
Decreased Global EEG Synchronization in Amyloid Positive Mild Cognitive Impairment and Alzheimer’s Disease Patients—Relationship to APOE ε4

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. CSF Analysis
2.3. Resting-State EEG Recordings and Analyses
2.4. Statistical Analysis
3. Results
3.1. Demographics and Clinical Characteristics
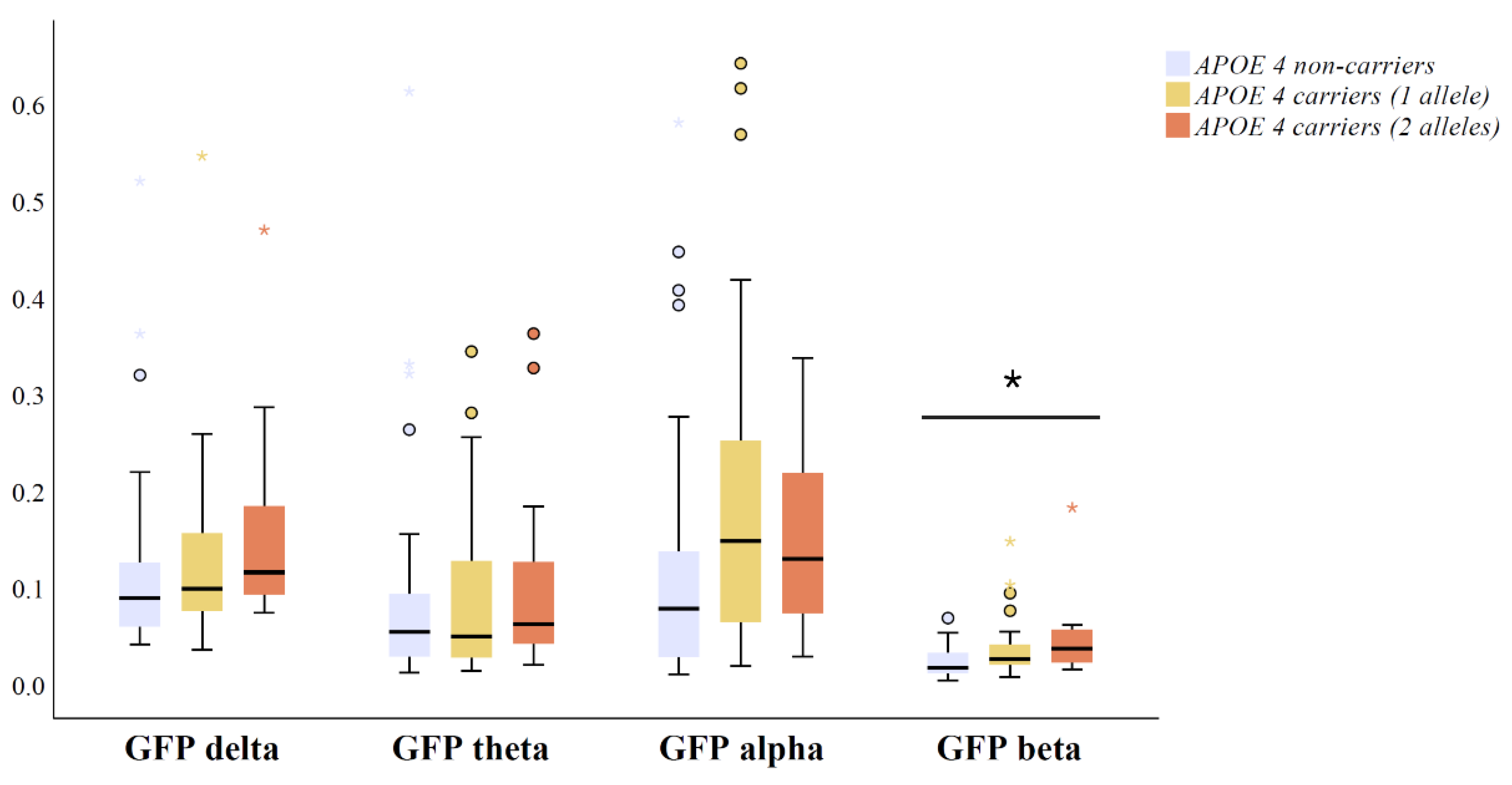
3.2. Relationship between Global EEG Power and APOE Genotype in Amyloid Positive MCI and AD Patients
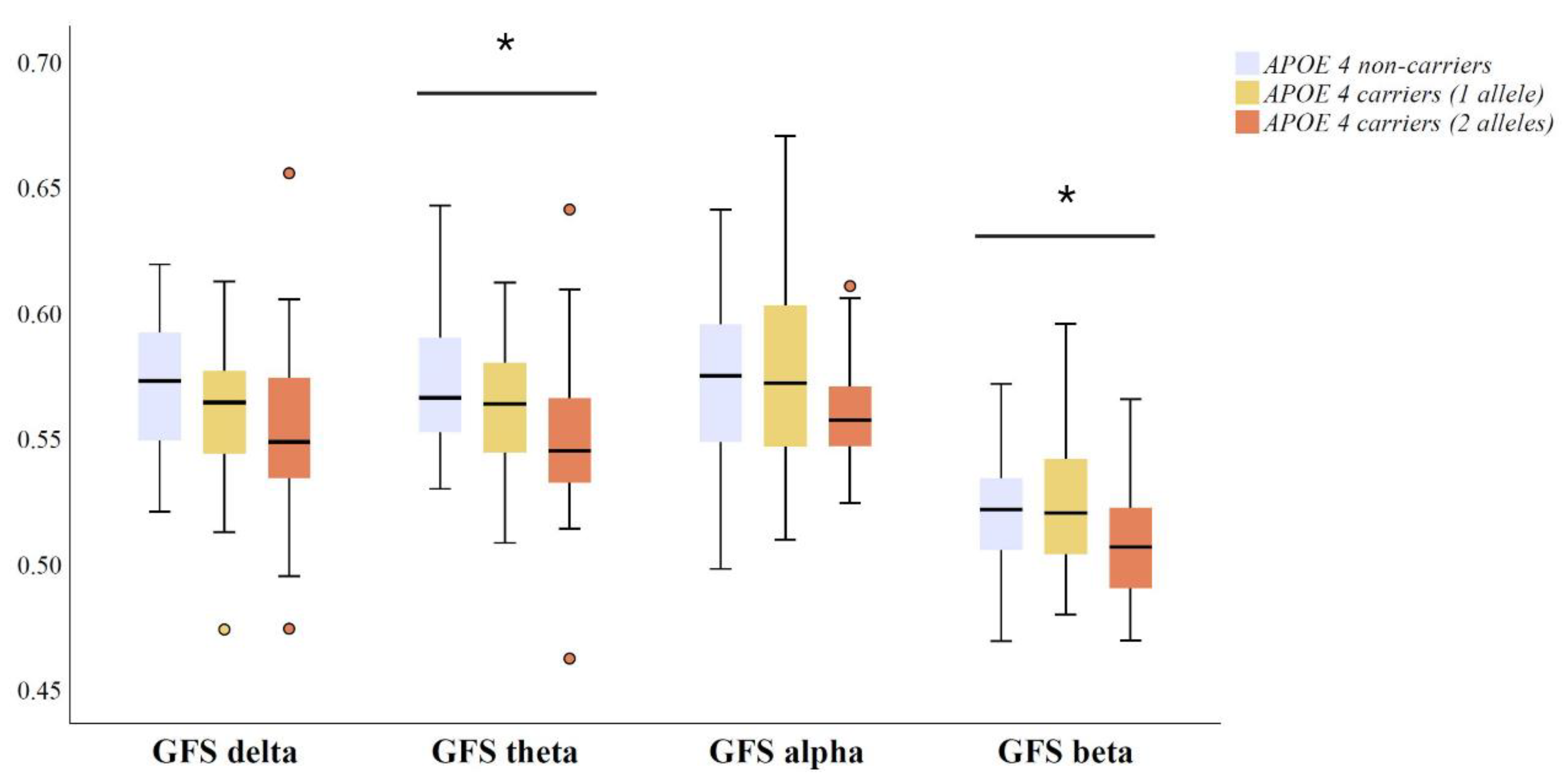
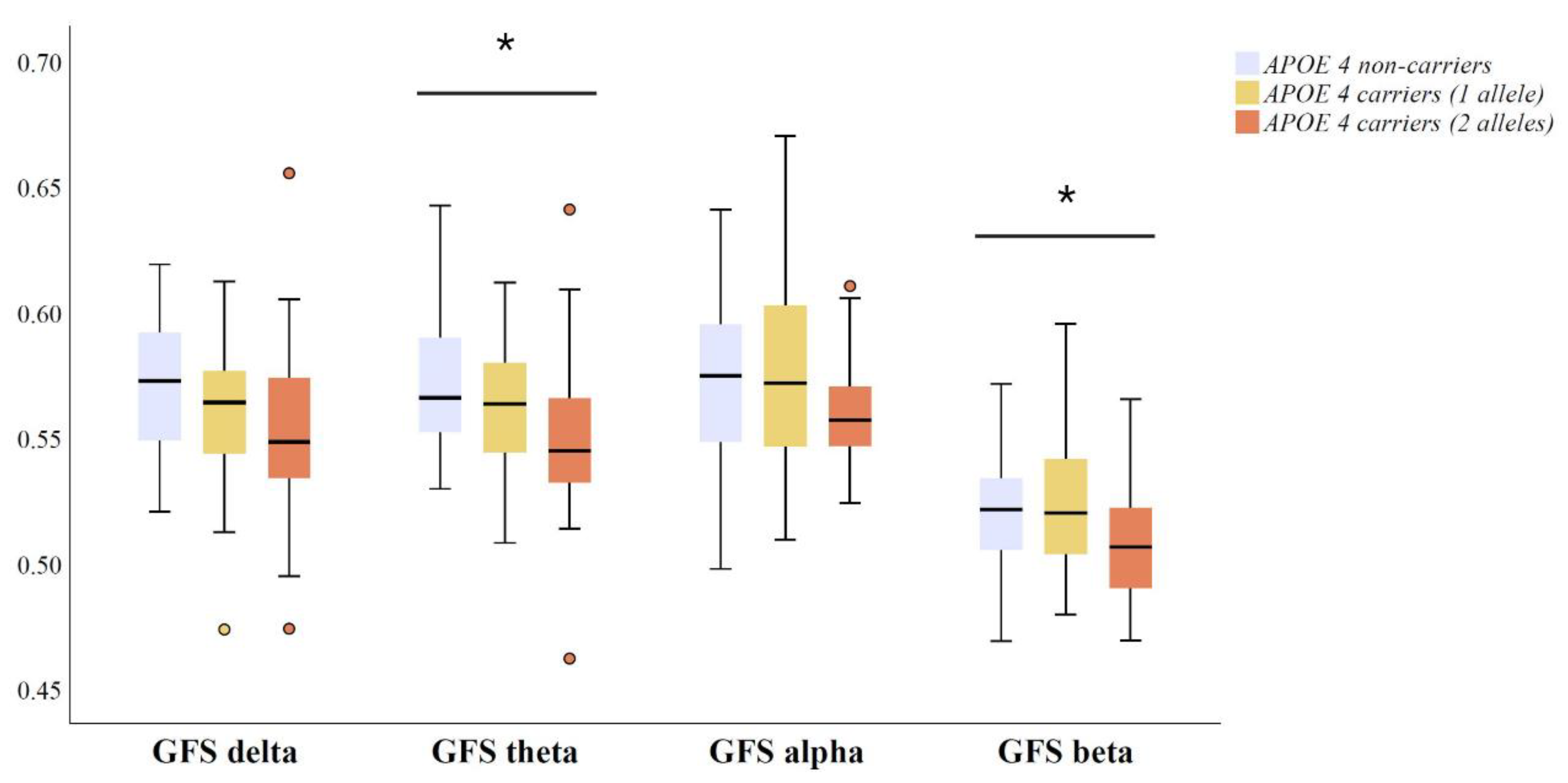
3.3. Relationship between Global EEG Synchronization and APOE Genotype in Amyloid Positive MCI and AD Patients
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef]
- Dubois, B.; Feldman, H.H.; Jacova, C.; Hampel, H.; Molinuevo, J.L.; Blennow, K.; DeKosky, S.T.; Gauthier, S.; Selkoe, D.; Bateman, R.; et al. Advancing research diagnostic criteria for Alzheimer’s disease: The IWG-2 criteria. Lancet Neurol. 2014, 13, 614–629. [Google Scholar] [CrossRef]
- Lourida, I.; Hannon, E.; Littlejohns, T.J.; Langa, K.M.; Hyppönen, E.; Kuzma, E.; Llewellyn, D.J. Association of Lifestyle and Genetic Risk With Incidence of Dementia. JAMA 2019, 322, 430–437. [Google Scholar] [CrossRef]
- Grant, W.B.; Campbell, A.; Itzhaki, R.F.; Savory, J. The significance of environmental factors in the etiology of Alzheimer’s disease. J. Alzheimers Dis. 2002, 4, 179–189. [Google Scholar] [CrossRef]
- Karch, C.M.; Goate, A.M. Alzheimer’s disease risk genes and mechanisms of disease pathogenesis. Biol. Psychiatry 2015, 77, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Farrer, L.A.; Cupples, L.A.; Haines, J.L.; Hyman, B.; Kukull, W.A.; Mayeux, R.; Myers, R.H.; Pericak-Vance, M.A.; Risch, N.; van Duijn, C.M. Effects of age, sex, and ethnicity on the association between apolipoprotein E genotype and Alzheimer disease. A meta-analysis. APOE and Alzheimer Disease Meta Analysis Consortium. JAMA 1997, 278, 1349–1356. [Google Scholar] [CrossRef]
- Liu, C.C.; Liu, C.C.; Kanekiyo, T.; Xu, H.; Bu, G. Apolipoprotein E and Alzheimer disease: Risk, mechanisms and therapy. Nat. Rev. Neurol. 2013, 9, 106–118. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Basak, J.M.; Holtzman, D.M. The role of apolipoprotein E in Alzheimer’s disease. Neuron 2009, 63, 287–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiman, E.M.; Chen, K.; Liu, X.; Bandy, D.; Yu, M.; Lee, W.; Ayutyanont, N.; Keppler, J.; Reeder, S.A.; Langbaum, J.B.; et al. Fibrillar amyloid-beta burden in cognitively normal people at 3 levels of genetic risk for Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2009, 106, 6820–6825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano, J.M.; Kim, J.; Stewart, F.R.; Jiang, H.; DeMattos, R.B.; Patterson, B.W.; Fagan, A.M.; Morris, J.C.; Mawuenyega, K.G.; Cruchaga, C.; et al. Human apoE isoforms differentially regulate brain amyloid-beta peptide clearance. Sci. Transl. Med. 2011, 3, 89ra57. [Google Scholar] [CrossRef] [Green Version]
- Brecht, W.J.; Harris, F.M.; Chang, S.; Tesseur, I.; Yu, G.Q.; Xu, Q.; Dee Fish, J.; Wyss-Coray, T.; Buttini, M.; Mucke, L.; et al. Neuron-specific apolipoprotein e4 proteolysis is associated with increased tau phosphorylation in brains of transgenic mice. J. Neurosci. 2004, 24, 2527–2534. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.R.; Morgan, D.; Mance, J.; Matthew, W.D.; Laskowitz, D.T. Apolipoprotein E modulates glial activation and the endogenous central nervous system inflammatory response. J. Neuroimmunol. 2001, 114, 107–113. [Google Scholar] [CrossRef]
- Sheline, Y.I.; Morris, J.C.; Snyder, A.Z.; Price, J.L.; Yan, Z.; D’Angelo, G.; Liu, C.; Dixit, S.; Benzinger, T.; Fagan, A.; et al. APOE4 allele disrupts resting state fMRI connectivity in the absence of amyloid plaques or decreased CSF Aβ42. J. Neurosci. 2010, 30, 17035–17040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippini, N.; MacIntosh, B.J.; Hough, M.G.; Goodwin, G.M.; Frisoni, G.B.; Smith, S.M.; Matthews, P.M.; Beckmann, C.F.; Mackay, C.E. Distinct patterns of brain activity in young carriers of the APOE-ε4 allele. Proc. Natl. Acad. Sci. USA 2009, 106, 7209–7214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stassen, H.H.; Lykken, D.T.; Propping, P.; Bomben, G. Genetic determination of the human EEG. Hum. Genet. 1988, 80, 165–176. [Google Scholar] [CrossRef]
- Lykken, D.T.; Tellegen, A.; Thorkelson, K. Genetic determination of EEG frequency spectra. Biol. Psychol. 1974, 1, 245–259. [Google Scholar] [CrossRef]
- Smit, D.J.; Posthuma, D.; Boomsma, D.I.; Geus, E.J. Heritability of background EEG across the power spectrum. Psychophysiology 2005, 42, 691–697. [Google Scholar] [CrossRef]
- Schomer, D.L.; Lopes da Silva, F. Niedermeyer’s Electroencephalography: Basic Principles, Clinical Applications, and Related Fields; Wolters Kluwer Health: Philadelphia, PA, USA, 2015. [Google Scholar]
- Smailovic, U.; Jelic, V. Neurophysiological Markers of Alzheimer’s Disease: Quantitative EEG Approach. Neurol. Ther. 2019, 8, 37–55. [Google Scholar] [CrossRef] [Green Version]
- van Beijsterveldt, C.E.; Molenaar, P.C.; de Geus, E.J.; Boomsma, D.I. Heritability of human brain functioning as assessed by electroencephalography. Am. J. Hum. Genet. 1996, 58, 562–573. [Google Scholar]
- van Beijsterveldt, C.E.; van Baal, G.C. Twin and family studies of the human electroencephalogram: A review and a meta-analysis. Biol. Psychol. 2002, 61, 111–138. [Google Scholar] [CrossRef]
- Selkoe, D.J. Alzheimer’s disease is a synaptic failure. Science 2002, 298, 789–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koffie, R.M.; Hyman, B.T.; Spires-Jones, T.L. Alzheimer’s disease: Synapses gone cold. Mol. Neurodegener. 2011, 6, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, C.M. Electrical Neuroimaging; Cambridge University Press: Cambridge, UK, 2009; 238p. [Google Scholar]
- Dierks, T.; Ihl, R.; Frolich, L.; Maurer, K. Dementia of the Alzheimer type: Effects on the spontaneous EEG described by dipole sources. Psychiatry Res. 1993, 50, 151–162. [Google Scholar] [CrossRef]
- Jelic, V.; Johansson, S.E.; Almkvist, O.; Shigeta, M.; Julin, P.; Nordberg, A.; Winblad, B.; Wahlund, L.O. Quantitative electroencephalography in mild cognitive impairment: Longitudinal changes and possible prediction of Alzheimer’s disease. Neurobiol. Aging 2000, 21, 533–540. [Google Scholar] [CrossRef]
- Huang, C.; Wahlund, L.; Dierks, T.; Julin, P.; Winblad, B.; Jelic, V. Discrimination of Alzheimer’s disease and mild cognitive impairment by equivalent EEG sources: A cross-sectional and longitudinal study. Clin. Neurophysiol. 2000, 111, 1961–1967. [Google Scholar] [CrossRef]
- Koenig, T.; Prichep, L.; Dierks, T.; Hubl, D.; Wahlund, L.O.; John, E.R.; Jelic, V. Decreased EEG synchronization in Alzheimer’s disease and mild cognitive impairment. Neurobiol. Aging 2005, 26, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Jelic, V.; Shigeta, M.; Julin, P.; Almkvist, O.; Winblad, B.; Wahlund, L.O. Quantitative electroencephalography power and coherence in Alzheimer’s disease and mild cognitive impairment. Dementia 1996, 7, 314–323. [Google Scholar] [CrossRef]
- Adler, G.; Brassen, S.; Jajcevic, A. EEG coherence in Alzheimer’s dementia. J. Neural Transm. 2003, 110, 1051–1058. [Google Scholar] [CrossRef]
- Pozzi, D.; Petracchi, M.; Sabe, L.; Golimstock, A.; Garcia, H.; Starkstein, S. Quantified electroencephalographic correlates of neuropsychological deficits in Alzheimer’s disease. J. Neuropsychiatry Clin. Neurosci. 1995, 7, 61–67. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, S.H.; Park, G.; Kim, S.; Bae, S.M.; Kim, D.W.; Im, C.H. Clinical implications of quantitative electroencephalography and current source density in patients with Alzheimer’s disease. Brain Topogr. 2012, 25, 461–474. [Google Scholar] [CrossRef]
- Park, Y.-M.; Che, H.-J.; Im, C.-H.; Jung, H.-T.; Bae, S.-M.; Lee, S.-H. Decreased EEG synchronization and its correlation with symptom severity in Alzheimer’s disease. Neurosci. Res. 2008, 62, 112–117. [Google Scholar] [CrossRef]
- Ma, C.C.; Liu, A.J.; Liu, A.H.; Zhou, X.Y.; Zhou, S.N. Electroencephalogram global field synchronization analysis: A new method for assessing the progress of cognitive decline in Alzheimer’s disease. Clin. EEG Neurosci. 2014, 45, 98–103. [Google Scholar] [CrossRef]
- Smailovic, U.; Kåreholt, I.; Koenig, T.; Ashton, N.J.; Winblad, B.; Höglund, K.; Nilsson, P.; Zetterberg, H.; Blennow, K.; Jelic, V. Synaptic molecular and neurophysiological markers are independent predictors of progression in Alzheimer’s disease. J. Alzheimers Dis. 2021, 83, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Luckhaus, C.; Grass-Kapanke, B.; Blaeser, I.; Ihl, R.; Supprian, T.; Winterer, G.; Zielasek, J.; Brinkmeyer, J. Quantitative EEG in progressing vs stable mild cognitive impairment (MCI): Results of a 1-year follow-up study. Int. J. Geriatr. Psychiatry 2008, 23, 1148–1155. [Google Scholar] [CrossRef]
- Smailovic, U.; Koenig, T.; Kareholt, I.; Andersson, T.; Kramberger, M.G.; Winblad, B.; Jelic, V. Quantitative EEG power and synchronization correlate with Alzheimer’s disease CSF biomarkers. Neurobiol. Aging 2018, 63, 88–95. [Google Scholar] [CrossRef]
- Stomrud, E.; Hansson, O.; Minthon, L.; Blennow, K.; Rosen, I.; Londos, E. Slowing of EEG correlates with CSF biomarkers and reduced cognitive speed in elderly with normal cognition over 4 years. Neurobiol. Aging 2010, 31, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehtovirta, M.; Partanen, J.; Könönen, M.; Hiltunen, J.; Helisalmi, S.; Hartikainen, P.; Riekkinen Sr, P.; Soininen, H. A Longitudinal Quantitative EEG Study of Alzheimer’s Disease: Relation to Apolipoprotein E Polymorphism. Dement. Geriatr. Cogn. Disord. 2000, 11, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Babiloni, C.; Benussi, L.; Binetti, G.; Cassetta, E.; Dal Forno, G.; Del Percio, C.; Ferreri, F.; Ferri, R.; Frisoni, G.; Ghidoni, R.; et al. Apolipoprotein E and alpha brain rhythms in mild cognitive impairment: A multicentric Electroencephalogram study. Ann. Neurol. 2006, 59, 323–334. [Google Scholar] [CrossRef]
- de Waal, H.; Stam, C.J.; de Haan, W.; van Straaten, E.C.; Blankenstein, M.A.; Scheltens, P.; van der Flier, W.M. Alzheimer’s disease patients not carrying the apolipoprotein E epsilon4 allele show more severe slowing of oscillatory brain activity. Neurobiol. Aging 2013, 34, 2158–2163. [Google Scholar] [CrossRef]
- Jelic, V.; Julin, P.; Shigeta, M.; Nordberg, A.; Lannfelt, L.; Winblad, B.; Wahlund, L.O. Apolipoprotein E epsilon4 allele decreases functional connectivity in Alzheimer’s disease as measured by EEG coherence. J. Neurol. Neurosurg. Psychiatry 1997, 63, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Winblad, B.; Palmer, K.; Kivipelto, M.; Jelic, V.; Fratiglioni, L.; Wahlund, L.O.; Nordberg, A.; Bäckman, L.; Albert, M.; Almkvist, O.; et al. Mild cognitive impairment—beyond controversies, towards a consensus: Report of the International Working Group on Mild Cognitive Impairment. J. Intern. Med. 2004, 256, 240–246. [Google Scholar] [CrossRef]
- World Health Organization. The ICD-10 Classification of Mental and Behavioural Disorders: Clinical Descriptions and Diagnostic Guidelines; WHO: Geneva, Switzerland, 1992. [Google Scholar]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimers Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Olsson, A.; Vanderstichele, H.; Andreasen, N.; De Meyer, G.; Wallin, A.; Holmberg, B.; Rosengren, L.; Vanmechelen, E.; Blennow, K. Simultaneous measurement of beta-amyloid(1-42), total tau, and phosphorylated tau (Thr181) in cerebrospinal fluid by the xMAP technology. Clin. Chem. 2005, 51, 336–345. [Google Scholar] [CrossRef]
- Motter, R.; Vigo-Pelfrey, C.; Kholodenko, D.; Barbour, R.; Johnson-Wood, K.; Galasko, D.; Chang, L.; Miller, B.; Clark, C.; Green, R.; et al. Reduction of beta-amyloid peptide42 in the cerebrospinal fluid of patients with Alzheimer’s disease. Ann. Neurol. 1995, 38, 643–648. [Google Scholar] [CrossRef]
- Lehmann, D.; Skrandies, W. Reference-free identification of components of checkerboard-evoked multichannel potential fields. Electroencephalogr. Clin. Neurophysiol. 1980, 48, 609–621. [Google Scholar] [CrossRef]
- Koenig, T.; Lehmann, D.; Saito, N.; Kuginuki, T.; Kinoshita, T.; Koukkou, M. Decreased functional connectivity of EEG theta-frequency activity in first-episode, neuroleptic-naive patients with schizophrenia: Preliminary results. Schizophr. Res. 2001, 50, 55–60. [Google Scholar] [CrossRef]
- Fonseca, L.C.; Tedrus, G.M.; Prandi, L.R.; Andrade, A.C. Quantitative electroencephalography power and coherence measurements in the diagnosis of mild and moderate Alzheimer’s disease. Arquivos de Neuro-Psiquiatria 2011, 69, 297–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stam, C.J.; Van Der Made, Y.; Pijnenburg, Y.A.L.; Scheltens, P. EEG synchronization in mild cognitive impairment and Alzheimer’s disease. Acta Neurologica Scandinavica 2003, 108, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Stam, C.J.; Nolte, G.; Daffertshofer, A. Phase lag index: Assessment of functional connectivity from multi channel EEG and MEG with diminished bias from common sources. Hum. Brain Mapp. 2007, 28, 1178–1193. [Google Scholar] [CrossRef]
- Axmacher, N.; Schmitz, D.P.; Wagner, T.; Elger, C.E.; Fell, J. Interactions between medial temporal lobe, prefrontal cortex, and inferior temporal regions during visual working memory: A combined intracranial EEG and functional magnetic resonance imaging study. J. Neurosci. 2008, 28, 7304–7312. [Google Scholar] [CrossRef] [PubMed]
- Hanslmayr, S.; Spitzer, B.; Bäuml, K.H. Brain oscillations dissociate between semantic and nonsemantic encoding of episodic memories. Cereb. Cortex 2009, 19, 1631–1640. [Google Scholar] [CrossRef] [Green Version]
- Hanslmayr, S.; Staresina, B.P.; Bowman, H. Oscillations and Episodic Memory: Addressing the Synchronization/Desynchronization Conundrum. Trends Neurosci. 2016, 39, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.; Mueller, H.M. “Too Many betas do not Spoil the Broth”: The Role of Beta Brain Oscillations in Language Processing. Front. Psychol. 2012, 3, 201. [Google Scholar] [CrossRef] [Green Version]
- Piantoni, G.; Kline, K.A.; Eagleman, D.M. Beta oscillations correlate with the probability of perceiving rivalrous visual stimuli. J. Vis. 2010, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Wimmer, K.; Ramon, M.; Pasternak, T.; Compte, A. Transitions between Multiband Oscillatory Patterns Characterize Memory-Guided Perceptual Decisions in Prefrontal Circuits. J. Neurosci. 2016, 36, 489–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamiński, J.; Brzezicka, A.; Gola, M.; Wróbel, A. β band oscillations engagement in human alertness process. Int. J. Psychophysiol. 2012, 85, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Gola, M.; Kamiński, J.; Brzezicka, A.; Wróbel, A. β band oscillations as a correlate of alertness--changes in aging. Int. J. Psychophysiol. 2012, 85, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.; Nordberg, A.; Westman, E. Biological subtypes of Alzheimer disease: A systematic review and meta-analysis. Neurology 2020, 94, 436–448. [Google Scholar] [CrossRef] [Green Version]
- Lehtovirta, M.; Partanen, J.; Könönen, M.; Soininen, H.; Helisalmi, S.; Mannermaa, A.; Ryynänen, M.; Hartikainen, P.; Riekkinen, P. Spectral analysis of EEG in Alzheimer’s disease: Relation to apolipoprotein E polymorphism. Neurobiol. Aging 1996, 17, 523–526. [Google Scholar] [CrossRef]
- Ponomareva, N.V.; Korovaitseva, G.I.; Rogaev, E.I. EEG alterations in non-demented individuals related to apolipoprotein E genotype and to risk of Alzheimer disease. Neurobiol. Aging 2008, 29, 819–827. [Google Scholar] [CrossRef]
- Olsson, B.; Lautner, R.; Andreasson, U.; Öhrfelt, A.; Portelius, E.; Bjerke, M.; Hölttä, M.; Rosén, C.; Olsson, C.; Strobel, G.; et al. CSF and blood biomarkers for the diagnosis of Alzheimer’s disease: A systematic review and meta-analysis. Lancet Neurol. 2016, 15, 673–684. [Google Scholar] [CrossRef]
- Mitchell, A.J.; Shiri-Feshki, M. Rate of progression of mild cognitive impairment to dementia—Meta-analysis of 41 robust inception cohort studies. Acta Psychiatr. Scand. 2009, 119, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.C. Mild cognitive impairment as a diagnostic entity. J. Intern. Med. 2004, 256, 183–194. [Google Scholar] [CrossRef]
- DeTure, M.A.; Dickson, D.W. The neuropathological diagnosis of Alzheimer’s disease. Mol. Neurodegener. 2019, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Crary, J.F. Primary age-related tauopathy and the amyloid cascade hypothesis: The exception that proves the rule? J. Neurol. Neuromedicine 2016, 1, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Jack, C.R., Jr. PART and SNAP. Acta Neuropathol. 2014, 128, 773–776. [Google Scholar] [CrossRef] [Green Version]
- Tsuang, D.; Leverenz, J.B.; Lopez, O.L.; Hamilton, R.L.; Bennett, D.A.; Schneider, J.A.; Buchman, A.S.; Larson, E.B.; Crane, P.K.; Kaye, J.A.; et al. APOE ε4 increases risk for dementia in pure synucleinopathies. JAMA Neurol. 2013, 70, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Chuang, Y.F.; Hayden, K.M.; Norton, M.C.; Tschanz, J.; Breitner, J.C.; Welsh-Bohmer, K.A.; Zandi, P.P. Association between APOE epsilon4 allele and vascular dementia: The Cache County study. Dement. Geriatr. Cogn. Disord. 2010, 29, 248–253. [Google Scholar] [CrossRef] [Green Version]
- Müller, E.G.; Edwin, T.H.; Stokke, C.; Navelsaker, S.S.; Babovic, A.; Bogdanovic, N.; Knapskog, A.B.; Revheim, M.E. Amyloid-β PET-Correlation with cerebrospinal fluid biomarkers and prediction of Alzheimer´s disease diagnosis in a memory clinic. PLoS ONE 2019, 14, e0221365. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| APOE ε4 Non-Carriers | APOEε4 Heterozygous Carriers (One Allele) | APOEε4 Homozygous Carriers (Two Alleles) | p-Value | |
|---|---|---|---|---|
| N (total) | 31 | 42 | 28 | |
| MCI | 13 | 22 | 15 | |
| AD | 18 | 20 | 13 | |
| Age (years) | 65.03 ± 9.17 | 65.79 ± 8.54 | 64.04 ± 5.31 | 0.766 |
| Sex (M/F) | 15/16 | 17/25 | 9/19 | 0.447 |
| Education (years) | 11.97 ± 3.80 | 12.39 ± 3.80 | 12.68 ± 3.42 | 0.631 |
| MMSE a | 24.73 ± 4.32 | 26.71 ± 2.76 | 25.11 ± 4.14 | 0.092 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smailovic, U.; Johansson, C.; Koenig, T.; Kåreholt, I.; Graff, C.; Jelic, V. Decreased Global EEG Synchronization in Amyloid Positive Mild Cognitive Impairment and Alzheimer’s Disease Patients—Relationship to APOE ε4. Brain Sci. 2021, 11, 1359. https://doi.org/10.3390/brainsci11101359
Smailovic U, Johansson C, Koenig T, Kåreholt I, Graff C, Jelic V. Decreased Global EEG Synchronization in Amyloid Positive Mild Cognitive Impairment and Alzheimer’s Disease Patients—Relationship to APOE ε4. Brain Sciences. 2021; 11(10):1359. https://doi.org/10.3390/brainsci11101359
Chicago/Turabian StyleSmailovic, Una, Charlotte Johansson, Thomas Koenig, Ingemar Kåreholt, Caroline Graff, and Vesna Jelic. 2021. "Decreased Global EEG Synchronization in Amyloid Positive Mild Cognitive Impairment and Alzheimer’s Disease Patients—Relationship to APOE ε4" Brain Sciences 11, no. 10: 1359. https://doi.org/10.3390/brainsci11101359
APA StyleSmailovic, U., Johansson, C., Koenig, T., Kåreholt, I., Graff, C., & Jelic, V. (2021). Decreased Global EEG Synchronization in Amyloid Positive Mild Cognitive Impairment and Alzheimer’s Disease Patients—Relationship to APOE ε4. Brain Sciences, 11(10), 1359. https://doi.org/10.3390/brainsci11101359