, and empirical studies. In Handbook of Attachment: Theory, Research, and Clinical Applications, 2nd ed.; Cassidy, J., Shaver, P.R., Eds.; Guilford Press: New York, NY, USA, 2008; pp. 552–598. [Google Scholar]
- Lyons-Ruth, K.; Jacobvitz, D. Attachment disorganization from infancy to adulthood: Neurobiological correlates, parenting contexts, and pathways to disorder. In Handbook of Attachment: Theory, Research, and Clinical Applications, 3rd ed.; Cassidy, J., Shaver, P.R., Eds.; Guilford Press: New York, NY, USA, 2016; pp. 667–695. [Google Scholar]
- Mikulincer, M.; Shaver, P.R. The attachment behavioral system in adulthood: Activation, psychodynamics, and interpersonal processes. In Advances in Experimental Social Psychology; Zanna, M.P., Ed.; Academic Press: San Diego, CA, USA, 2003; pp. 53–152. [Google Scholar]
- Meares, R. The Metaphor of Play. Origin and Breakdown of Personal Being, 3rd ed.; Routledge: London, UK, 2005. [Google Scholar]
- Liotti, G. Disorganized/disoriented attachment in the etiology of the dissociative disorders. Dissociation 1992, 5, 196–204. [Google Scholar]
- Carlson, E.A.; Egeland, B.; Sroufe, L.A. A prospective investigation of the development of borderline personality symptoms. Dev. Psychopathol. 2009, 21, 1311–1334. [Google Scholar] [CrossRef]
- Lyons-Ruth, K.; Bureau, J.-F.; Holmes, B.; Easterbrooks, A.; Brooks, N. Borderline symptoms and suicidality/self-injury in late adolescence: Prospectively observed relationship correlates in infancy and childhood. Psychiatry Res. 2013, 206, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Lyons-Ruth, K.; Pechtel, P.; Yoon, S.A.; Anderson, C.M.; Teicher, M.H. Disorganized attachment in infancy predicts greater amygdala volume in adulthood. Behav. Brain Res. 2016, 308, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Pechtel, P.; Lyons-Ruth, K.; Anderson, C.M.; Teicher, M.H. Sensitive periods of amygdala development: The role of maltreatment in preadolescence. Neuroimage 2014, 97, 236–244. [Google Scholar] [CrossRef] [Green Version]
- Buchheim, A.; Erk, S.; George, C.; Kächele, H.; Ruchsow, M.; Spitzer, M.; Kircher, T.; Walter, H. Measuring attachment representation in an fMRI environment: A pilot study. Psychopathology 2006, 39, 144–152. [Google Scholar] [CrossRef]
- Buchheim, A.; Erk, S.; George, C.; Kächele, H.; Martius, P.; Pokorny, D.; Spitzer, M.; Walter, H. Neural response during the activation of the attachment system in patients with borderline personality disorder: An fMRI study. Front. Hum. Neurosci. 2016, 10, 389. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Fonagy, P.; Allen, J.; Strathearn, L. Mothers’ unresolved trauma blunts amygdala response to infant distress. Soc. Neurosci. 2014, 9, 352–363. [Google Scholar] [CrossRef] [Green Version]
- Van Hoof, M.J.; Riem, M.M.; Garrett, A.S.; van der Wee, N.J.; van IJzendoorn, M.H.; Vermeiren, R.R.M. Unresolved–disorganized attachment adjusted for a general psychopathology factor associated with atypical amygdala resting-state functional connectivity. Eur. J. Psychotraumatol. 2019, 10, 1583525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hoof, M.J.; Riem, M.; Garrett, A.; Pannekoek, N.; van der Wee, N.; van IJzendoorn, M.; Vermeiren, R. Unresolved–disorganized attachment is associated with smaller hippocampus and increased functional connectivity beyond psychopathology. J. Trauma Stress 2019, 32, 742–752. [Google Scholar] [CrossRef] [Green Version]
- Rifkin-Graboi, A.; Tan, H.M.; Shaun, G.K.Y.; Sim, L.W.; Sanmugam, S.; Chong, Y.S.; Tan, K.H.; Shek, L.; Gluckman, P.D.; Chen, H.; et al. An initial investigation of neonatal neuroanatomy, caregiving, and levels of disorganized behavior. P. Natl. Acad. Sci. USA 2019, 116, 16787–16792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortes Hidalgo, A.P.; Muetzel, R.; Luijk, M.P.; Bakermans-Kranenburg, M.J.; El Marroun, H.; Vernooij, M.W.; van IJzendoorn, M.H.; White, T.; Tiemeier, H. Observed infant-parent attachment and brain morphology in middle childhood–A population-based study. Dev. Cogn. Neurosci. 2019, 40, 100724. [Google Scholar] [CrossRef] [PubMed]
- Khoury, J.E.; Zona, K.; Bertha, E.; Choi-Kain, L.; Hennighausen, K.; Lyons-Ruth, K. Disorganized attachment interactions among young adults with borderline personality disorder, other diagnoses, and no diagnosis. J. Pers. Disord. 2020, 34, 764–784. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Battaglia, S.; Sciamanna, G.; Tortora, F.; Laricchiuta, D. Memories are not written in stone: Re-writing fear memories by means of non-invasive brain stimulation and optogenetic manipulations. Neurosci. Biobehav. Rev. 2021, 127, 334–352. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Battaglia, S.; Garofalo, S.; Tortora, F.; Avenanti, A.; di Pellegrino, G. State-dependent TMS over prefrontal cortex disrupts fear-memory reconsolidation and prevents the return of fear. Curr. Biol. 2020, 30, 3672–3679. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Garofalo, S.; di Pellegrino, G.; Starita, F. Revaluing the role of vmPFC in the acquisition of Pavlovian threat conditioning in humans. J. Neurosci. 2020, 40, 8491–8500. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Battaglia, S.; Avenanti, A.; di Pellegrino, G. Don’t Hurt Me No More: State-dependent Transcranial Magnetic Stimulation for the treatment of specific phobia. J. Affect. Disord. 2021, 286, 78–79. [Google Scholar] [CrossRef] [PubMed]
- Spitoni, G.F.; Zingaretti, P.; Giovanardi, G.; Antonucci, G.; Galati, G.; Lingiardi, V.; Cruciani, G.; Titone, G.; Boccia, M. Disorganized Attachment pattern affects the perception of Affective Touch. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- George, C.; Kaplan, N.; Main, M. Adult Attachment Interview. Unpublished Manuscript, 3rd ed.; Department of Psychology, University of California: Berkley, CA, USA, 1996. [Google Scholar]
- Krueger, R.F.; Derringer, J.; Markon, K.E.; Watson, D.; Skodol, A.E. Initial construction of a maladaptive personality trait model and inventory for DSM-5. Psychol. Med. 2012, 42, 1879–1890. [Google Scholar] [CrossRef] [Green Version]
- Derogatis, L.R. The Symptom Checklist 90-R: Administration, Scoring and Procedures Manual, 3rd ed.; National Computing Systems: Minneapolis, MN, USA, 1994. [Google Scholar]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. A functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage 2012, 59, 2142–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desikan, R.S.; Ségonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage 2006, 31, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Chumbley, J.; Worsley, K.; Flandin, G.; Friston, K. Topological FDR for neuroimaging. Neuroimage 2010, 49, 3057–3064. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, R.P.; Biggs, A.; Chen, G.; Pine, D.S.; Grillon, C. Contextual fear conditioning in humans: Cortical-hippocampal and amygdala contributions. J. Neurosci. 2008, 28, 6211–6219. [Google Scholar] [CrossRef]
- Büchel, C.; Dolan, R.J.; Armony, J.L.; Friston, K.J. Amygdala–hippocampal involvement in human aversive trace conditioning revealed through event-related functional magnetic resonance imaging. J. Neurosci. 1999, 19, 10869–10876. [Google Scholar] [CrossRef] [PubMed]
- Gorman, J.M.; Kent, J.M.; Sullivan, G.M.; Coplan, J.D. Neuroanatomical hypothesis of panic disorder, revised. Am. J. Psychiatry 2000, 157, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.D.; Reiman, E.M.; Bradley, M.M.; Lang, P.J.; Ahern, G.L.; Davidson, R.J.; Schwartz, G.E. Neuroanatomical correlates of pleasant and unpleasant emotion. Neuropsychologia 1997, 35, 1437–1444. [Google Scholar] [CrossRef]
- Morrison, S.E.; Salzman, C.D. Re-valuing the amygdala. Curr. Opin. Neurobiol. 2010, 20, 221–230. [Google Scholar] [CrossRef]
- Sehlmeyer, C.; Schöning, S.; Zwitserlood, P.; Pfleiderer, B.; Kircher, T.; Arolt, V.; Konrad, C. Human fear conditioning and extinction in neuroimaging: A systematic review. PLoS ONE 2009, 4, e5865. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Serio, G.; Scarpazza, C.; D’Ausilio, A.; Borgomaneri, S. Frozen in (e) motion: How reactive motor inhibition is influenced by the emotional content of stimuli in healthy and psychiatric populations. Behav. Res. Ther. 2021, 146, 103963. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Serio, G.; Battaglia, S. Please, don’t do it! Ten years of progress of non-invasive brain stimulation in action inhibition. Cortex 2020, 132, 404–422. [Google Scholar] [CrossRef]
- Jedd, K.; Hunt, R.H.; Cicchetti, D.; Hunt, E.; Cowell, R.; Rogosch, F.; Toth, S.L.; Thomas, K.M. Long-term consequences of childhood maltreatment: Altered amygdala functional connectivity. Dev. Psychopathol. 2015, 27, 1577–1589. [Google Scholar] [CrossRef] [Green Version]
- Dean, A.C.; Kohno, M.; Hellemann, G.; London, E.D. Childhood maltreatment and amygdala connectivity in methamphetamine dependence: A pilot study. Brain Behav. 2014, 4, 867–876. [Google Scholar] [CrossRef]
- Lupien, S.J.; Parent, S.; Evans, A.C.; Tremblay, R.E.; Zelazo, P.D.; Corbo, V.; Pruessner, J.C.; Séguin, J.R. Larger amygdala but no change in hippocampal volume in 10-year-old children exposed to maternal depressive symptomatology since birth. Proc. Natl. Acad. Sci. USA 2011, 108, 14324–14329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schore, A.N. Attachment trauma and the developing right brain: Origins of pathological dissociation. In Dissociation and the Dissociative Disorders: DSM-V and Beyond; Dell, P.F., O’Neil, J.A., Eds.; Routledge/Taylor & Francis Group: London, UK, 2009; pp. 107–141. [Google Scholar]
- Caldji, C.; Tannenbaum, B.; Sharma, S.; Francis, D.; Plotsky, P.M.; Meaney, M.J. Maternal care during infancy regulates the development of neural systems mediating the expression of fearfulness in the rat. Proc. Natl. Acad. Sci. USA 1998, 95, 5335–5340. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.M.; Jing, D.; Yang, R.R.; Tottenham, N.; Lee, F.S.; Casey, B.J. Early-life stress has persistent effects on amygdala function and development in mice and humans. Proc. Natl. Acad. Sci. USA 2013, 110, 18274–18278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raineki, C.; Moriceau, S.; Sullivan, R.M. Developing a neurobehavioral animal model of infant attachment to an abusive caregiver. Biol. Psychiatry 2010, 67, 1137–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, C.; Machado, C.J.; Bliwise, N.G.; Bachevalier, J. Maturation of the hippocampal formation and amygdala in Macaca mulatta: A volumetric magnetic resonance imaging study. Hippocampus 2010, 20, 922–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uematsu, A.; Matsui, M.; Tanaka, C.; Takahashi, T.; Noguchi, K.; Suzuki, M.; Nishijo, H. Developmental trajectories of amygdala and hippocampus from infancy to early adulthood in healthy individuals. PLoS ONE 2012, 7, e46970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Werff, S.J.; Pannekoek, J.N.; Veer, I.M.; van Tol, M.J.; Aleman, A.; Veltman, D.J.; Zitman, F.G.; Rombouts, S.A.R.B.; Elzinga, B.M.; van der Wee, N.J.A. Resting-state functional connectivity in adults with childhood emotional maltreatment. Psychol. Med. 2013, 43, 1825–1836. [Google Scholar] [CrossRef]
- Wang, L.; Dai, Z.; Peng, H.; Tan, L.; Ding, Y.; He, Z.; Zhang, Y.; Xia, M.; Li, Z.; Li, W.; et al. Overlapping and segregated resting-state functional connectivity in patients with major depressive disorder with and without childhood neglect. Hum. Brain Mapp. 2014, 35, 1154–1166. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.M.; White, D.; Hadley, J.; Hutcheson, N.; Shelton, R.; Sreenivasan, K.; Deshpande, G. Early life trauma and directional brain connectivity within major depression. Hum. Brain Mapp. 2014, 35, 4815–4826. [Google Scholar] [CrossRef]
- Strawn, J.R.; Bitter, S.M.; Weber, W.A.; Chu, W.J.; Whitsel, R.M.; Adler, C.; Cerullo, M.A.; Eliassen, J.; Strakowski, S.M.; Del Bello, M.P. Neurocircuitry of generalized anxiety disorder in adolescents: A pilot functional neuroimaging and functional connectivity study. Depress. Anxiety 2012, 29, 939–947. [Google Scholar] [CrossRef]
- Rutherford, H.J.; Lindell, A.K. Thriving and surviving: Approach and avoidance motivation and lateralization. Emot. Rev. 2011, 3, 333–343. [Google Scholar] [CrossRef]
- Tottenham, N.; Shapiro, M.; Telzer, E.H.; Humphreys, K.L. Amygdala response to mother. Dev. Sci. 2012, 15, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Dannlowski, U.; Kugel, H.; Huber, F.; Stuhrmann, A.; Redlich, R.; Grotegerd, D.; Dohm, K.; Sehlmeyer, C.; Konrad, C.; Baune, B.T.; et al. Childhood maltreatment is associated with an automatic negative emotion processing bias in the amygdala. Hum. Brain Mapp. 2013, 34, 2899–2909. [Google Scholar] [CrossRef] [PubMed]
- Herringa, R.J.; Birn, R.M.; Ruttle, P.L.; Burghy, C.A.; Stodola, D.E.; Davidson, R.J.; Essex, M.J. Childhood maltreatment is associated with altered fear circuitry and increased internalizing symptoms by late adolescence. Proc. Natl. Acad. Sci. USA 2013, 110, 19119–19124. [Google Scholar] [CrossRef] [Green Version]
- Fonagy, P.; Target, M.; Gergely, G. Attachment and borderline personality disorder: A theory and some evidence. Psychiatry Clin. 2000, 23, 103–122. [Google Scholar] [CrossRef]
- Lecompte, V.; Moss, E.; Cyr, C.; Pascuzzo, K. Preschool attachment, self-esteem and the development of preadolescent anxiety and depressive symptoms. Attach. Hum. Dev. 2014, 16, 242–260. [Google Scholar] [CrossRef]
- Mosquera, D.; Gonzalez, A.; Leeds, A.M. Early experience, structural dissociation, and emotional dysregulation in borderline personality disorder: The role of insecure and disorganized attachment. Bord. Personal. Disord. Emot. Dysregul. 2014, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Beesdo, K.; Lau, J.Y.; Guyer, A.E.; McClure-Tone, E.B.; Monk, C.S.; Nelson, E.E.; Fromm, S.J.; Goldwin, M.A.; Wittchen, H.; Leibenluft, E.; et al. Common and distinct amygdala-function perturbations in depressed vs anxious adolescents. Arch. Gen. Psychiatry 2009, 66, 275–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donegan, N.H.; Sanislow, C.A.; Blumberg, H.P.; Fulbright, R.K.; Lacadie, C.; Skudlarski, P.; Gore, J.C.; Olson, I.R.; McGlashan, T.H.; Wexler, B.E. Amygdala hyperreactivity in borderline personality disorder: Implications for emotional dysregulation. Biol. Psychiatry 2003, 54, 1284–1293. [Google Scholar] [CrossRef] [Green Version]
- MacMillan, S.; Szeszko, P.R.; Moore, G.J.; Madden, R.; Lorch, E.; Ivey, J.; Banerjee, S.P.; Rosenberg, D.R. Increased amygdala: Hippocampal volume ratios associated with severity of anxiety in pediatric major depression. J. Child Adolesc. Psychopharmacol. 2003, 13, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Okita, K.; Ghahremani, D.G.; Payer, D.E.; Robertson, C.L.; Dean, A.C.; Mandelkern, M.A.; London, E.D. Emotion dysregulation and amygdala dopamine D2-type receptor availability in methamphetamine users. Drug Alcohol Depend. 2016, 161, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Schreiner, M.W.; Klimes-Dougan, B.; Mueller, B.A.; Eberly, L.E.; Reigstad, K.M.; Carstedt, P.A.; Thomas, K.M.; Hunt, R.H.; Lim, K.O.; Cullen, K.R. Multi-modal neuroimaging of adolescents with non-suicidal self-injury: Amygdala functional connectivity. J. Affect. Disord. 2017, 221, 47–55. [Google Scholar] [CrossRef]
- Weniger, G.; Lange, C.; Sachsse, U.; Irle, E. Amygdala and hippocampal volumes and cognition in adult survivors of childhood abuse with dissociative disorders. Acta Psychiatry Scand. 2008, 118, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Botvinick, M.; Nystrom, L.E.; Fissell, K.; Carter, C.S.; Cohen, J.D. Conflict monitoring versus selection-for-action in anterior cingulate cortex. Nature 1999, 402, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; Braver, T.S.; Barch, D.M.; Botvinick, M.M.; Noll, D.; Cohen, J.D. Anterior cingulate cortex, error detection, and the online monitoring of performance. Science 1998, 280, 747–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Hof, P.R.; Guise, K.G.; Fossella, J.A.; Posner, M.I. The functional integration of the anterior cingulate cortex during conflict processing. Cereb. Cortex 2008, 18, 796–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nee, D.E.; Kastner, S.; Brown, J.W. Functional heterogeneity of conflict, error, task-switching, and unexpectedness effects within medial prefrontal cortex. Neuroimage 2011, 54, 528–540. [Google Scholar] [CrossRef] [Green Version]
- Ridderinkhof, K.R.; Van Den Wildenberg, W.P.; Segalowitz, S.J.; Carter, C.S. Neurocognitive mechanisms of cognitive control: The role of prefrontal cortex in action selection, response inhibition, performance monitoring, and reward-based learning. Brain Cogn. 2004, 56, 129–140. [Google Scholar] [CrossRef]
- Garofalo, S.; Timmermann, C.; Battaglia, S.; Maier, M.E.; di Pellegrino, G. Mediofrontal negativity signals unexpected timing of salient outcomes. J. Cogn. Neurosci. 2017, 29, 718–727. [Google Scholar] [CrossRef]
- Alexander, W.H.; Brown, J.W. The role of the anterior cingulate cortex in prediction error and signaling surprise. Top. Cogn. Sci. 2019, 11, 119–135. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, S.T.; Price, J.L. Limbic connections of the orbital and medial prefrontal cortex in macaque monkeys. J. Comp. Neurol. 1995, 363, 615–641. [Google Scholar] [CrossRef]
- Van Hoesen, G.W.; Morecraft, R.J.; Vogt, B.A. Connections of the monkey cingulate cortex. In Neurobiology of Cingulate Cortex and Limbic Thalamus; Vogt, B.A., Gabriel, M., Eds.; Birkhäuser: Boston, MA, USA, 1993; pp. 249–284. [Google Scholar]
- Etkin, A.; Egner, T.; Kalisch, R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends Cogn. Sci. 2011, 15, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Etkin, A.; Wager, T.D. Functional neuroimaging of anxiety: A meta-analysis of emotional processing in PTSD, social anxiety disorder, and specific phobia. Am. J. Psychiatry 2007, 164, 1476–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, K.L.; Fitzgerald, D.A.; Nathan, P.J.; Tancer, M.E. Association between amygdala hyperactivity to harsh faces and severity of social anxiety in generalized social phobia. Biol. Psychiatry 2006, 59, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Boccia, M.; D’Amico, S.; Bianchini, F.; Marano, A.; Giannini, A.M.; Piccardi, L. Different neural modifications underpin PTSD after different traumatic events: An fMRI meta-analytic study. Brain Imaging Behav. 2016, 10, 226–237. [Google Scholar] [CrossRef]
- Northoff, G.; Bermpohl, F. Cortical midline structures and the self. Trends Cogn. Sci. 2004, 8, 102–107. [Google Scholar] [CrossRef]
- Northoff, G.; Heinzel, A.; De Greck, M.; Bermpohl, F.; Dobrowolny, H.; Panksepp, J. Self-referential processing in our brain—A meta-analysis of imaging studies on the self. Neuroimage 2006, 31, 440–457. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G.; Qin, P.; Nakao, T. Rest-stimulus interaction in the brain: A review. Trends Neurosci. 2010, 33, 277–284. [Google Scholar] [CrossRef]
- Scalabrini, A.; Mucci, C.; Northoff, G. Is our self related to personality? A neuropsychodynamic model. Front. Hum. Neurosci. 2018, 12, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, P.; Northoff, G. How is our self related to midline regions and the default-mode network? Neuroimage 2011, 57, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G. Is the self a higher-order or fundamental function of the brain? The “basis model of self-specificity” and its encoding by the brain’s spontaneous activity. Cogn. Neurosci. 2016, 7, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; MacDonald III, A.W.; Ross, L.L.; Stenger, V.A. Anterior cingulate cortex activity and impaired self-monitoring of performance in patients with schizophrenia: An event-related fMRI study. Am. J. Psychiatry 2001, 158, 1423–1428. [Google Scholar] [CrossRef]
- Bush, G.; Luu, P.; Posner, M.I. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn. Sci. 2000, 4, 215–222. [Google Scholar] [CrossRef]
- Blizard, R.A. Disorganized attachment, development of dissociated self states, and a relational approach to treatment. J. Trauma Dissociatio. 2003, 4, 27–50. [Google Scholar] [CrossRef]
- Heath, R.G.; Harper, J.W. Ascending projections of the cerebellar fastigial nucleus to the hippocampus, amygdala, and other temporal lobe sites: Evoked potential and histological studies in monkeys and cats. Exp. Neurol. 1974, 45, 268–287. [Google Scholar] [CrossRef]
- Sang, L.; Qin, W.; Liu, Y.; Han, W.; Zhang, Y.; Jiang, T.; Yu, C. Resting-state functional connectivity of the vermal and hemispheric subregions of the cerebellum with both the cerebral cortical networks and subcortical structures. Neuroimage 2012, 61, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D. The role of the cerebellum in cognition and emotion: Personal reflections since 1982 on the dysmetria of thought hypothesis, and its historical evolution from theory to therapy. Neuropsychol. Rev. 2010, 20, 236–260. [Google Scholar] [CrossRef] [PubMed]
- Dalenberg, J.R.; Hoogeveen, H.R.; Renken, R.J.; Langers, D.R.; ter Horst, G.J. Functional specialization of the male insula during taste perception. Neuroimage 2015, 119, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Dimitrova, A.; Kolb, F.P.; Elles, H.G.; Maschke, M.; Forsting, M.; Diener, H.C.; Timmann, D. Cerebellar responses evoked by nociceptive leg withdrawal reflex as revealed by event-related FMRI. J. Neurophysiol. 2003, 90, 1877–1886. [Google Scholar] [CrossRef] [Green Version]
- Dube, A.A.; Duquette, M.; Roy, M.; Lepore, F.; Duncan, G.; Rainville, P. Brain activity associated with the electrodermal reactivity to acute heat pain. Neuroimage 2009, 45, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Petrowski, K.; Wintermann, G.B.; Hübner, T.; Smolka, M.N.; Donix, M. Neural Responses to Faces of Attachment Figures and Unfamiliar Faces: Associations with Organized and Disorganized Attachment Representations. J. Nerv. Ment. Dis. 2019, 207, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Habas, C.; Manto, M. Probing the neuroanatomy of the cerebellum using tractography. In Handbook of Clinical Neurology; Manto, M., Huisman, T.A.G.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 235–249. [Google Scholar] [CrossRef]
- Schmahmann, J.D.; Sherman, J.C. The cerebellar cognitive affective syndrome. Brain 1998, 121, 561–579. [Google Scholar] [CrossRef]
- Hanson, J.L.; Chandra, A.; Wolfe, B.L.; Pollak, S.D. Association between income and the hippocampus. PLoS ONE 2011, 6, e18712. [Google Scholar] [CrossRef] [Green Version]
- Hanson, J.L.; Nacewicz, B.M.; Sutterer, M.J.; Cayo, A.A.; Schaefer, S.M.; Rudolph, K.D.; Shirtcliff, E.A.; Pollak, S.D.; Davidson, R.J. Behavioral problems after early life stress: Contributions of the hippocampus and amygdala. Biol. Psychiatry 2015, 77, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Noble, K.G.; Houston, S.M.; Kan, E.; Sowell, E.R. Neural correlates of socioeconomic status in the developing human brain. Dev. Sci. 2012, 15, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Rao, U.; Chen, L.A.; Bidesi, A.S.; Shad, M.U.; Thomas, M.A.; Hammen, C.L. Hippocampal changes associated with early-life adversity and vulnerability to depression. Biol. Psychiatry 2010, 67, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, K.; Lind, T.; Dozier, M. Neurobiological consequences of neglect and abuse. In Handbook of Child Maltreatment: Contemporary Issues in Research and Policy; Korbin, J.E., Krugman, K.D., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 205–223. [Google Scholar]
- Daskalakis, N.P.; De Kloet, E.R.; Yehuda, R.; Malaspina, D.; Kranz, T.M. Early life stress effects on glucocorticoid—BDNF interplay in the hippocampus. Front. Mol. Neurosci. 2015, 8, 68. [Google Scholar] [CrossRef] [Green Version]
- Von Werne Baes, C.; de Carvalho Tofoli, S.M.; Martins, C.M.S.; Juruena, M.F. Assessment of the hypothalamic–pituitary–adrenal axis activity: Glucocorticoid receptor and mineralocorticoid receptor function in depression with early life stress–A systematic review. Acta Neuropsychiatr. 2012, 24, 4–15. [Google Scholar] [CrossRef]
- Cecchini, M.; Aceto, P.; Altavilla, D.; Palumbo, L.; Lai, C. The role of the eyes in processing an intact face and its scrambled image: A dense array ERP and low-resolution electromagnetic tomography (sLORETA) study. Soc. Neurosci. 2013, 8, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.; Altavilla, D.; Ronconi, A.; Aceto, P. Fear of missing out (FOMO) is associated with activation of the right middle temporal gyrus during inclusion social cue. Comput. Hum. Behav. 2016, 61, 516–521. [Google Scholar] [CrossRef]
- Ren, J.; Huang, F.; Zhou, Y.; Zhuang, L.; Xu, J.; Gao, C.; Qin, S.; Luo, J. The function of the hippocampus and middle temporal gyrus in forming new associations and concepts during the processing of novelty and usefulness features in creative designs. Neuroimage 2020, 214, 116751. [Google Scholar] [CrossRef] [PubMed]
- Pears, K.C.; Fisher, P.A. Emotion understanding and theory of mind among maltreated children in foster care: Evidence of deficits. Dev. Psychopathol. 2005, 17, 47–65. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Organized Attachment (N = 11, 4 F) Mean (SD) | Unresolved/ Disorganized Attachment (N = 7, 4 F) Mean (SD) | Mann-Whitney’s U | p Value | Cohen’s d | |
|---|---|---|---|---|---|
| PID-5—Negative Affect | 1.024 (0.43) | 1.118 (0.65) | 35.000 | 0.751 | 0.171 |
| PID-5—Detachment | 0.504 (0.33) | 0.564 (0.78) | 46.000 | 0.497 | 0.100 |
| PID-5—Antagonism | 0.393 (0.31) | 0.383 (0.30) | 39.000 | 0.964 | 0.033 |
| PID-5—Disinhibition | 0.688 (0.42) | 0.842 (0.61) | 31.000 | 0.497 | 0.294 |
| PID-5—Psychoticism | 0.462 (0.53) | 0.900 (0.83) | 25.000 | 0.221 | 0.629 |
| SCL-90-R—Somatization | 46.909 (4.93) | 51.714 (9.30) | 25.000 | 0.219 | 0.646 |
| SCL-90-R—Obsessive-Compulsive | 47.000 (8.96) | 53.857 (11.98) | 23.500 | 0.173 | 0.648 |
| SCL-90-R—Interpersonal Sensitivity | 48.182 (9.03) | 51.857 (12.17) | 31.000 | 0.492 | 0.343 |
| SCL-90-R—Depression | 50.818 (11.64) | 54.286 (12.91) | 33.500 | 0.649 | 0.282 |
| SCL-90-R—Anxiety | 52.636 (11.23) | 52.143 (10.72) | 37.000 | 0.892 | 0.045 |
| SCL-90-R—Hostility | 49.455 (12.84) | 54.143 (11.768) | 24.500 | 0.203 | 0.381 |
| SCL-90-R—Phobic Anxiety | 49.273 (5.62) | 53.857 (13.40) | 34.000 | 0.681 | 0.446 |
| SCL-90-R—Paranoid Ideation | 46.636 (10.25) | 54.429 (12.05) | 23.500 | 0.173 | 0.697 |
| SCL-90-R—Psychoticism | 46.727 (8.34) | 50.429 (10.15) | 29.000 | 0.384 | 0.399 |
| SCL-90-R—Global Score | 49.455 (10.59) | 52.571 (12.39) | 31.000 | 0.496 | 0.270 |
| Seed/Target | x | y | z | Cluster Size | Cluster p-FDR | Cluster p-unc | Peak p-unc | Effect |
|---|---|---|---|---|---|---|---|---|
| Amygdala LH | ||||||||
| Lobule VIII LH | −32 | −56 | −56 | 64 | 0.018471 | 0.000324 | 0.000028 | − |
| Amygdala RH | ||||||||
| ACC | 2 | 44 | 16 | 57 | 0.037827 | 0.000714 | 0.000066 | + |
| Hippocampus RH | ||||||||
| pMTG RH | 54 | −64 | 6 | 86 | 0.002723 | 0.000066 | 0.000007 | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruciani, G.; Boccia, M.; Lingiardi, V.; Giovanardi, G.; Zingaretti, P.; Spitoni, G.F. An Exploratory Study on Resting-State Functional Connectivity in Individuals with Disorganized Attachment: Evidence for Key Regions in Amygdala and Hippocampus. Brain Sci. 2021, 11, 1539. https://doi.org/10.3390/brainsci11111539
Cruciani G, Boccia M, Lingiardi V, Giovanardi G, Zingaretti P, Spitoni GF. An Exploratory Study on Resting-State Functional Connectivity in Individuals with Disorganized Attachment: Evidence for Key Regions in Amygdala and Hippocampus. Brain Sciences. 2021; 11(11):1539. https://doi.org/10.3390/brainsci11111539
Chicago/Turabian StyleCruciani, Gianluca, Maddalena Boccia, Vittorio Lingiardi, Guido Giovanardi, Pietro Zingaretti, and Grazia Fernanda Spitoni. 2021. "An Exploratory Study on Resting-State Functional Connectivity in Individuals with Disorganized Attachment: Evidence for Key Regions in Amygdala and Hippocampus" Brain Sciences 11, no. 11: 1539. https://doi.org/10.3390/brainsci11111539
APA StyleCruciani, G., Boccia, M., Lingiardi, V., Giovanardi, G., Zingaretti, P., & Spitoni, G. F. (2021). An Exploratory Study on Resting-State Functional Connectivity in Individuals with Disorganized Attachment: Evidence for Key Regions in Amygdala and Hippocampus. Brain Sciences, 11(11), 1539. https://doi.org/10.3390/brainsci11111539