A Review on Ultrasonic Neuromodulation of the Peripheral Nervous System: Enhanced or Suppressed Activities?


Abstract
Featured Application
Abstract
1. Introduction
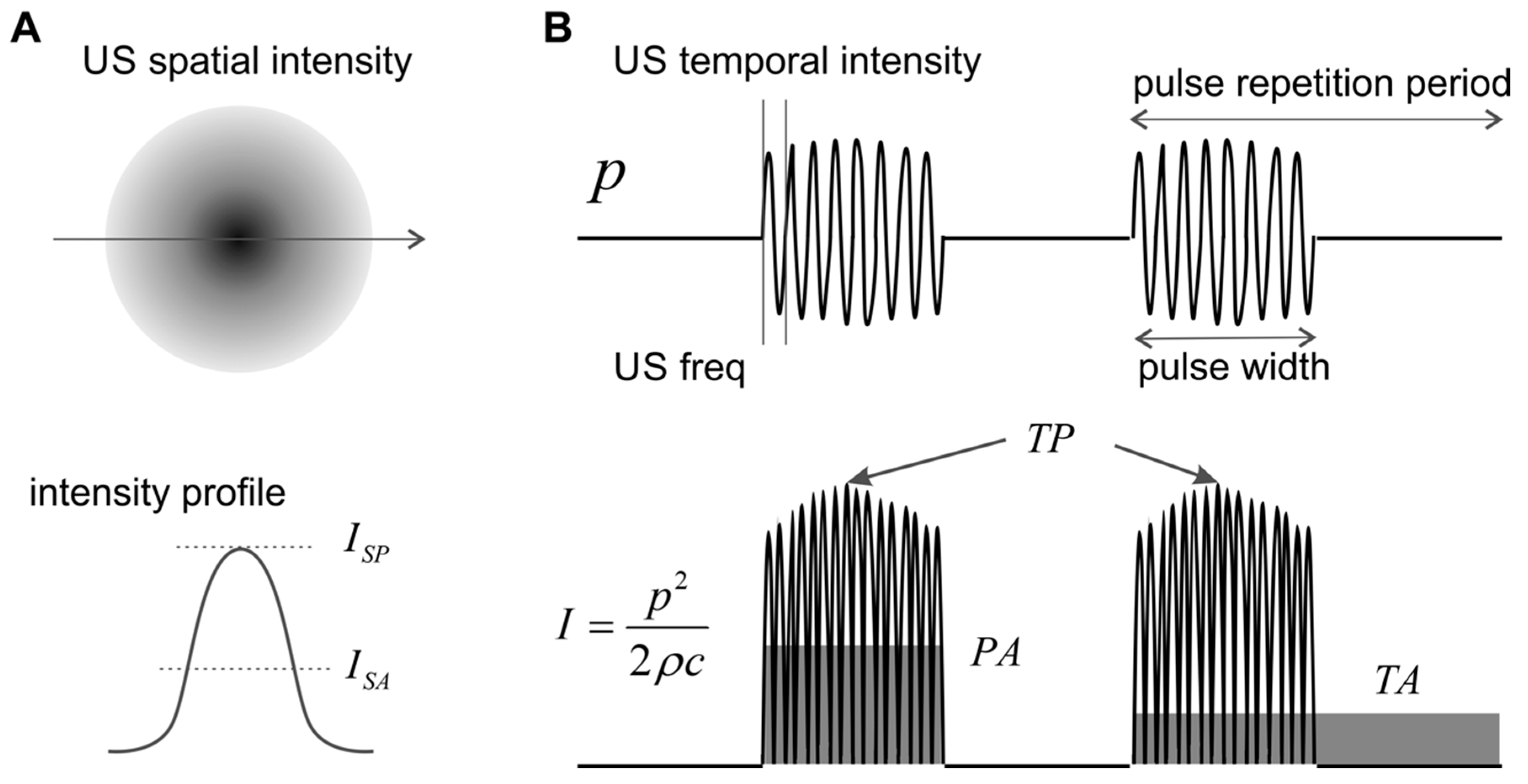
2. Peripheral Ultrasonic Neuromodulation—Technical Specifications
2.1. US Frequency
2.2. Pulse Repetition Frequency
2.3. Duty Cycle
2.4. Intensity
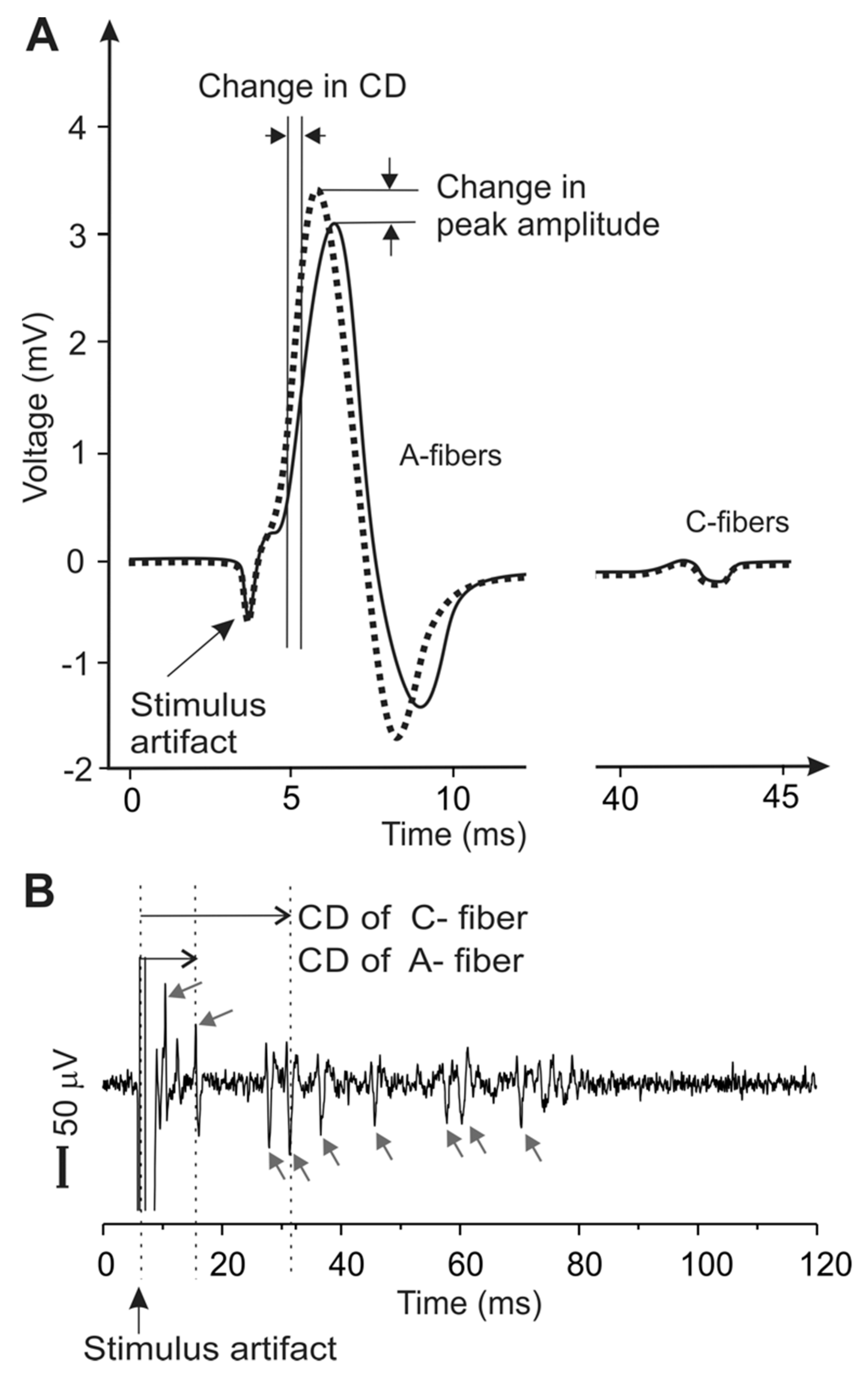
2.5. Metrics for Peripheral Neural Activities
2.6. Biological Preparations to Assess Peripheral US Neuromodulation
3. Ultrasonic Neuromodulation of the Peripheral Nervous System
4. Mechanisms of Peripheral US Neuromodulation
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dubner, R.; Gold, M. The neurobiology of pain. Proc. Nat. Acad. Sci. USA 1999, 96, 7627–7630. [Google Scholar] [CrossRef] [PubMed]
- Woolf, C.J. Overcoming obstacles to developing new analgesics. Nat. Med. 2010, 16, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Seeley, W.W.; Crawford, R.K.; Zhou, J.; Miller, B.L.; Greicius, M.D. Neurodegenerative diseases target large-scale human brain networks. Neuron 2009, 62, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Tracey, I.; Dickenson, A. SnapShot: Pain perception. Cell 2012, 148, 1308. [Google Scholar] [CrossRef]
- De Ridder, D.; Perera, S.; Vanneste, S. State of the art: Novel applications for cortical stimulation. Neuromodul. J. Int. Neuromod. Soci. 2017, 20, 206–214. [Google Scholar] [CrossRef]
- Hunter, C.; Dave, N.; Diwan, S.; Deer, T. Neuromodulation of pelvic visceral pain: Review of the literature and case series of potential novel targets for treatment. Pain Pract. 2013, 13, 3–17. [Google Scholar] [CrossRef]
- Goroszeniuk, T.; Pang, D. Peripheral neuromodulation: A review. Curr. Pain Headache Rep. 2014, 18, 412. [Google Scholar] [CrossRef]
- Zhang, T.C.; Janik, J.J.; Grill, W.M. Mechanisms and models of spinal cord stimulation for the treatment of neuropathic pain. Brain Res. 2014, 1569, 19–31. [Google Scholar] [CrossRef]
- Tyler, W.J.; Lani, S.W.; Hwang, G.M. Ultrasonic modulation of neural circuit activity. Curr. Opin. Neurobiol. 2018, 50, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Chervyakov, A.V.; Chernyavsky, A.Y.; Sinitsyn, D.O.; Piradov, M.A. Possible mechanisms underlying the therapeutic effects of transcranial magnetic stimulation. Front. Hum. Neurosci. 2015, 9, 303. [Google Scholar] [CrossRef]
- Klein, M.M.; Treister, R.; Raij, T.; Pascual-Leone, A.; Park, L.; Nurmikko, T.; Lenz, F.; Lefaucheur, J.P.; Lang, M.; Hallett, M.; et al. Transcranial magnetic stimulation of the brain: Guidelines for pain treatment research. Pain 2015, 156, 1601–1614. [Google Scholar] [CrossRef] [PubMed]
- Chernov, M.; Roe, A.W. Infrared neural stimulation: A new stimulation tool for central nervous system applications. Neurophotonics 2014, 1, 011011. [Google Scholar] [CrossRef]
- Kobus, T.; Vykhodtseva, N.; Pilatou, M.; Zhang, Y.; McDannold, N. Safety validation of repeated blood-brain barrier disruption using focused ultrasound. Ultrasound Med. Biol. 2016, 42, 481–492. [Google Scholar] [CrossRef]
- Kamimura, H.A.; Wang, S.; Chen, H.; Wang, Q.; Aurup, C.; Acosta, C.; Carneiro, A.A.; Konofagou, E.E. Focused ultrasound neuromodulation of cortical and subcortical brain structures using 1.9 MHz. Med. Phys. 2016, 43, 5730. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Park, M.Y.; Lee, S.D.; Lee, W.; Chiu, A.; Yoo, S.S. Suppression of EEG visual-evoked potentials in rats through neuromodulatory focused ultrasound. Neuroreport 2015, 26, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.C.; Liu, H.L.; Lai, H.Y.; Lin, C.Y.; Tsai, H.C.; Pei, Y.C. Neuromodulation accompanying focused ultrasound-induced blood-brain barrier opening. Sci. Rep. 2015, 5, 15477. [Google Scholar] [CrossRef] [PubMed]
- Mehic, E.; Xu, J.M.; Caler, C.J.; Coulson, N.K.; Moritz, C.T.; Mourad, P.D. Increased anatomical specificity of neuromodulation via modulated focused ultrasound. PLoS ONE 2014, 9, e86939. [Google Scholar] [CrossRef]
- Kim, H.; Chiu, A.; Lee, S.D.; Fischer, K.; Yoo, S.S. Focused ultrasound-mediated non-invasive brain stimulation: Examination of sonication parameters. Brain Stimul. 2014, 7, 748–756. [Google Scholar] [CrossRef]
- Kim, H.; Lee, S.D.; Chiu, A.; Yoo, S.S.; Park, S. Estimation of the spatial profile of neuromodulation and the temporal latency in motor responses induced by focused ultrasound brain stimulation. Neuroreport 2014, 25, 475–479. [Google Scholar] [CrossRef]
- King, R.L.; Brown, J.R.; Newsome, W.T.; Pauly, K.B. Effective parameters for ultrasound-induced in vivo neurostimulation. Ultrasound Med. Biol. 2013, 39, 312–331. [Google Scholar] [CrossRef]
- Yoo, S.S.; Kim, H.; Min, B.K.; Franck, E.; Park, S. Transcranial focused ultrasound to the thalamus alters anesthesia time in rats. Neuroreport 2011, 22, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Tufail, Y.; Matyushov, A.; Baldwin, N.; Tauchmann, M.L.; Georges, J.; Yoshihiro, A.; Tillery, S.I.; Tyler, W.J. Transcranial pulsed ultrasound stimulates intact brain circuits. Neuron 2010, 66, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Hynynen, K.; McDannold, N.; Sheikov, N.A.; Jolesz, F.A.; Vykhodtseva, N. Local and reversible blood-brain barrier disruption by noninvasive focused ultrasound at frequencies suitable for trans-skull sonications. Neuroimage 2005, 24, 12–20. [Google Scholar] [CrossRef]
- Wattiez, N.; Constans, C.; Deffieux, T.; Daye, P.M.; Tanter, M.; Aubry, J.F.; Pouget, P. Transcranial ultrasonic stimulation modulates single-neuron discharge in macaques performing an antisaccade task. Brain Stimul. 2017, 10, 1024–1031. [Google Scholar] [CrossRef]
- Verhagen, L.; Gallea, C.; Folloni, D.; Constans, C.; Jensen, D.E.; Ahnine, H.; Roumazeilles, L.; Santin, M.; Ahmed, B.; Lehericy, S.; et al. Offline impact of transcranial focused ultrasound on cortical activation in primates. eLife 2019, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Fishman, P.S.; Frenkel, V. Focused ultrasound: An emerging therapeutic modality for neurologic disease. Neurotherapeutics 2017, 14, 393–404. [Google Scholar] [CrossRef]
- Bachtold, M.R.; Rinaldi, P.C.; Jones, J.P.; Reines, F.; Price, L.R. Focused ultrasound modifications of neural circuit activity in a mammalian brain. Ultrasound Med. Biol. 1998, 24, 557–565. [Google Scholar] [CrossRef]
- Tyler, W.J.; Tufail, Y.; Finsterwald, M.; Tauchmann, M.L.; Olson, E.J.; Majestic, C. Remote excitation of neuronal circuits using low-intensity, low-frequency ultrasound. PLoS ONE 2008, 3, e3511. [Google Scholar] [CrossRef]
- Sato, T.; Shapiro, M.G.; Tsao, D.Y. Ultrasonic neuromodulation causes widespread cortical activation via an indirect auditory mechanism. Neuron 2018, 98, 1031–1041.e5. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Hamilton, I.I.M.; Offutt, S.J.; Gloeckner, C.D.; Li, T.; Kim, Y.; Legon, W.; Alford, J.K.; Lim, H.H. Ultrasound produces extensive brain activation via a cochlear pathway. Neuron 2018, 98, 1020–1030.e4. [Google Scholar] [CrossRef]
- Menz, M.D.; Oralkan, Ö.; Khuri-Yakub, P.T.; Baccus, S.A. Precise neural stimulation in the retina using focused ultrasound. J. Neurosci. 2013, 33, 4550–4560. [Google Scholar] [CrossRef] [PubMed]
- Foreman, R.D.; Linderoth, B. Neural mechanisms of spinal cord stimulation. Int. Rev. Neurobiol. 2012, 107, 87–119. [Google Scholar] [PubMed]
- Frahm, K.S.; Hennings, K.; Vera-Portocarrero, L.; Wacnik, P.W.; Morch, C.D. Nerve fiber activation during peripheral nerve field stimulation: Importance of electrode orientation and estimation of area of paresthesia. Neuromodul. J. Int. Neuromodul. Soc. 2016, 19, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Morch, C.D.; Nguyen, G.P.; Wacnik, P.W.; Andersen, O.K. Mathematical model of nerve fiber activation during low back peripheral nerve field stimulation: Analysis of electrode implant depth. Neuromodul. J. Int. Neuromodul. Soc. 2014, 17, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Chakravarthy, K.; Nava, A.; Christo, P.J.; Williams, K. Review of recent advances in peripheral nerve stimulation (PNS). Curr. Pain Headache Rep. 2016, 20, 60. [Google Scholar] [CrossRef] [PubMed]
- Krames, E.S. The dorsal root ganglion in chronic pain and as a target for neuromodulation: A review. Neuromodul. J. Int. Neuromodul. Soc. 2015, 18, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Liem, L.; van Dongen, E.; Huygen, F.J.; Staats, P.; Kramer, J. The dorsal root ganglion as a therapeutic target for chronic pain. Reg. Anesth. Pain Med. 2016, 41, 511–519. [Google Scholar] [CrossRef]
- Omer, N.; Steve, K.; Shy, S. Ultrasonic neuromodulation. J. Neural Eng. 2016, 13, 031003. [Google Scholar]
- Patel, P.R.; Luk, A.; Durrani, A.; Dromi, S.; Cuesta, J.; Angstadt, M.; Dreher, M.R.; Wood, B.J.; Frenkel, V. In vitro and in vivo evaluations of increased effective beam width for heat deposition using a split focus high intensity ultrasound (HIFU) transducer. Int. J. Hyperth. 2008, 24, 537–549. [Google Scholar] [CrossRef]
- Naor, O.; Hertzberg, Y.; Zemel, E.; Kimmel, E.; Shoham, S. Towards multifocal ultrasonic neural stimulation II: Design considerations for an acoustic retinal prosthesis. J. Neural Eng. 2012, 9, 026006. [Google Scholar] [CrossRef]
- Baker, M.L.; Dalrymple, G.V. Biological effects of diagnostic ultrasound: A review. Radiology 1978, 126, 479–483. [Google Scholar] [CrossRef]
- Casella, D.P.; Dudley, A.G.; Clayton, D.B.; Pope, J.C., IV; Tanaka, S.T.; Thomas, J.; Adams, M.C.; Brock, J.W., III; Caskey, C.F. Modulation of the rat micturition reflex with transcutaneous ultrasound. Neurourol. Urodyn. 2017, 36, 1996–2002. [Google Scholar] [CrossRef]
- Ilham, S.J.; Chen, L.; Guo, T.; Emadi, S.; Hoshino, K.; Feng, B. In vitro single-unit recordings reveal increased peripheral nerve conduction velocity by focused pulsed ultrasound. Biomed. Phys. Eng. Express. 2018, 4, 045004. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.F. High intensity focused ultrasound in clinical tumor ablation. World J. Clin. Oncol. 2011, 2, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Xu, G. Damage effect of high-intensity focused ultrasound on breast cancer tissues and their vascularities. World J. Surg. Oncol. 2016, 14, 153. [Google Scholar] [CrossRef] [PubMed]
- Copelan, A.; Hartman, J.; Chehab, M.; Venkatesan, A.M. High-intensity focused ultrasound: Current status for image-guided therapy. Semin. Interv. Radiol. 2015, 32, 398–415. [Google Scholar] [CrossRef] [PubMed]
- Juan, E.J.; Gonzalez, R.; Albors, G.; Ward, M.P.; Irazoqui, P. Vagus nerve modulation using focused pulsed ultrasound: Potential applications and preliminary observations. Int. J. Imaging Syst. Technol. 2014, 24, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ilham, S.J.; Guo, T.; Emadi, S.; Feng, B. In vitro multichannel single-unit recordings of action potentials from mouse sciatic nerve. Biomed. Phys. Eng. Express. 2017, 3, 045020. [Google Scholar] [CrossRef]
- Lee, Y.F.; Lin, C.C.; Cheng, J.S.; Chen, G.S. High-intensity focused ultrasound attenuates neural responses of sciatic nerves isolated from normal or neuropathic rats. Ultrasound Med. Biol. 2015, 41, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Colucci, V.; Strichartz, G.; Jolesz, F.; Vykhodtseva, N.; Hynynen, K. Focused ultrasound effects on nerve action potential in vitro. Ultrasound Med. Biol. 2009, 35, 1737–1747. [Google Scholar] [CrossRef]
- Tsui, P.H.; Wang, S.H.; Huang, C.C. In vitro effects of ultrasound with different energies on the conduction properties of neural tissue. Ultrasonics 2005, 43, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Mihran, R.T.; Barnes, F.S.; Wachtel, H. Temporally-specific modification of myelinated axon excitability in vitro following a single ultrasound pulse. Ultrasound Med. Biol. 1990, 16, 297–309. [Google Scholar] [CrossRef]
- Yoo, S.S.; Lee, W.; Kim, H. Pulsed application of focused ultrasound to the LI4 elicits deqi sensations: Pilot study. Complement. Ther. Med. 2014, 22, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.L.; Little, J.W.; Vaezy, S. Image-guided high-intensity focused ultrasound for conduction block of peripheral nerves. Ann. Biomed. Eng. 2007, 35, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Li, B.Y.; Qiao, G.F.; Feng, B.; Zhao, R.B.; Lu, Y.J.; Schild, J.H. Electrophysiological and neuroanatomical evidence of sexual dimorphism in aortic baroreceptor and vagal afferents in rat. Am. J. Physiol. Regul. Integr. Comp. Phys. 2008, 295, R1301–R1310. [Google Scholar] [CrossRef] [PubMed]
- Li, B.Y.; Feng, B.; Tsu, H.Y.; Schild, J.H. Unmyelinated visceral afferents exhibit frequency dependent action potential broadening while myelinated visceral afferents do not. Neurosci. Lett. 2007, 421, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Gebhart, G.F. In vitro functional characterization of mouse colorectal afferent endings. J. Vis. Exp. JoVE 2015, 95, 52310. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Gebhart, G.F. Characterization of silent afferents in the pelvic and splanchnic innervations of the mouse colorectum. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G170–G180. [Google Scholar] [CrossRef]
- Feng, B.; Zhu, Y.; La, J.H.; Wills, Z.P.; Gebhart, G.F. Experimental and computational evidence for an essential role of NaV1.6 in spike initiation at stretch-sensitive colorectal afferent endings. J. Neurophysiol. 2015, 113, 2618–2634. [Google Scholar] [CrossRef] [PubMed]
- Hoffer, J.A.; Loeb, G.E.; Pratt, C.A. Single unit conduction velocities from averaged nerve cuff electrode records in freely moving cats. J. Neurosci. Methods 1981, 4, 211–225. [Google Scholar] [CrossRef]
- Sharkey, K.A.; Cervero, F. An in vitro method for recording single unit afferent activity from mesenteric nerves innervating isolated segments of rat ileum. J. Neurosci. Methods 1986, 16, 149–156. [Google Scholar] [CrossRef]
- Schalow, G.; Lang, G. Recording of single unit potentials in human spinal nerve roots: A new diagnostic tool. Acta Neurochir. 1987, 86, 25–29. [Google Scholar] [CrossRef]
- Guiloff, R.J.; Modarres-Sadeghi, H. Preferential generation of recurrent responses by groups of motor neurons in man. Conventional and single unit F wave studies. Brain 1991, 114, 1771–1801. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Ekedahl, R.; Hallin, R.G. Consistency of unitary shapes in dual lead recordings from myelinated fibres in human peripheral nerves: Evidence for extracellular single-unit recordings in microneurography. Exp. Brain Res. 1998, 120, 470–478. [Google Scholar] [CrossRef]
- Kagitani, F.; Uchida, S.; Hotta, H.; Aikawa, Y. Manual acupuncture needle stimulation of the rat hindlimb activates groups I, II, III and IV single afferent nerve fibers in the dorsal spinal roots. Jpn. J. Physiol. 2005, 55, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, M.; Schmidt, R. Microneurographic single-unit recordings to assess receptive properties of afferent human C-fibers. Neurosci. Lett. 2010, 470, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Gaunt, R.A.; Bruns, T.M.; Crammond, D.J.; Tomycz, N.D.; Moossy, J.J.; Weber, D.J. Single- and multi-unit activity recorded from the surface of the dorsal root ganglia with non-penetrating electrode arrays. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2011, 2011, 6713–6716. [Google Scholar] [PubMed]
- Srinivasan, A.; Tipton, J.; Tahilramani, M.; Kharbouch, A.; Gaupp, E.; Song, C.; Venkataraman, P.; Falcone, J.; Lacour, S.P.; Stanley, G.B.; et al. A regenerative microchannel device for recording multiple single-unit action potentials in awake, ambulatory animals. Eur. J. Neurosci. 2016, 43, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Florez-Paz, D.M.; Tong, C.-K.; Hoffman, B.U.; Lee, S.A.; Konofagou, E.E.; Lumpkin, E.A. Focused ultrasound evoked responses in dorsal root ganglion neurons (DRG) and HEK293 cells. Biophys. J. 2018, 114, 673a. [Google Scholar] [CrossRef]
- Joe, E.-H.; Angelides, K. Clustering of voltage-dependent sodium channels on axons depends on Schwann cell contact. Nature 1992, 356, 333–335. [Google Scholar] [CrossRef]
- Harvey, E.N. The effect of high frequency sound waves on heart muscle and other irritable tissues. Am. J. Physiol. 1929, 91, 284–290. [Google Scholar] [CrossRef]
- Gavrilov, L.R.; Gersuni, G.V.; Ilyinski, O.B.; Tsirulnikov, E.M.; Shchekanov, E.E. A study of reception with the use of focused ultrasound. I. Effects on the skin and deep receptor structures in man. Brain Res. 1977, 135, 265–277. [Google Scholar] [CrossRef]
- Ab Ithel Davies, I.; Gavrilov, L.R.; Tsirulnikov, E.M. Application of focused ultrasound for research on pain. Pain 1996, 67, 17–27. [Google Scholar] [CrossRef]
- Dalecki, D.; Child, S.Z.; Raeman, C.H.; Carstensen, E.L. Tactile perception of ultrasound. J. Acoust. Soc. Am. 1995, 97, 3165–3170. [Google Scholar] [CrossRef]
- Tsirulnikov, E.M.; Vartanyan, I.A.; Gersuni, G.V.; Rosenblyum, A.S.; Pudov, V.I.; Gavrilov, L.R. Use of amplitude-modulated focused ultrasound for diagnosis of hearing disorders. Ultrasound Med. Biol. 1988, 14, 277–285. [Google Scholar] [CrossRef]
- Foster, K.R.; Wiederhold, M.L. Auditory responses in cats produced by pulsed ultrasound. J. Acoust. Soc. Am. 1978, 63, 1199–1205. [Google Scholar] [CrossRef]
- Gavrilov, L.R.; Gersuni, G.V.; Ilyinsky, O.B.; Tsirulnikov, E.M.; Shchekanov, E.E. A study of reception with the use of focused ultrasound. II. Effects on the animal receptor structures. Brain Res. 1977, 135, 279–285. [Google Scholar] [CrossRef]
- Saffari, N.; Wright, C.J.; Rothwell, J. Ultrasound neuro-stimulation effects of peripheral axons in-vitro. J. Acoust. Soc. Am. 2017, 142, 2668. [Google Scholar] [CrossRef]
- Foley, J.L.; Little, J.W.; Vaezy, S. Effects of high-intensity focused ultrasound on nerve conduction. Muscle Nerve 2008, 37, 241–250. [Google Scholar] [CrossRef]
- Lee, Y.F.; Lin, C.C.; Cheng, J.S.; Chen, G.S. Nerve conduction block in diabetic rats using high-intensity focused ultrasound for analgesic applications. BJA Br. J. Anaesth. 2015, 114, 840–846. [Google Scholar] [CrossRef]
- Moore, J.H.; Gieck, J.H.; Saliba, E.N.; Perrin, D.H.; Ball, D.W.; McCue, F.C. The biophysical effects of ultrasound on median nerve distal latencies. Electromyogr. Clin. Neurophysiol. 2000, 40, 169–180. [Google Scholar]
- Deer, T.R.; Grigsby, E.; Weiner, R.L.; Wilcosky, B.; Kramer, J.M. A prospective study of dorsal root ganglion stimulation for the relief of chronic pain. Neuromodul. J. Int. Neuromodul. Soc. 2013, 16, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Liem, L.; Russo, M.; Huygen, F.J.; Van Buyten, J.P.; Smet, I.; Verrills, P.; Cousins, M.; Brooker, C.; Levy, R.; Deer, T.; et al. A multicenter, prospective trial to assess the safety and performance of the spinal modulation dorsal root ganglion neurostimulator system in the treatment of chronic pain. Neuromodul. J. Int. Neuromodul. Soc. 2013, 16, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Van Buyten, J.P.; Smet, I.; Liem, L.; Russo, M.; Huygen, F. Stimulation of dorsal root Ganglia for the management of complex regional pain syndrome: A prospective case series. Pain Pract. 2015, 15, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Wahab, R.A.; Choi, M.; Liu, Y.; Krauthamer, V.; Zderic, V.; Myers, M.R. Mechanical bioeffects of pulsed high intensity focused ultrasound on a simple neural model. Med. Phys. 2012, 39, 4274–4283. [Google Scholar] [CrossRef]
- Young, R.R.; Henneman, E. Functional effects of focused ultrasound on mammalian nerves. Science 1961, 134, 1521–1522. [Google Scholar] [CrossRef]
- Young, R.R.; Henneman, E. Reversible block of nerve conduction by ultrasound: Ultrasonic blocking of nerve fibers. Arch. Neurol. 1961, 4, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Johnston, K.; Tapia-Siles, C.; Gerold, B.; Postema, M.; Cochran, S.; Cuschieri, A.; Prentice, P. Periodic shock-emission from acoustically driven cavitation clouds: A source of the subharmonic signal. Ultrasonics 2014, 54, 2151–2158. [Google Scholar] [CrossRef]
- Xu, D.; Pollock, M. Experimental nerve thermal injury. Brain J. Neurol. 1994, 117, 375–384. [Google Scholar] [CrossRef]
- Ernberg, L.A.; Adler, R.S.; Lane, J. Ultrasound in the detection and treatment of a painful stump neuroma. Skelet. Radiol. 2003, 32, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.L.; Little, J.W.; Starr, F.L.; 3rd Frantz, C.; Vaezy, S. Image-guided HIFU neurolysis of peripheral nerves to treat spasticity and pain. Ultrasound Med. Biol. 2004, 30, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Beyder, A.; Rae, J.L.; Bernard, C.; Strege, P.R.; Sachs, F.; Farrugia, G. Mechanosensitivity of Nav1.5, a voltage-sensitive sodium channel. J. Physiol. 2010, 588, 4969–4985. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.A.; Lin, W.; Morris, T.; Banderali, U.; Juranka, P.F.; Morris, C.E. Membrane trauma and Na+ leak from Nav1.6 channels. Am. J. Physiol. Cell Physiol. 2009, 297, C823–C834. [Google Scholar] [CrossRef]
- Shcherbatko, A.; Ono, F.; Mandel, G.; Brehm, P. Voltage-dependent sodium channel function is regulated through membrane mechanics. Biophys. J. 1999, 77, 1945–1959. [Google Scholar] [CrossRef]
- Honoré, E.; Patel, A.J.; Chemin, J.; Suchyna, T.; Sachs, F. Desensitization of mechano-gated K2P channels. Proc. Natl. Acad. Sci. USA 2006, 103, 6859–6864. [Google Scholar] [CrossRef] [PubMed]
- Tsunozaki, M.; Bautista, D.M. Mammalian somatosensory mechanotransduction. Curr. Opin. Neurobiol. 2009, 19, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.H.; Ranade, S.; Weyer, A.D.; Dubin, A.E.; Baba, Y.; Qiu, Z.; Petrus, M.; Miyamoto, T.; Reddy, K.; Lumpkin, E.A.; et al. Piezo2 is required for Merkel-cell mechanotransduction. Nature 2014, 509, 622–626. [Google Scholar] [CrossRef] [PubMed]
- Ranade, S.S.; Woo, S.H.; Dubin, A.E.; Moshourab, R.A.; Wetzel, C.; Petrus, M.; Mathur, J.; Bégay, V.; Coste, B.; Mainquist, J.; et al. Piezo2 is the major transducer of mechanical forces for touch sensation in mice. Nature 2014, 516, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Heimburg, T.; Jackson, A.D. On soliton propagation in biomembranes and nerves. Proc. Natl. Acad. Sci. USA 2005, 102, 9790–9795. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.G.; Miller, B.A.; Hristova, K.; Usherwood, P.N. Flexoelectric effects in model and native membranes containing ion channels. Eur. Biophys. J. EBJ 1993, 22, 289–300. [Google Scholar] [CrossRef]
- Plaksin, M.; Kimmel, E.; Shoham, S. Cell-type-selective effects of intramembrane cavitation as a unifying theoretical framework for ultrasonic neuromodulation. eNeuro 2016, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Babakhanian, M.; Yang, L.; Nowroozi, B.; Saddik, G.; Boodaghians, L.; Blount, P.; Grundfest, W. Effects of low intensity focused ultrasound on liposomes containing channel proteins. Sci. Rep. 2018, 8, 17250. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| PNS Endings (Research) | Effect | Freq (MHz) | PRF (Hz) | DC (%) | Intensity (W/cm2) | Duration (mSec) | Metric for Modulation | Preparation Type |
|---|---|---|---|---|---|---|---|---|
| Human hand nerve endings [72] (Gavrilov et al., 1977) | Tactile, warm, cold, itch, and pain sensation | 0.48 0.887 2.67 | 100% | 160–30,000 | 1, 10, 100 | Verbal report | Clinical study | |
| Human skin, soft tissue, bone, joint [73] (ab Ithel Davies et al., 1996) | Pain sensation | 0.48–2.67 | 12–15,000 | 1–100 | Verbal report | Clinical study | ||
| Human fingers and upper forearm [74] (Dalecki et al., 1995) | Tactile sensation | 2.2 | 50–1000 | 50% | 150 | 0.1 to 100 | Verbal report | Clinical study |
| Human ear [75] (Tsirulnikov et al., 1988) | Activation of acoustic nerve fibers | 2.5 | 125–8000 | 50% | 1–5 | 0.05–0.1 | Verbal report | Clinical study |
| Human acupuncture point [53] (Yoo et al., 2014) | Deqi sensation | 0.65 | 50 | 10% | 1–3 (SPPA) | 1000 | Verbal report | Clinical study |
| Cat ear [76] (Foster and Wiederhold, 1978) | Activation of auditory nerve | 5 | 100% | 30 | 0.068 | CAP | In vivo | |
| Frog Pacinian corpuscle [77] (Gavrilov et al., 1977) | Activation | 0.48 | 100% | 0.4–2.5 | 0.1 to 100 | CAP | Ex vivo |
| PNS Axons (Research) | Effect | Freq (MHz) | PRF (Hz) | DC (%) | Intensity (W/cm2) | Duration (mSec) | Metric for Modulation | Preparation Type |
|---|---|---|---|---|---|---|---|---|
| Enhanced Activities | ||||||||
| Human median nerve [81] (Moore et al., 2000) | Increase conduction velocity | 1–3 | 50–100% | 1 | 480,000 | Sensory and motor latency | Clinical study | |
| Mouse sciatic nerve [43] (Ilham et al., 2018) | Increase conduction velocity | 1.1 | 200,000 | 20–40% | 0.91–28.2 | 40,000 | Single-unit | Ex vivo |
| Rat posterior tibia nerve [42] (Casella et al., 2017) | Inhibition of rhythmic bladder contraction | 0.25 | 2000 | 0.9 (MPa) | 300 | Bladder contraction | In vivo | |
| Bullfrog sciatic nerve [51] (Tsui et al., 2005) | Enhanced conduction, increased conduction velocity. | 3.5 | 2 | <1% | 1–3 W | CAP | Ex vivo | |
| Crab leg nerve [78] (Saffari et al., 2017) | Direct activation | Ex vivo | ||||||
| Suppressed Activities | ||||||||
| Rabbit sciatic nerve [54] (Foley et al., 2007) | Nerve conduction block | 3.2 | 58% | 1930 (SATA) | 10,000 | Flexion muscle force | In vivo | |
| Rat sciatic nerve [79] (Foley et al., 2008) | Nerve conduction block | 5.7 | 390–7890 (SPTP) | 5000 | Muscle activities | In vivo | ||
| Rat sciatic nerve [80] (Lee et al., 2015) | Nerve conduction block | 2.68 | 2290–2810 (SATA) | 3000–7000 | CAP | Ex vivo | ||
| Rat vagus nerve [47] (Juan et al., 2014) | Inhibition of conduction, reduced conduction velocity | 1.1 | 20–1000 | 18.7–93.4 | 15,000 | CAP | In vivo | |
| Earth worm giant axon [85] (Wahab et al., 2012) | Inhibition of conduction, reduced conduction velocity | 0.825 | 100 | 10% | 0.1–0.7 (MPa) | 15,000–75,000 | CAP | In vivo |
| Frog sciatic nerve [86,87] (Young and Henneman, 1961) | Nerve conduction block | 2.7 | 0.33–0.5 | 11–30% | 1150 (SATA) | 9800 | CAP | Ex vivo |
| Bullfrog sciatic [50] (Colucci et al., 2009) | Nerve conduction block | 0.66–1.98 | 10, 20 | 1–20% | 370 | 30,000 | CAP | Ex vivo |
| Mixed Effects | ||||||||
| Frog sciatic nerve [52] (Mihran et al., 1990) | Enhanced and suppressed excitability | 2–7 | 3–20 kHz | 100–800 (SPTP) | 0.5 | CAP | Ex vivo |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, B.; Chen, L.; Ilham, S.J. A Review on Ultrasonic Neuromodulation of the Peripheral Nervous System: Enhanced or Suppressed Activities? Appl. Sci. 2019, 9, 1637. https://doi.org/10.3390/app9081637
Feng B, Chen L, Ilham SJ. A Review on Ultrasonic Neuromodulation of the Peripheral Nervous System: Enhanced or Suppressed Activities? Applied Sciences. 2019; 9(8):1637. https://doi.org/10.3390/app9081637
Chicago/Turabian StyleFeng, Bin, Longtu Chen, and Sheikh J. Ilham. 2019. "A Review on Ultrasonic Neuromodulation of the Peripheral Nervous System: Enhanced or Suppressed Activities?" Applied Sciences 9, no. 8: 1637. https://doi.org/10.3390/app9081637
APA StyleFeng, B., Chen, L., & Ilham, S. J. (2019). A Review on Ultrasonic Neuromodulation of the Peripheral Nervous System: Enhanced or Suppressed Activities? Applied Sciences, 9(8), 1637. https://doi.org/10.3390/app9081637