1. Introduction
The tomato (
Solanum lycopersicum) is one of the most widely cultivated crops worldwide. In 2022, China led the global tomato production with 68,241,810 tons, followed by India with 20,694,000 tons, and Turkey with 13,000,000 tons [
1]. Tomatoes are an important source of essential nutrients, including vitamins, carotenoids, and phenolic compounds, which contribute to various health benefits such as reduced risks of cancer, osteoporosis, and cardiovascular disease [
2,
3,
4]. Whether consumed raw or cooked, tomatoes play a vital role in human nutrition.
The quality of tomato seeds is crucial for sustaining agricultural productivity, as it directly affects germination, seedling vigor, and the eventual crop yield. With the increase in agricultural production, the risk of seed contamination and infection has also risen, posing a significant threat to crop performance [
5]. Chemical treatments, such as hydrogen peroxide, sorbic acid, peracetic acid, and sodium benzoate, are commonly used to manage seed-borne diseases; however, these methods raise concerns regarding environmental sustainability and safety [
6]. In response to the growing demand for sustainable food systems, the agricultural and food industries are increasingly adopting environmentally friendly technologies that ensure both safety and the preservation of nutritional quality [
7].
The pulsed electric field (PEF) treatment is an advanced non-thermal processing technology that uses short bursts of high-voltage electric pulses (typically 20–80 kV/cm for nanoseconds to microseconds) to inactivate microorganisms while preserving the nutritional and sensory properties of food. These high-intensity electric fields induce electroporation—the formation of transient or permanent pores in cell membranes—without causing significant temperature increases in the treated material. The PEF is valued for its ease of operation, automation potential, and short processing time, making it an attractive alternative to conventional thermal treatments. The technology has demonstrated effectiveness in microbial inactivation, shelf-life extension, and the preservation of bioactive compounds in various liquid foods [
8,
9]. It is widely used in food pasteurization to ensure microbial safety while maintaining quality [
10,
11].
Despite its proven application in liquid foods, the potential of PEFs to improve seed quality—particularly germination and viability—remains underexplored. The processing of solid agricultural materials such as seeds requires novel PEF system designs and treatment chamber configurations to accommodate their physical properties. Recent advancements by our research group have led to the development of a novel PEF system capable of treating dry products such as grains, seeds, and spices. This system has been successfully tested for seed surface disinfection [
12,
13,
14,
15] and for spice treatments aimed at mycotoxin reduction and
Aspergillus spp. inactivation [
16,
17]. However, further studies are necessary to explore the potential of PEF technology to enhance the seed vigor and germination across different seed types. This study aims to evaluate the effects of the PEF treatment on tomato seed germination, stress tolerance, and microbial surface decontamination, contributing to the development of a more sustainable and efficient approach to seed treatments.
3. Results and Discussion
The germination rate of tomato seeds is a critical factor that influences the success of the crop establishment, yield potential, and resource efficiency. A high germination rate ensures a uniform seedling emergence, leading to synchronized plant growth and development, which is essential for optimizing productivity. It minimizes the need for overseeding, reducing input costs and maximizing the efficient use of water, nutrients, and space. Additionally, fast and consistent germination helps seedlings avoid stress and vulnerability to diseases or pests, contributing to healthier and more robust plants. Therefore, improving germination rates is vital for enhancing the overall performance and profitability of tomato cultivation [
20,
21]. The germination rate of tomato seeds increased with both the germination time and the energy applied. In the control samples, the germination rate rose from 0.00 ± 0.00% on the second day to 57.78 ± 1.92% on the seventh day. In comparison, the highest germination rate for PEF-treated tomato seeds increased from 3.33 ± 0.33% at 6.80 J on the second day to 72.81 ± 3.85% at 15.36 J on the seventh day (
p ≤ 0.05). PEF treatments applied at 1.36, 3.21, 5.76, 6.48, and 15.36 J provided a significant increase in the germination rate of the tomato seeds at the seventh day (
p ≤ 0.05) (
Table 2).
While germination is an important criterion in terms of seed strength, the normal seedling ratio of germinated seeds is also important. The normal seedling rate is a crucial parameter for evaluating the quality and viability of tomato seeds, as it directly impacts the successful establishment of healthy plants. A higher rate of normal seedlings indicates that the majority of seeds can develop into robust, well-structured plants capable of achieving optimal growth and productivity. This is particularly important in tomato cultivation, where uniformity in seedling vigor can lead to synchronized flowering, fruiting, and harvest times, thus improving the overall crop management and yield. Additionally, a higher normal seedling rate reduces the need for reseeding and minimizes input costs, ensuring the efficient use of resources such as water, nutrients, and space. Therefore, maintaining a high normal seedling rate is vital for maximizing the economic and agronomic success of tomato production [
22,
23,
24]. The normal seedling rate of the tomato seeds was 73.20 ± 3.67%, and this number ranged from 80.43 ± 1.74% for 1.07 J to 94.62 ± 0.45% for 17.28 J, revealing a significant increase by all PEF treatments (
p ≤ 0.05) (
Table 3).
In practical terms, such increases in seed performance can have meaningful impacts on the crop establishment and overall productivity, especially under suboptimal conditions such as salinity or cold stress. Improved germination ensures a more uniform and timely emergence of seedlings, which is essential for synchronized growth and effective crop management in mechanized agricultural systems.
From a grower’s perspective, a 10–15% increase in germination and seedling vigor can reduce the need for replanting, lower input costs (such as seeds, water, and fertilizers), and enhance the use efficiency of the available arable land. This is particularly critical in stress-prone environments, where abiotic factors often compromise the crop stand and reduce the yield potential. Moreover, early seedling vigor is often associated with an improved nutrient and water uptake efficiency, which can influence downstream developmental stages, including the flowering, fruit set, and final yield.
Germination under cold stress is a critical factor for the tomato seed performance, as it determines the seed’s ability to establish in suboptimal growing conditions. Cold stress can significantly slow down or inhibit germination, leading to poor crop establishment and uneven growth. For tomato seeds, which are often sensitive to low temperatures, the ability to germinate efficiently under cold stress conditions is essential for early-season planting or cultivation in cooler climates. An enhanced cold tolerance during germination not only improves the seedling vigor and uniformity but also reduces the risk of seed decay and disease, which are more prevalent in slower-germinating seeds. Furthermore, successful germination under cold stress can extend the growing season, increase the yield potential, and offer growers greater flexibility in planting schedules [
25,
26,
27].
Tomato seeds were kept in two different environmental conditions, 7 days at 10 °C and 5 days at 24 °C, to determine their germination ability under cold stress. Both the control and PEF-treated samples showed no germination for 7 days at 10 °C; however all samples showed germination for 5 days at 24 °C. While the germination rate for the control samples was 22.22 ± 1.92%, the germination rate for the PEF-treated samples ranged from 12.22 ± 1.92% to 46.67 ± 3.33%. The germination rate of the PEF-treated samples by 1.92, 5.76, 10.88, 15.36, and 17.28 J was significantly higher than that of the control samples (
p ≤ 0.05) (
Table 4).
Electrical conductivity (EC) is a critical parameter in assessing the quality and viability of tomato seeds, as it reflects the integrity of seed membranes. High EC values indicate an increased leakage of electrolytes, typically resulting from damaged or deteriorated seed membranes. This leakage is associated with reduced seed vigor, lower germination rates, and a decreased seedling growth potential. Therefore, monitoring EC provides valuable insights into the physiological state of tomato seeds, enabling the early detection of deterioration and supporting the better management of seed storage and handling practices [
18,
28].
The EC values of the control tomato seed samples measured at 4, 8, and 24 h after the PEF treatment were 75.29 ± 1.88, 77.16 ± 2.38, and 86.04 ± 5.12 µS/cm·g, respectively. The EC of PEF-treated tomato seeds did not exhibit a consistent increasing or decreasing trend with rising energy levels, but a general increase was observed over time following the treatment. The lowest EC values among PEF-treated seeds were recorded at 3.21 J as 59.67 ± 4.21, 60.67 ± 4.13, and 65.38 ± 5.41 µS/cm·g at 4, 8, and 24 h, respectively. In contrast, the highest EC values were observed at 2.16 J as 81.34 ± 4.25, 84.51 ± 4.22, and 92.72 ± 9.51 µS/cm·g at the same time points. As the measurement time increased, the EC values of both the control and PEF-treated samples significantly rose (
p ≤ 0.05). Although some PEF-treated samples exhibited EC values close to those of the control, these differences were not statistically significant after 24 h (
Table 5).
The salt stress tolerance in tomato seeds refers to the ability of seeds to germinate and grow under saline conditions, which is a crucial trait for ensuring crop productivity in areas affected by soil salinity. High salt concentrations can disrupt the water uptake, induce osmotic stress, and cause ion toxicity, leading to reduced seed germination, poor seedling development, and an overall plant growth inhibition. Tomato seed varieties with higher salt tolerances can better maintain their cellular homeostasis, osmotic balance, and ion compartmentalization under saline stress. The identification and cultivation of salt-tolerant tomato varieties is essential for sustainable agriculture in salt-affected regions. This can be achieved through genetic breeding, seed priming techniques, and the use of biostimulants or treatments like PEF to enhance the seed vigor and resilience [
24,
28,
29].
Under the 100 mM NaCl salt stress, the germination rate of both untreated and PEF-treated tomato seeds steadily increased from day 7 to day 12, regardless of the applied energy level. In control seeds, the germination progressed from 0.00 ± 0.00% on day 7 to 30.00 ± 0.60% on day 12. Among the treated groups, the earliest notable germination (23.33 ± 0.60%) was observed on day 7 at 1.07 J. The highest overall germination rate (62.00 ± 0.90%) by day 12 was achieved at 17.28 J, followed closely by 6.48 J (61.00 ± 0.46%) and 1.07 J (60.30 ± 1.90%). Energy levels between 1.07 J and 17.28 J generally produced superior germination performances compared to the control, particularly from day 10 onward. Statistically significant improvements (
p ≤ 0.05) in germination were observed at 1.07, 1.36, 1.92, 3.21, 4.08, 5.35, 5.76, 6.48, 10.88, 15.36, and 17.28 J treatments by day 12. These results suggest that moderate- to high-energy PEF treatments can effectively mitigate the salt-stress-induced germination inhibition in tomato seeds (
Table 6).
Under the 200 mM NaCl salt stress, the germination of untreated tomato seeds remained limited, increasing only from 0.00 ± 0.00% on day 7 to 13.30 ± 0.00% by day 12. In contrast, the PEF treatment enhanced the germination across various energy levels, with the earliest notable germination on day 7 observed at 3.21 J (3.30 ± 0.40%). By day 12, the highest germination rate (53.03 ± 0.00%) was recorded at 10.80 J, followed by 17.28 J (50.00 ± 0.60%) and 15.36 J (43.30 ± 0.00%). Energy levels between 3.21 J and 17.28 J were particularly effective in improving the germination under high salt stress. Treatments at 1.07, 1.36, 1.92, 2.16, 3.21, 4.08, 6.48, 6.80, 8.55, 9.60, 10.80, 15.36, and 17.28 J yielded significantly higher germination rates than the control by day 12 (
p ≤ 0.05), suggesting that PEFs can alleviate the salt-induced germination inhibition in tomato seeds at moderate- to high-energy applications (
Table 7).
The presence of a high number of total bacteria on tomato seeds may indicate contamination that could negatively impact seed health. Some bacteria can cause seed rot, reduce germination rates, or even transmit seed-borne diseases, which may affect young seedlings and lower the overall crop yield. Fungal contamination, particularly by molds and yeasts, can damage seeds by colonizing their surfaces, leading to decay or reduced seedling vigor. Fungal infections may weaken the seed coat, resulting in improper or delayed germination. Elevated levels of mold and yeast can also create an inhospitable environment for seed growth by consuming nutrients or producing toxic metabolites that inhibit sprouting [
30]. Therefore, the inactivation of the endogenous microflora is crucial, as both bacteria and molds can adversely affect the germination process. The mean initial counts of TMAB and TMY in tomato seeds were 4.00 ± 0.03 and 3.06 ± 0.03 log CFU/g, respectively. Both TMAB and TMY counts were reduced to non-detectable levels following the application of 17.28 kV/cm. All PEF treatments resulted in significant reductions in the initial mean counts of TMAB and TMY (
p ≤ 0.05) (
Table 8).
Seedling development and root formation are critical stages in the early growth of plants, laying the foundation for overall health, productivity, and resilience. A strong, healthy seedling with a well-developed root system is essential for efficient nutrient and water uptake, which directly influences the plant’s growth rate, vigor, and ability to withstand environmental stressors, such as drought, pests, and disease. Proper root formation ensures a firm anchorage in the soil, reducing the risk of lodging or uprooting under adverse conditions. Additionally, the root system plays a key role in supporting the above-ground development by facilitating the absorption of essential nutrients necessary for photosynthesis and overall plant metabolism. An early and robust seedling growth increases the likelihood of higher yields, enabling the plant to reach its full genetic potential in terms of both productivity and quality [
31].
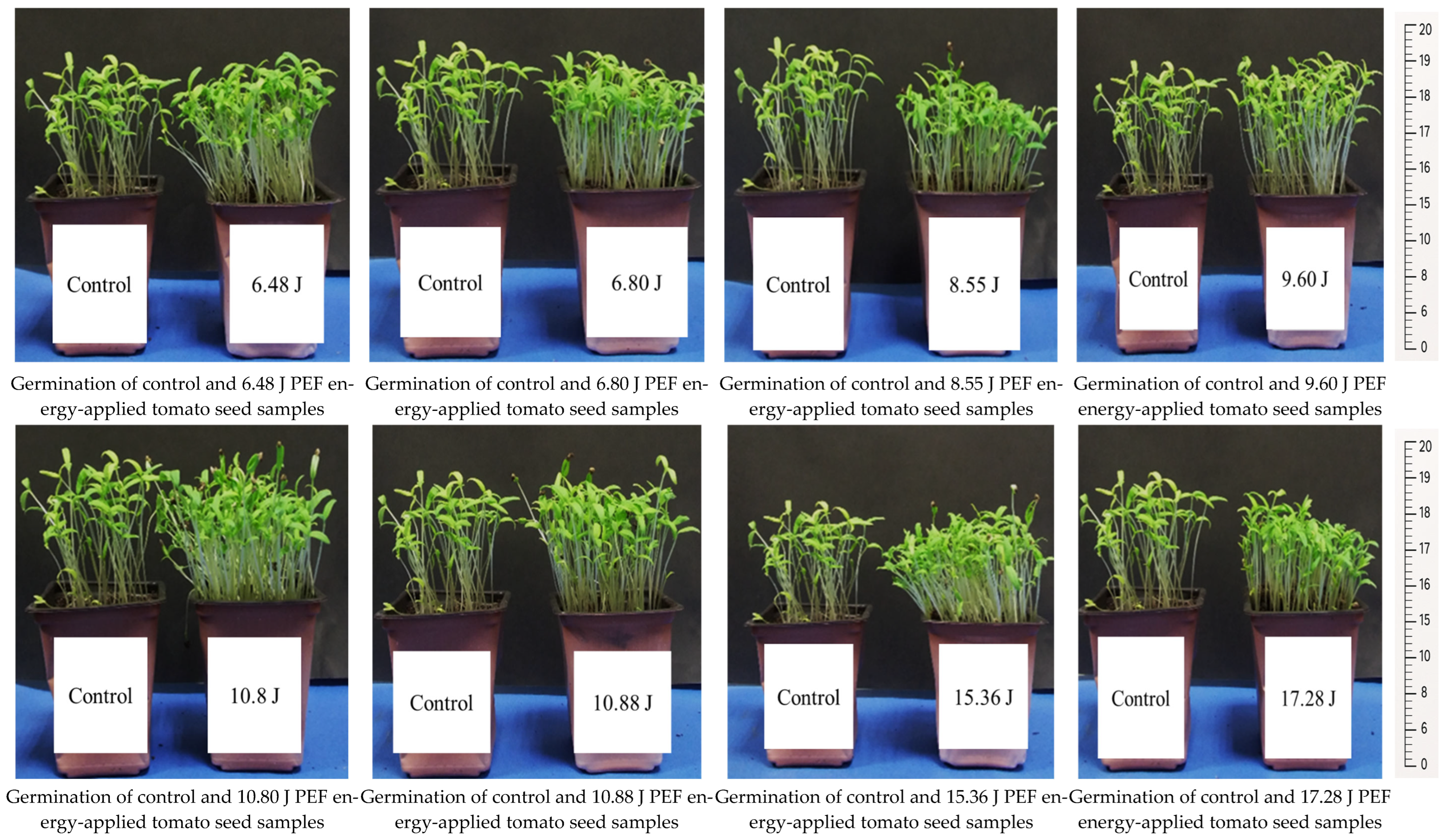
The seedling development and root formation in both control and PEF-treated tomato seeds revealed that the number of seedlings was higher in the PEF-treated group, despite the same number of seeds being planted. The PEF-treated seedlings also exhibited a stronger growth, with thicker stems compared to the control samples (
Figure 1). Additionally, they showed a more robust root formation than the control group (
Figure 2). Germination studies under salt stress indicated that the number of germinated seedlings was higher in the PEF-treated group under both 100 mM and 200 mM salt stress conditions. Furthermore, PEF-treated seeds produced taller seedlings with a stronger structural development (
Figure 3 and
Figure 4).
The inactivation mechanism of the PEF treatment is explained by thermal effects, electroporation, and the formation of reactive oxygen species (ROS). The high-intensity PEF application leads to a temperature increase in and around the cell, which can cause the denaturation of cellular proteins and the disruption of the cell membrane structure. Electroporation occurs when the permeability of the cell membrane increases due to the application of an electric field. During this process, temporary pores form in the membrane, and if the electric field is strong enough, the damage becomes permanent, ultimately resulting in cell death [
32,
33]. Additionally, ROS may be generated under high electric fields, and these free radicals can damage the cell membrane and internal components, contributing further to cell death. The effectiveness of this inactivation mechanism depends on several factors, including the frequency, duration, energy density, waveform, and pulse width of the PEF application [
34,
35].
Tomato seeds exposed to pulsed electromagnetic fields with pulse energies of 35–80 J/pulse for 10 to 15 min demonstrated enhanced plant growth [
36]. Similarly, tomato seeds subjected to alternating current (AC) electric fields of 4–12 kV/cm and AC magnetic fields of 3–1000 G for 15–60 s germinated 1.1–2.8 times faster than control seeds. However, the application of a 12 kV electric field for 60 s had an inhibitory effect. It was also observed that germination rates increased by 1.1 to 2.8 times following the electric field treatment [
37]. Another study reported that tomato seeds treated with an electric field of 20 kV/cm for 20 s achieved a 100% germination rate, compared to 76% in untreated seeds [
38].
The disruption of the cell membrane by PEFs leads to an imbalance in the membrane’s transport system, resulting in increased permeability. This elevated permeability enhances the nutrient and water uptake from the soil. Permeability is typically assessed by measuring changes in EC. An increase in the EC of the surrounding medium reflects the electrolyte leakage from seeds into water, indicating membrane damage [
39]. In corn grains, a significant increase in EC was observed over time, suggesting that cell damage may be more time-dependent, rather than being directly caused by the PEF treatment. Similar results were reported for cucumber [
13], corn [
12], and wheat grains [
18], where EC increases were primarily attributed to time rather than the PEF application itself.
Salinity is a major factor reducing crop yields, as high salt concentrations cause hyperosmotic stress and ion imbalance [
40], necessitating mechanisms for maintaining homeostasis, detoxification, and growth to develop salt tolerance. Salinity stress adversely affects plant growth by inducing osmotic stress and ionic toxicity, which hinder photosynthesis, nutrient uptake, and the overall development [
41]. However, salinity tolerance remains a complex and not fully understood process. The PEF treatment may enhance the salt tolerance under 100 mM and 200 mM NaCl stress conditions, possibly by activating one or more of the plant’s stress tolerance mechanisms. An increased salt tolerance following PEF treatments has also been reported in cucumber seeds [
13], corn [
12], and wheat grains [
18].
The natural microflora of seeds, including bacteria, can influence the seed germination and seedling growth. While various chemicals, such as calcium cyanamide (CaCN₂), are used to disinfect seeds and improve productivity, chemical treatments have drawbacks, including adverse effects on worker safety and the environment [
42,
43]. As a more sustainable alternative, the PEF treatment has shown promise for seed surface disinfection. For example, a PEF application resulted in a 63.33% reduction in
Aspergillus parasiticus in corn seeds [
12], a 64.37% reduction in red pepper flakes [
16], and a 60% reduction in sesame seeds [
17]. Significant reductions in the endogenous microflora of wheat grains [
18,
44], cucumber seeds [
13], and corn grains [
12] were also achieved at higher energy levels. Similarly, the substantial inactivation of the total aerobic mesophilic bacteria (TAMB) and total mold and yeast (TMY) was observed in cabbage, lettuce, garden rocket, wheat [
15,
18], cucumber [
13], and corn [
12] under high-energy PEF treatments. Additionally, PEFs have been effective in inactivating seed-borne pathogens, such as
Drechslera graminea,
Xanthomonas campestris pv.
campestris,
Fusarium graminearum, and
Alternaria brassicae, on red cabbage seeds with an increased treatment time and frequency [
14].










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}