
Reduced Axon Calibre in the Associative Striatum of the Sapap3 Knockout Mouse

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
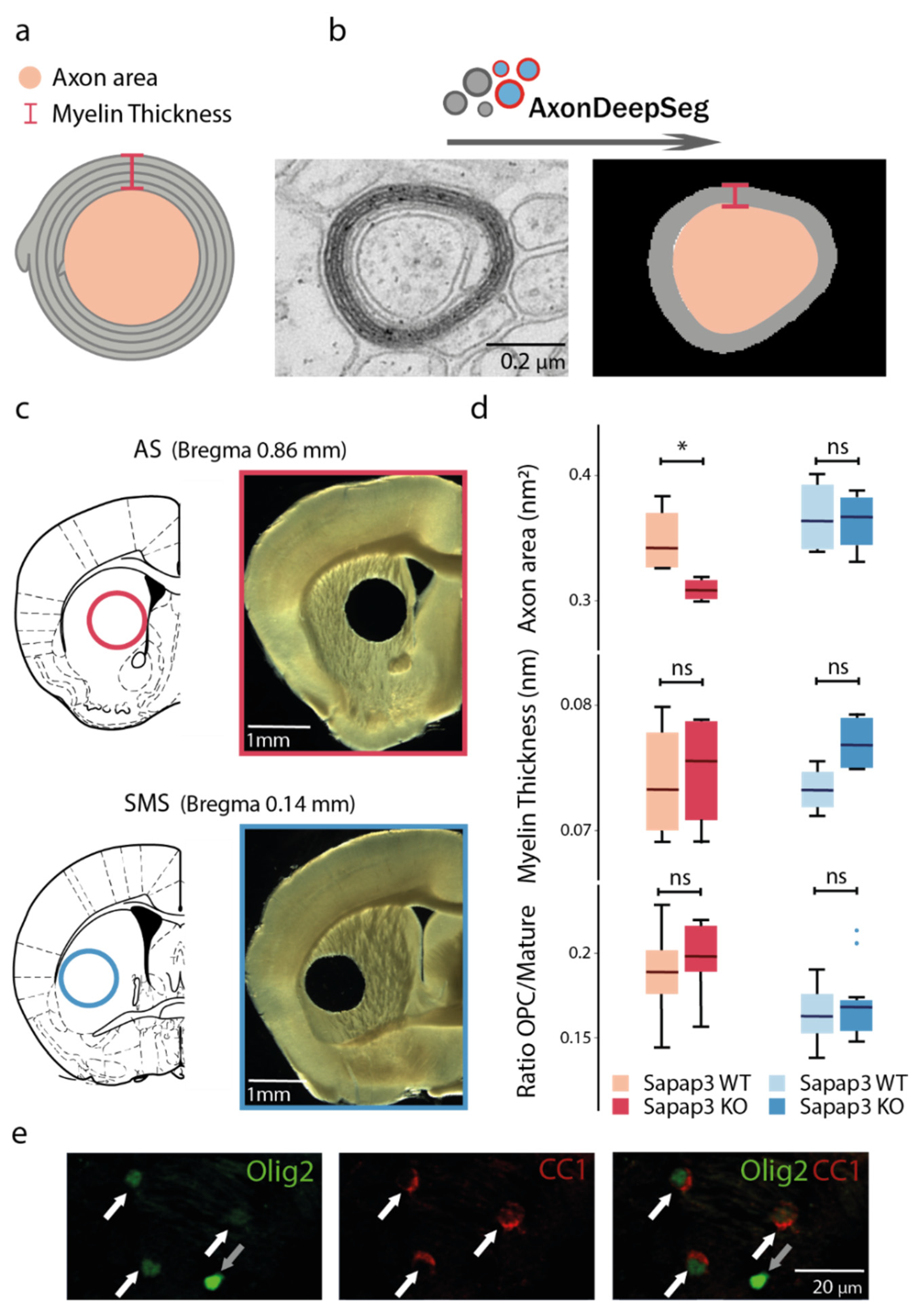
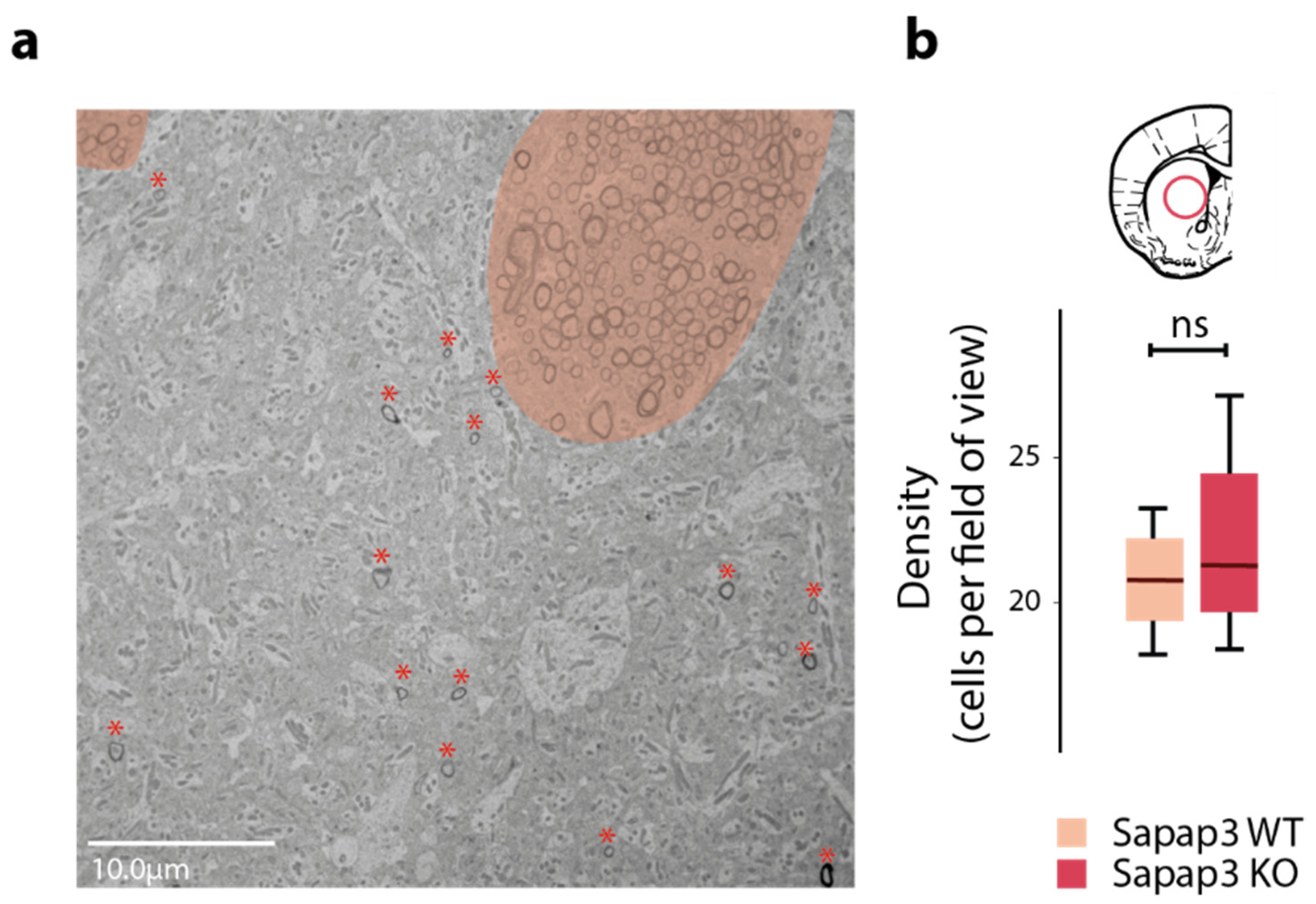
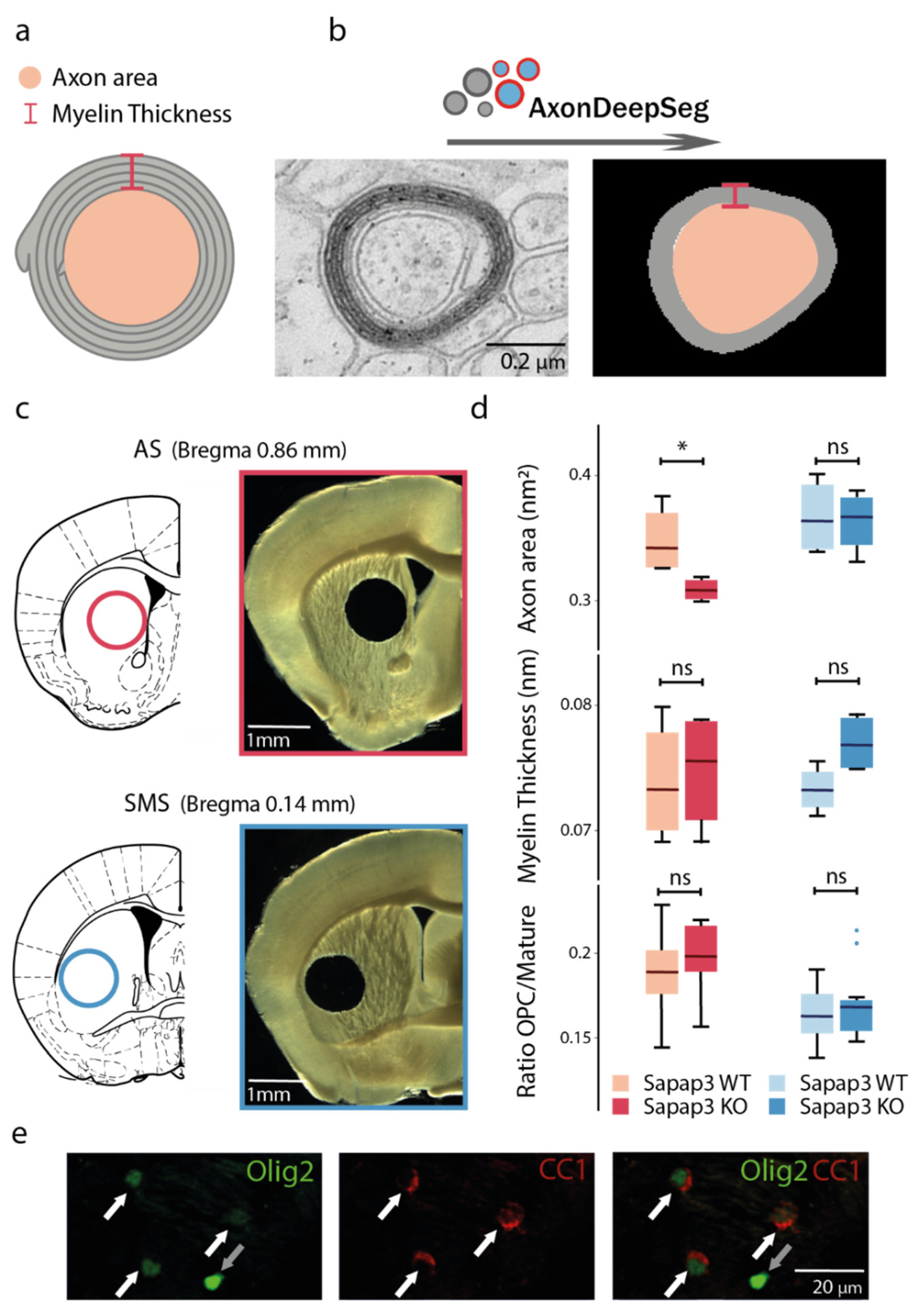
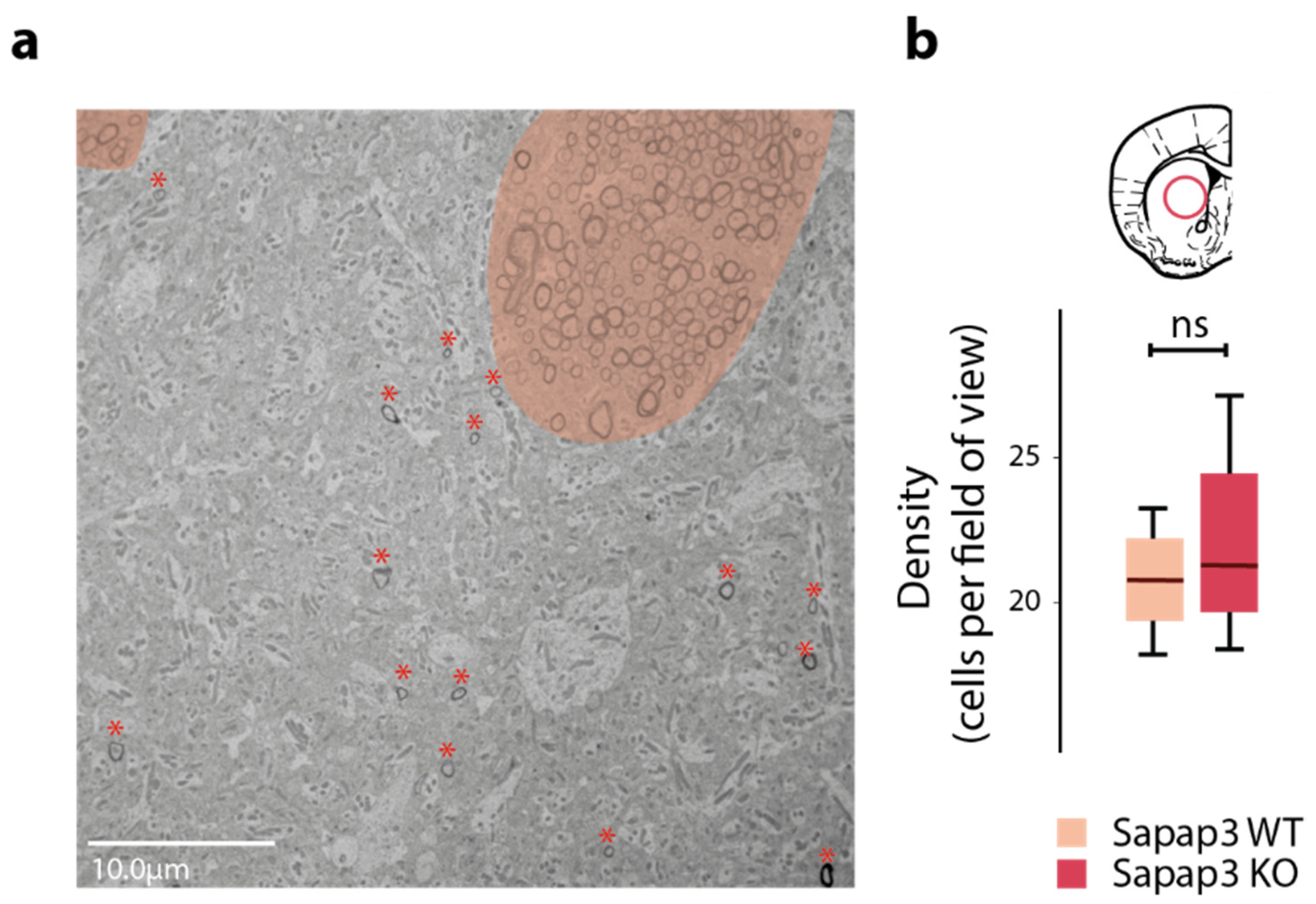
3.1. Axon Calibre Is Diminished in the Associative but Not in the Sensorimotor Striatum of Sapap3-KO Mice
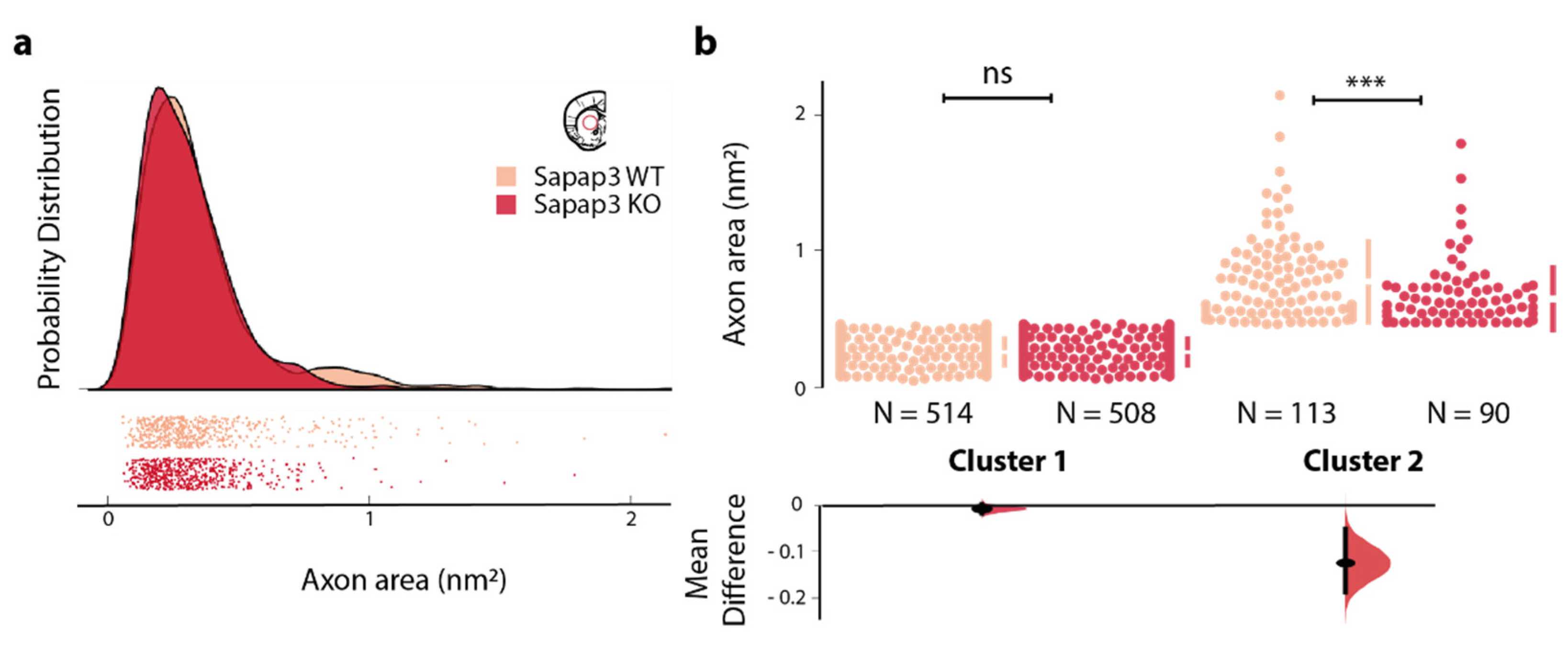
3.2. The Reduction in Axon Calibre Arises from a Subpopulation of Axons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Logan, G.D. Skill and automaticity: Relations, implications, and future directions. Can. J. Psychol. Rev. Can. Psychol. 1985, 39, 367–386. [Google Scholar] [CrossRef] [Green Version]
- Haith, A.M.; Krakauer, J.W. The multiple effects of practice: Skill, habit and reduced cognitive load. Curr. Opin. Behav. Sci. 2018, 20, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Langen, M.; Kas, M.J.H.; Staal, W.G.; van Engeland, H.; Durston, S. The neurobiology of repetitive behavior: Of mice. Neurosci. Biobehav. Rev. 2011, 35, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Vicente, A.M.; Martins, G.J.; Costa, R.M. Cortico-basal ganglia circuits underlying dysfunctional control of motor behaviors in neuropsychiatric disorders. Curr. Opin. Genet. Dev. 2020, 65, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.; Kim, S.-J. The pathophysiology of restricted repetitive behavior. J. Neurodev. Disord. 2009, 1, 114–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, G.E.; DeLong, M.R.; Strick, P.L. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci. 1986, 9, 357–381. [Google Scholar] [CrossRef]
- Lee, J.; Wang, W.; Sabatini, B.L. Anatomically segregated basal ganglia pathways allow parallel behavioral modulation. Nat. Neurosci. 2020, 23, 1388–1398. [Google Scholar] [CrossRef]
- Voorn, P.; Vanderschuren, L.J.M.J.; Groenewegen, H.J.; Robbins, T.W.; Pennartz, C.M.A. Putting a spin on the dorsal–ventral divide of the striatum. Trends Neurosci. 2004, 27, 468–474. [Google Scholar] [CrossRef]
- Tremblay, L.; Worbe, Y.; Thobois, S.; Sgambato-Faure, V.; Féger, J. Selective dysfunction of basal ganglia subterritories: From movement to behavioral disorders. Mov. Disord. 2015, 30, 1155–1170. [Google Scholar] [CrossRef]
- Greene, D.J.; Williams, A.C., III; Koller, J.M.; Schlaggar, B.L.; Black, K.J. Brain structure in pediatric Tourette syndrome. Mol. Psychiatry 2017, 22, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Müller-Vahl, K.R.; Grosskreutz, J.; Prell, T.; Kaufmann, J.; Bodammer, N.; Peschel, T. Tics are caused by alterations in prefrontal areas, thalamus and putamen, while changes in the cingulate gyrus reflect secondary compensatory mechanisms. BMC Neurosci. 2014, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Kobets, A.; Du, J.-C.; Lennington, J.; Li, L.; Banasr, M.; Duman, R.S.; Vaccarino, F.M.; DiLeone, R.J.; Pittenger, C. Targeted ablation of cholinergic interneurons in the dorsolateral striatum produces behavioral manifestations of Tourette syndrome. Proc. Natl. Acad. Sci. USA 2015, 112, 893–898. [Google Scholar] [CrossRef] [Green Version]
- Thomalla, G.; Siebner, H.R.; Jonas, M.; Bäumer, T.; Biermann-Ruben, K.; Hummel, F.; Gerloff, C.; Müller-Vahl, K.; Schnitzler, A.; Orth, M.; et al. Structural changes in the somatosensory system correlate with tic severity in gilles de la Tourette syndrome. Brain 2009, 132, 765–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worbe, Y.; Marrakchi-Kacem, L.; Lecomte, S.; Valabregue, R.; Poupon, F.; Guevara, P.; Tucholka, A.; Mangin, J.-F.; Vidailhet, M.; Lehericy, S.; et al. Altered structural connectivity of cortico-striato-pallido-thalamic networks in gilles de la Tourette syndrome. Brain 2015, 138, 472–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, L.R.; Phelps, M.E.; Mazziotta, J.C.; Guze, B.H.; Schwartz, J.M.; Selin, C.E. Local cerebral glucose metabolic rates in obsessive-compulsive disorder: A comparison with rates in unipolar depression and in normal controls. Arch. Gen. Psychiatry 1987, 44, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Brody, A.L.; Maidment, K.M.; Dunkin, J.J.; Colgan, M.; Alborzian, S.; Phelps, M.E.; Baxter, L.R. Localized orbitofrontal and subcortical metabolic changes and predictors of response to paroxetine treatment in obsessive-compulsive disorder. Neuropsychopharmacology 1999, 21, 683–693. [Google Scholar] [CrossRef]
- Whiteside, S.P.; Port, J.D.; Abramowitz, J.S. A meta—Analysis of functional neuroimaging in obsessive—Compulsive disorder. Psychiatry Res. Neuroimaging 2004, 132, 69–79. [Google Scholar] [CrossRef]
- van den Heuvel, O.A.; van Wingen, G.; Soriano-Mas, C.; Alonso, P.; Chamberlain, S.R.; Nakamae, T.; Denys, D.; Goudriaan, A.E.; Veltman, D.J. Brain Circuitry of Compulsivity. Eur. Neuropsychopharmacol. 2016, 26, 810–827. [Google Scholar] [CrossRef] [Green Version]
- Robbins, T.W.; Vaghi, M.M.; Banca, P. Obsessive-compulsive disorder: Puzzles and prospects. Neuron 2019, 102, 27–47. [Google Scholar] [CrossRef]
- Worbe, Y.; Mallet, L.; Golmard, J.-L.; Béhar, C.; Durif, F.; Jalenques, I.; Damier, P.; Derkinderen, P.; Pollak, P.; Anheim, M.; et al. Repetitive behaviours in patients with gilles de la Tourette syndrome: Tics, compulsions, or both? PLoS ONE 2010, 5, e12959. [Google Scholar] [CrossRef]
- Rothenberger, A.; Roessner, V. Psychopharmacotherapy of obsessive-compulsive symptoms within the framework of Tourette syndrome. Curr. Neuropharmacol. 2019, 17, 703–709. [Google Scholar] [CrossRef]
- Hartmann, A.; Millet, B. Repetitive movements and behaviors in neurological and psychiatric practice: Distinctions and similarities between Tourette disorder and obsessive—Compulsive disorder. Rev. Neurol. 2018, 174, 199–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddy, C.M.; Cavanna, A.E. Tourette syndrome and obsessive compulsive disorder: Compulsivity along the continuum. J. Obs. -Compuls. Relat. Disord. 2014, 3, 363–371. [Google Scholar] [CrossRef]
- Berridge, K.C.; Whishaw, I.Q. Cortex, striatum and cerebellum: Control of serial order in a grooming sequence. Exp. Brain Res. 1992, 90, 275–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalueff, A.V.; Stewart, A.M.; Song, C.; Berridge, K.C.; Graybiel, A.M.; Fentress, J.C. Neurobiology of rodent self-grooming and its value for translational neuroscience. Nat. Rev. Neurosci. 2016, 17, 45–59. [Google Scholar] [CrossRef] [Green Version]
- Shmelkov, S.V.; Hormigo, A.; Jing, D.; Proenca, C.C.; Bath, K.G.; Milde, T.; Shmelkov, E.; Kushner, J.S.; Baljevic, M.; Dincheva, I.; et al. Slitrk5 deficiency impairs corticostriatal circuitry and leads to obsessive-compulsive–like behaviors in mice. Nat. Med. 2010, 16, 598–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welch, J.M.; Lu, J.; Rodriguiz, R.M.; Trotta, N.C.; Peca, J.; Ding, J.-D.; Feliciano, C.; Chen, M.; Adams, J.P.; Luo, J.; et al. Cortico-striatal synaptic defects and OCD-like behaviours in Sapap3-mutant mice. Nature 2007, 448, 894–900. [Google Scholar] [CrossRef] [Green Version]
- Burguière, E.; Monteiro, P.; Feng, G.; Graybiel, A.M. Optogenetic stimulation of lateral orbitofronto-striatal pathway suppresses compulsive behaviors. Science 2013, 340, 1243–1246. [Google Scholar] [CrossRef] [Green Version]
- van den Boom, B.J.G.; Pavlidi, P.; Wolf, C.J.H.; Mooij, A.H.; Willuhn, I. Automated classification of self-grooming in mice using open-source software. J. Neurosci. Methods 2017, 289, 48–56. [Google Scholar] [CrossRef]
- Corbit, V.L.; Manning, E.E.; Gittis, A.H.; Ahmari, S.E. Strengthened inputs from secondary motor cortex to striatum in a mouse model of compulsive behavior. J. Neurosci. 2019, 39, 2965–2975. [Google Scholar] [CrossRef] [Green Version]
- Peñagarikano, O.; Abrahams, B.S.; Herman, E.I.; Winden, K.C.; Gdalyahu, A.; Dong, H.; Sonnenblick, L.I.; Gruver, R.; Almajano, J.; Bragin, A.; et al. Absence of CNTNAP2 leads to epilepsy, neuronal migration abnormalities, and core autism-related deficits. Cell 2011, 147, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Li, L.; Pittenger, C. Ablation of fast-spiking interneurons in the dorsal striatum, recapitulating abnormalities seen post-mortem in Tourette syndrome, produces anxiety and elevated grooming. Neuroscience 2016, 324, 321–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Y.; Ade, K.K.; Caffall, Z.; Ilcim Ozlu, M.; Eroglu, C.; Feng, G.; Calakos, N. Circuit-selective striatal synaptic dysfunction in the Sapap3 knockout mouse model of obsessive-compulsive disorder. Biol. Psychiatry 2014, 75, 623–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamothe, H.; Schreiweis, C.; Lavielle, O.; Mallet, L.; Burguiere, E. Not only compulsivity: The SAPAP3-KO mouse reconsidered as a comorbid model expressing a spectrum of pathological repetitive behaviors. bioRxiv 2021. [Google Scholar] [CrossRef]
- Rizzo, F.; Nespoli, E.; Abaei, A.; Bar-Gad, I.; Deelchand, D.K.; Fegert, J.; Rasche, V.; Hengerer, B.; Boeckers, T.M. Aripiprazole selectively reduces motor tics in a young animal model for Tourette’s syndrome and comorbid attention deficit and hyperactivity disorder. Front. Neurol. 2018, 9, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waxman, S.G. Determinants of conduction velocity in myelinated nerve fibers. Muscle Nerve 1980, 3, 141–150. [Google Scholar] [CrossRef]
- Friede, R.L. Relation between myelin sheath thickness, internode geometry, and sheath resistance. Exp. Neurol. 1986, 92, 234–247. [Google Scholar] [CrossRef]
- Raya, A.; Cuervo, A.M.; Macián, F.; Romero, F.J.; Romá, J. Nerve conduction velocity decrease and synaptic transmission alterations in caffeine-treated rats. Neurotoxicol. Teratol. 1994, 16, 11–15. [Google Scholar] [CrossRef]
- Chéreau, R.; Saraceno, G.E.; Angibaud, J.; Cattaert, D.; Nägerl, U.V. Superresolution imaging reveals activity-dependent plasticity of axon morphology linked to changes in action potential conduction velocity. Proc. Natl. Acad. Sci. USA 2017, 114, 1401–1406. [Google Scholar] [CrossRef] [Green Version]
- Barak, B.; Zhang, Z.; Liu, Y.; Nir, A.; Trangle, S.S.; Ennis, M.; Levandowski, K.M.; Wang, D.; Quast, K.; Boulting, G.L.; et al. Neuronal deletion of Gtf2i, associated with Williams syndrome, causes behavioral and myelin alterations rescuable by a remyelinating drug. Nat. Neurosci. 2019, 22, 700–708. [Google Scholar] [CrossRef]
- Gibson, E.M.; Purger, D.; Mount, C.W.; Goldstein, A.K.; Lin, G.L.; Wood, L.S.; Inema, I.; Miller, S.E.; Bieri, G.; Zuchero, J.B.; et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science 2014, 344, 1252304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Dietz, K.; DeLoyht, J.M.; Pedre, X.; Kelkar, D.; Kaur, J.; Vialou, V.; Lobo, M.K.; Dietz, D.M.; Nestler, E.J.; et al. Impaired adult myelination in the prefrontal cortex of socially isolated mice. Nat. Neurosci. 2012, 15, 1621–1623. [Google Scholar] [CrossRef] [Green Version]
- Makinodan, M.; Rosen, K.M.; Ito, S.; Corfas, G. A critical period for social experience-dependent oligodendrocyte maturation and myelination. Science 2012, 337, 1357–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenzie, I.A.; Ohayon, D.; Li, H.; de Faria, J.P.; Emery, B.; Tohyama, K.; Richardson, W.D. Motor skill learning requires active central myelination. Science 2014, 346, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Pesaresi, M.; Soon-Shiong, R.; French, L.; Kaplan, D.R.; Miller, F.D.; Paus, T. Axon diameter and axonal transport: In vivo and in vitro effects of androgens. Neuroimage 2015, 115, 191–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stedehouder, J.; Brizee, D.; Shpak, G.; Kushner, S.A. Activity-dependent myelination of parvalbumin interneurons mediated by axonal morphological plasticity. J. Neurosci. 2018, 38, 3631–3642. [Google Scholar] [CrossRef] [PubMed]
- Reess, T.J.; Rus, O.G.; Schmidt, R.; de Reus, M.A.; Zaudig, M.; Wagner, G.; Zimmer, C.; van den Heuvel, M.P.; Koch, K. Connectomics-based structural network alterations in obsessive-compulsive disorder. Transl. Psychiatry 2016, 6, e882. [Google Scholar] [CrossRef] [Green Version]
- Gan, J.; Zhong, M.; Fan, J.; Liu, W.; Niu, C.; Cai, S.; Zou, L.; Wang, Y.; Wang, Y.; Tan, C.; et al. Abnormal white matter structural connectivity in adults with obsessive-compulsive disorder. Transl. Psychiatry 2017, 7, e1062. [Google Scholar] [CrossRef]
- Zaimi, A.; Wabartha, M.; Herman, V.; Antonsanti, P.-L.; Perone, C.S.; Cohen-Adad, J. AxonDeepSeg: Automatic axon and myelin segmentation from microscopy data using convolutional neural networks. Sci. Rep. 2018, 8, 3816. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.W.; Harris, J.A.; Ng, L.; Winslow, B.; Cain, N.; Mihalas, S.; Wang, Q.; Lau, C.; Kuan, L.; Henry, A.M.; et al. A mesoscale connectome of the mouse brain. Nature 2014, 508, 207–214. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 2001; ISBN 978-0-12-547636-2. [Google Scholar]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.; Poggiali, D.; Whitaker, K.; Marshall, T.R.; Kievit, R.A. Raincloud plots: A multi-platform tool for robust data visualization. Wellcome Open Res. 2019, 4, 63. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.; Tumkaya, T.; Aryal, S.; Choi, H.; Claridge-Chang, A. Moving beyond p values: Data analysis with estimation graphics. Nat. Methods 2019, 16, 565–566. [Google Scholar] [CrossRef] [PubMed]
- Patel, E.; Kushwaha, D.S. Clustering cloud workloads: K-means vs gaussian mixture model. Procedia Comput. Sci. 2020, 171, 158–167. [Google Scholar] [CrossRef]
- Voyvodic, J.T. Target size regulates calibre and myelination of sympathetic axons. Nature 1989, 342, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Tomassy, G.S.; Berger, D.R.; Chen, H.-H.; Kasthuri, N.; Hayworth, K.J.; Vercelli, A.; Seung, H.S.; Lichtman, J.W.; Arlotta, P. Distinct profiles of myelin distribution along single axons of pyramidal neurons in the neocortex. Science 2014, 344, 319–324. [Google Scholar] [CrossRef] [Green Version]
- Boron, W.F.; Boulpaep, E.L. Medical Physiology, 3rd ed.; Elsevier: Philadelphia, PA, USA, 2016; ISBN 978-1-4557-4377-3. [Google Scholar]
- Rushton, W.A.H. A theory of the effects of fibre size in medullated nerve. J. Physiol. 1951, 115, 101–122. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, G.M.; Caminiti, R. Axon diameter relates to synaptic bouton size: Structural properties define computationally different types of cortical connections in primates. Brain Struct. Funct. 2017, 222, 1169–1177. [Google Scholar] [CrossRef]
- Maas, D.A.; Eijsink, V.D.; Spoelder, M.; van Hulten, J.A.; De Weerd, P.; Homberg, J.R.; Vallès, A.; Nait-Oumesmar, B.; Martens, G.J.M. Interneuron hypomyelination is associated with cognitive inflexibility in a rat model of schizophrenia. Nat. Commun. 2020, 11, 2329. [Google Scholar] [CrossRef]
- Wegiel, J.; Kaczmarski, W.; Flory, M.; Martinez-Cerdeno, V.; Wisniewski, T.; Nowicki, K.; Kuchna, I.; Wegiel, J. Deficit of corpus callosum axons, reduced axon diameter and decreased area are markers of abnormal development of interhemispheric connections in autistic subjects. Acta Neuropathol. Commun. 2018, 6, 143. [Google Scholar] [CrossRef]
- Hadjas, L.C.; Schartner, M.M.; Cand, J.; Creed, M.C.; Pascoli, V.; Lüscher, C.; Simmler, L.D. Projection-specific deficits in synaptic transmission in adult Sapap3-knockout mice. Neuropsychopharmacology 2020, 45, 2020–2029. [Google Scholar] [CrossRef]
- Remijnse, P.L.; Nielen, M.M.A.; van Balkom, A.J.L.M.; Cath, D.C.; van Oppen, P.; Uylings, H.B.M.; Veltman, D.J. Reduced orbitofrontal-striatal activity on a reversal learning task in obsessive-compulsive disorder. Arch. Gen. Psychiatry 2006, 63, 1225–1236. [Google Scholar] [CrossRef]
- Jung, W.H.; Yücel, M.; Yun, J.-Y.; Yoon, Y.B.; Cho, K.I.K.; Parkes, L.; Kim, S.N.; Kwon, J.S. Altered functional network architecture in orbitofronto-striato-thalamic circuit of unmedicated patients with obsessive-compulsive disorder. Hum. Brain Mapp. 2017, 38, 109–119. [Google Scholar] [CrossRef]
- Gillan, C.M.; Robbins, T.W.; Sahakian, B.J.; van den Heuvel, O.A.; van Wingen, G. The role of habit in compulsivity. Eur. Neuropsychopharmacol. 2016, 26, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Hadjas, L.C.; Lüscher, C.; Simmler, L.D. Aberrant habit formation in the Sapap3-knockout mouse model of obsessive-compulsive disorder. Sci. Rep. 2019, 9, 12061. [Google Scholar] [CrossRef] [PubMed]
- Waxman, S.G.; Bennett, M.V.L. Relative conduction velocities of small myelinated and non-myelinated fibres in the central nervous system. Nat. New Biol. 1972, 238, 217–219. [Google Scholar] [CrossRef] [PubMed]
- Weyand, T.G.; Gafka, A.C. Corticostriatal and corticotectal neurons in area 6 of the cat during fixation and eye movements. Vis. Neurosci. 1998, 15, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Filipović, M. Characterisation of Inputs and Outputs of Striatal Medium Spiny Neurons in Health and Disease. Ph.D. Thesis, University of Freiburg, Freiburg, Germany, 2019. [Google Scholar]
- Whitlock, D.G.; Nauta, W.J. Subcortical projections from the temporal neocortex in macaca mulatta. J. Comp. Neurol. 1956, 106, 183–212. [Google Scholar] [CrossRef]
- Bauswein, E.; Fromm, C.; Preuss, A. Corticostriatal cells in comparison with pyramidal tract neurons: Contrasting properties in the behaving monkey. Brain Res. 1989, 493, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Bishop, G.A.; Chang, H.T.; Kitai, S.T. Morphological and physiological properties of neostriatal neurons: An intracellular horseradish peroxidase study in the rat. Neuroscience 1982, 7, 179–191. [Google Scholar] [CrossRef]
- Kita, H.; Kosaka, T.; Heizmann, C.W. Parvalbumin-immunoreactive neurons in the rat neostriatum: A light and electron microscopic study. Brain Res. 1990, 536, 1–15. [Google Scholar] [CrossRef]
- Phelps, P.E.; Houser, C.R.; Vaughn, J.E. Immunocytochemical localization of choline acetyltransferase within the rat neostriatum: A correlated light and electron microscopic study of cholinergic neurons and synapses. J. Comp. Neurol. 1985, 238, 286–307. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K. Poor and protracted myelination as a contributory factor to neurodegenerative disorders. Neurobiol. Aging 2004, 25, 19–23. [Google Scholar] [CrossRef]
- Orimo, S.; Uchihara, T.; Kanazawa, T.; Itoh, Y.; Wakabayashi, K.; Kakita, A.; Takahashi, H. Unmyelinated axons are more vulnerable to degeneration than myelinated axons of the cardiac nerve in Parkinson’s disease. Neuropathol. Appl. Neurobiol. 2011, 37, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Rockland, K.S. Axon collaterals and brain states. Front. Syst. Neurosci. 2018, 12, 32. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.A.C.; Roth, S.; Rusch, E.S. Superficial layer pyramidal cells communicate heterogeneously between multiple functional domains of cat primary visual cortex. Nat. Commun. 2014, 5, 5252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koestinger, G.; Martin, K.A.C.; Roth, S.; Rusch, E.S. Synaptic connections formed by patchy projections of pyramidal cells in the superficial layers of cat visual cortex. Brain Struct. Funct. 2017, 222, 3025–3042. [Google Scholar] [CrossRef] [Green Version]
- Kalanithi, P.S.A.; Zheng, W.; Kataoka, Y.; DiFiglia, M.; Grantz, H.; Saper, C.B.; Schwartz, M.L.; Leckman, J.F.; Vaccarino, F.M. Altered parvalbumin-positive neuron distribution in basal ganglia of individuals with Tourette syndrome. Proc. Natl. Acad. Sci. USA 2005, 102, 13307–13312. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, Y.; Kalanithi, P.S.A.; Grantz, H.; Schwartz, M.L.; Saper, C.; Leckman, J.F.; Vaccarino, F.M. Decreased number of parvalbumin and cholinergic interneurons in the striatum of individuals with Tourette syndrome. J. Comp. Neurol. 2010, 518, 277–291. [Google Scholar] [CrossRef]
- Reig, R.; Silberberg, G. Multisensory integration in the mouse striatum. Neuron 2014, 83, 1200–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koós, T.; Tepper, J.M. Inhibitory control of neostriatal projection neurons by GABAergic interneurons. Nat. Neurosci. 1999, 2, 467–472. [Google Scholar] [CrossRef]
- Lauber, E.; Filice, F.; Schwaller, B. Dysregulation of parvalbumin expression in the Cntnap2−/− mouse model of autism spectrum disorder. Front. Mol. Neurosci. 2018, 11, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burguière, E.; Monteiro, P.; Mallet, L.; Feng, G.; Graybiel, A.M. Striatal circuits, habits, and implications for obsessive—Compulsive disorder. Curr. Opin. Neurobiol. 2015, 30, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Mallet, N.; Leblois, A.; Maurice, N.; Beurrier, C. Striatal cholinergic interneurons: How to elucidate their function in health and disease. Front. Pharmacol. 2019, 10, 1488. [Google Scholar] [CrossRef] [Green Version]
- Martos, Y.V.; Braz, B.Y.; Beccaria, J.P.; Murer, M.G.; Belforte, J.E. Compulsive social behavior emerges after selective ablation of striatal cholinergic interneurons. J. Neurosci. 2017, 37, 2849–2858. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lousada, E.; Boudreau, M.; Cohen-Adad, J.; Nait Oumesmar, B.; Burguière, E.; Schreiweis, C. Reduced Axon Calibre in the Associative Striatum of the Sapap3 Knockout Mouse. Brain Sci. 2021, 11, 1353. https://doi.org/10.3390/brainsci11101353
Lousada E, Boudreau M, Cohen-Adad J, Nait Oumesmar B, Burguière E, Schreiweis C. Reduced Axon Calibre in the Associative Striatum of the Sapap3 Knockout Mouse. Brain Sciences. 2021; 11(10):1353. https://doi.org/10.3390/brainsci11101353
Chicago/Turabian StyleLousada, Eliana, Mathieu Boudreau, Julien Cohen-Adad, Brahim Nait Oumesmar, Eric Burguière, and Christiane Schreiweis. 2021. "Reduced Axon Calibre in the Associative Striatum of the Sapap3 Knockout Mouse" Brain Sciences 11, no. 10: 1353. https://doi.org/10.3390/brainsci11101353
APA StyleLousada, E., Boudreau, M., Cohen-Adad, J., Nait Oumesmar, B., Burguière, E., & Schreiweis, C. (2021). Reduced Axon Calibre in the Associative Striatum of the Sapap3 Knockout Mouse. Brain Sciences, 11(10), 1353. https://doi.org/10.3390/brainsci11101353