
Exploring the Therapeutic Potential of Spilanthol from Acmella paniculata (Wall ex DC.) R. K. Jansen in Attenuating Neurodegenerative Diseases: A Multi-Faceted Approach Integrating In Silico and In Vitro Methodologies

 , , , , and
, , , , and 
Abstract
1. Introduction
2. Results and Discussion
2.1. In Silico ADMET Prediction
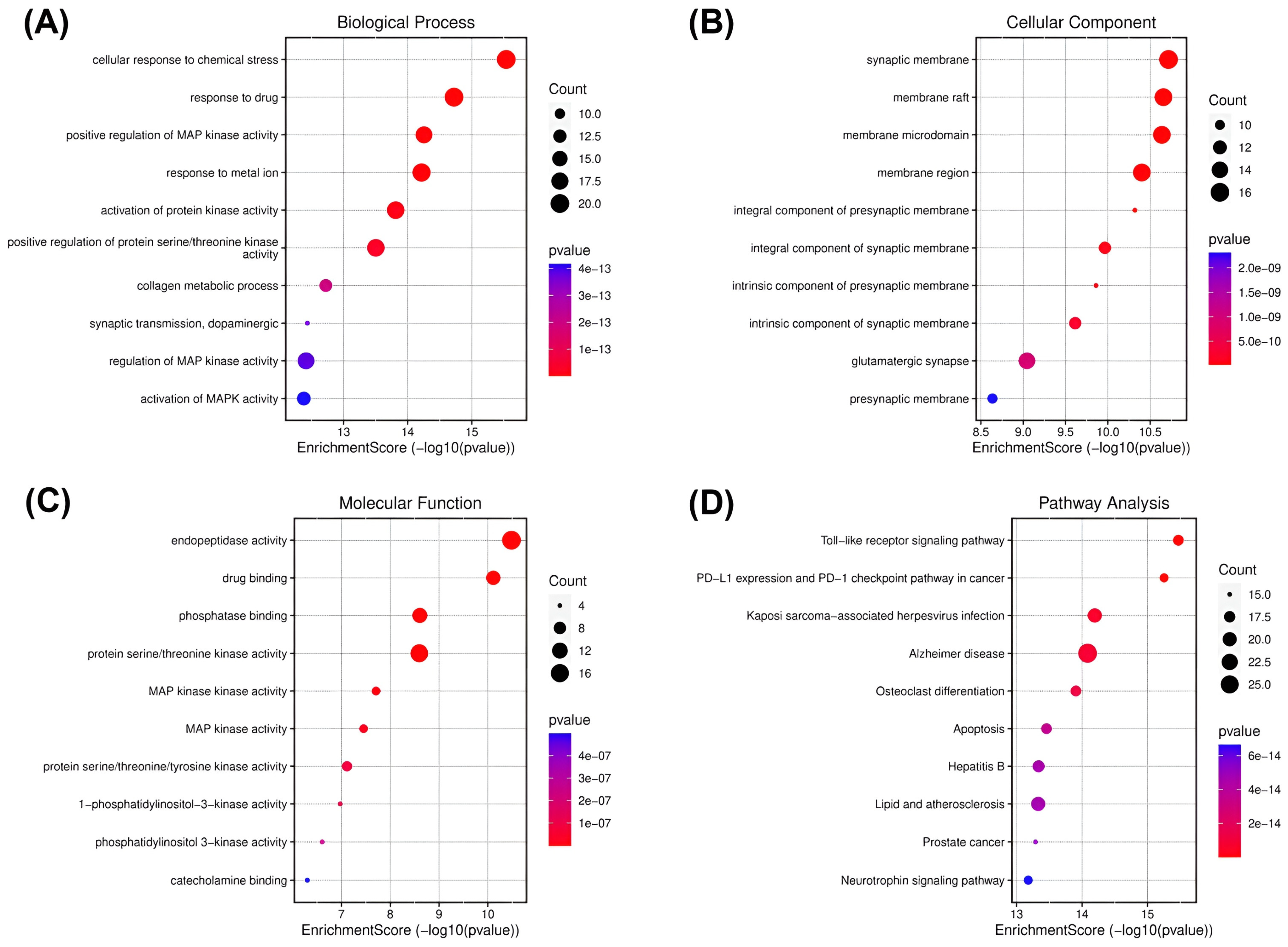
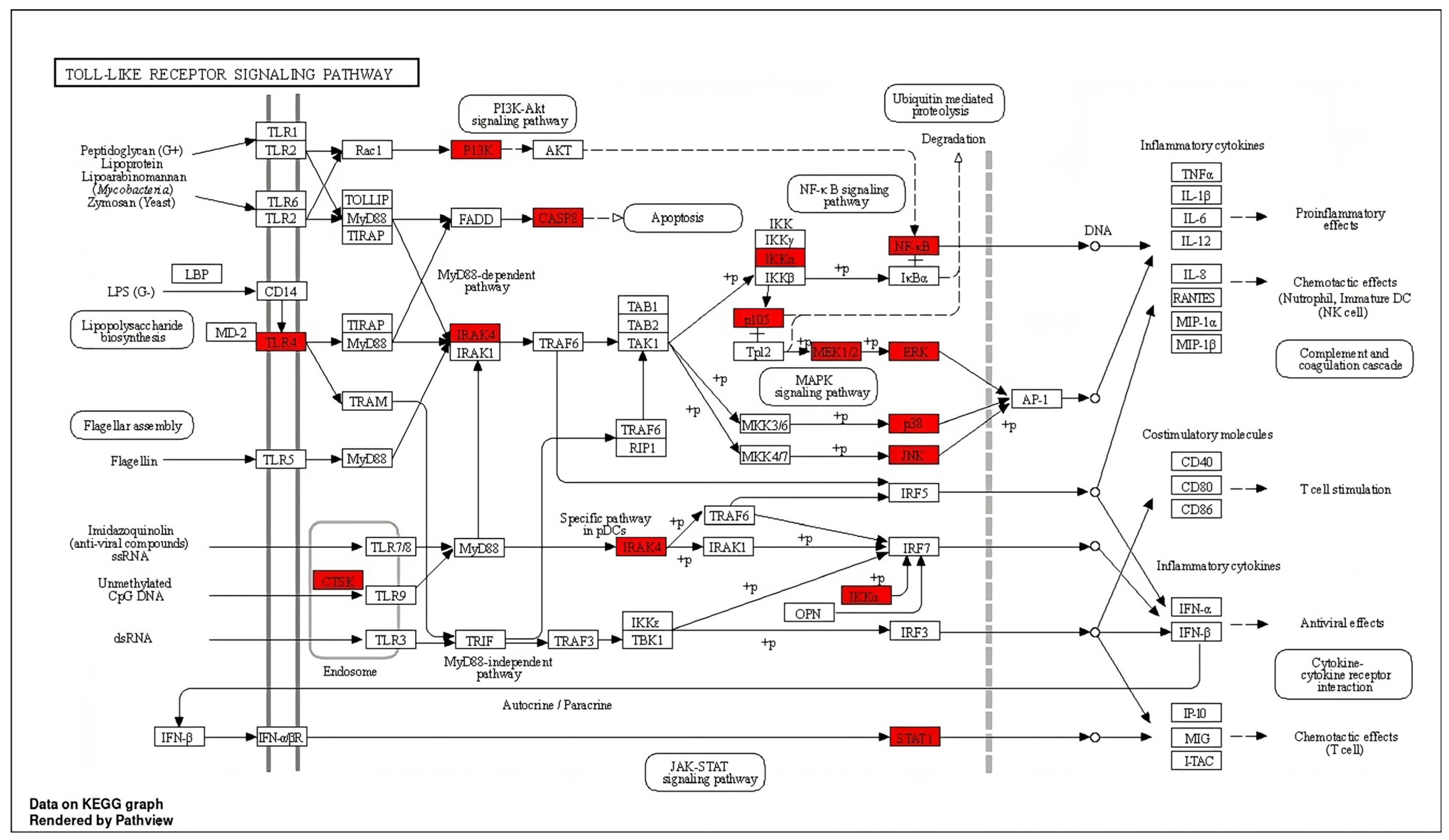
2.2. Network Pharmacology Analysis
2.3. Molecular Docking and Molecular Dynamics Simulation Analysis
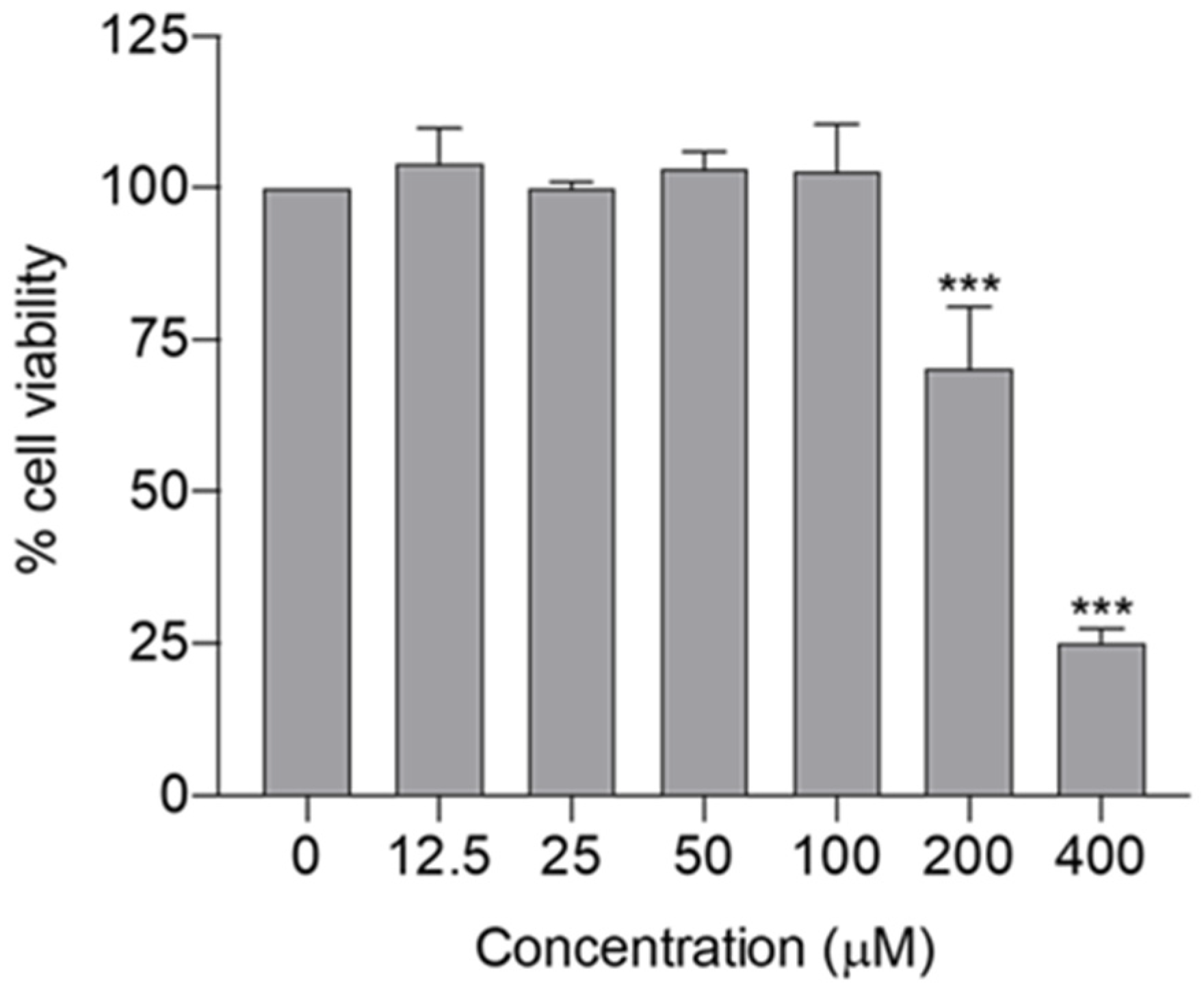
2.4. Assessment of Cytotoxicity of Spilanthol in BV-2 Microglial Cells
2.5. Evaluation of the Anti-Neuroinflammatory Efficacy of Spilanthol
3. Conclusions
4. Materials and Methods
4.1. Materials and Chemicals
4.2. In Silico ADMET Screening
4.3. Network Pharmacology Analysis
4.3.1. Acquisition of the Target Genes of Spilanthol
4.3.2. Acquisition of the Target Genes of Neurodegenerative Diseases
4.3.3. Construction of Protein–Protein Interaction (PPI) Network
4.3.4. GO Function Enrichment and KEGG Pathway Analysis
4.4. Dynamic Interactions: Insights from Docking and Molecular Dynamics
4.5. Extraction and Isolation
4.6. In Vitro Evaluation
4.6.1. BV-2 Cell Culture
4.6.2. Evaluation of the Cytotoxicity of Spilanthol
4.6.3. Anti-Neuroinflammatory Efficacy of Spilanthol
4.6.4. Nitrite Assay
4.6.5. Determination of IL-6 and TNF-α Levels Using ELISA
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TLR4 | Toll-like receptor 4 |
| NF-κB | Nuclear factor kappa B |
| IKKα | Inhibitory kappa B kinase α |
| IRAK4 | Interleukin-1 receptor-associated kinase 4 |
| MD2 | Myeloid differentiation factor 2 |
| MyD88 | Myeloid differentiation primary response protein 88 |
| IL-6 | Interleukin-6 |
| IL-1 | Interleukin-1 |
| TNF-α | Tumor necrosis factor-alpha |
| NO | Nitric oxide |
| iNOS | Inducible nitric oxide synthase |
| COX-2 | Cyclooxygenase-2 |
| LPS | Lipopolysaccharide |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| DMSO | Dimethyl sulfoxide |
| ELISA | Enzyme-linked immunosorbent assay |
| PPI | Protein–protein interactions |
| MCC | Maximal clique centrality |
| GO | Gene ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| DMEM | Dulbecco’s modified Eagle’s medium |
| FBS | Fetal bovine serum |
| PBS | Phosphate buffered saline |
| RMSD | Root-mean-square displacement |
| PDB | Protein data bank |
References
- Liu, S.; Zheng, G.; Chen, H.; Li, G.; Guo, X. Advances in integrative medicine for neurodegenerative diseases: From basic research to clinical practice. Front. Neurol. 2023, 14, 1197641. [Google Scholar] [CrossRef]
- Cai, C.; Fang, J.; Ke, H. Traditional medicine and phytochemicals for neurodegenerative diseases treatment: Application of interdisciplinary technologies in novel therapeutic target and drug discovery. Front. Neurosci. 2023, 17, 1268710. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.P.; Brown, R.H., Jr.; Cleveland, D.W. Decoding ALS: From genes to mechanism. Nature 2016, 539, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Yacoubian, T. Neurodegenerative disorders: Why do we need new therapies? In Drug Discovery Approaches for the Treatment of Neurodegenerative Disorders; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–16. [Google Scholar]
- Rubino, S.J.; Mayo, L.; Wimmer, I.; Siedler, V.; Brunner, F.; Hametner, S.; Madi, A.; Lanser, A.; Moreira, T.; Donnelly, D. Acute microglia ablation induces neurodegeneration in the somatosensory system. Nat. Commun. 2018, 9, 4578. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, D.; Mao, Q.; Xia, H. Role of neuroinflammation in neurodegeneration development. Signal Transduct. Target. Ther. 2023, 8, 267. [Google Scholar] [CrossRef]
- Leng, F.; Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: Where do we go from here? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; Zhou, J. Neuroinflammation in Parkinson’s disease and its potential as therapeutic target. Transl. Neurodegener. 2015, 4, 19. [Google Scholar] [CrossRef]
- Du, L.; Zhang, Y.; Chen, Y.; Zhu, J.; Yang, Y.; Zhang, H.-L. Role of microglia in neurological disorders and their potentials as a therapeutic target. Mol. Neurobiol. 2017, 54, 7567–7584. [Google Scholar] [CrossRef]
- Gao, Y.; Tu, D.; Yang, R.; Chu, C.-H.; Hong, J.-S.; Gao, H.-M. Through reducing ROS production, IL-10 suppresses caspase-1-dependent IL-1β maturation, thereby preventing chronic neuroinflammation and neurodegeneration. Int. J. Mol. Sci. 2020, 21, 465. [Google Scholar] [CrossRef]
- Abd Ghafar, S.A.; Salehuddin, N.S.; Abdul Rahman, N.Z.; Halib, N.; Mohamad Hanafiah, R. Transcriptomic profile analysis of Streptococcus mutans response to Acmella paniculata flower extracts. Evid. -Based Complement. Altern. Med. 2022, 2022, 7767940. [Google Scholar] [CrossRef]
- Meitei, L.R.; De, A.; Mao, A.A. An ethnobotanical study on the wild edible plants used by forest dwellers in Yangoupokpi Lokchao Wildlife Sanctuary, Manipur, India. Ethnobot. Res. Appl. 2022, 23, 5–18. [Google Scholar] [CrossRef]
- Panyadee, P.; Inta, A. Taxonomy and ethnobotany of Acmella (Asteraceae) in Thailand. Biodiversitas J. Biol. Divers. 2022, 23. [Google Scholar] [CrossRef]
- Patel, S.; Purohit, N.; Sapra, P.; Solanki, H.; Bishoyi, A.K. Genetic diversity analysis revealed the hot spot of Acmella paniculata (Wall ex DC.) RK Jansen existing in natural populations of Gujarat. Genet. Resour. Crop Evol. 2022, 69, 2249–2260. [Google Scholar] [CrossRef]
- Barbosa, A.F.; Carvalho, M.G.d.; Smith, R.E.; Sabaa-Srur, A.U. Spilanthol: Occurrence, extraction, chemistry and biological activities. Rev. Bras. Farmacogn. 2016, 26, 128–133. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Alegría-Arcos, M.; Barbosa, T.; Sepúlveda, F.; Combariza, G.; González, J.; Gil, C.; Martínez, A.; Ramírez, D. Network pharmacology reveals multitarget mechanism of action of drugs to be repurposed for COVID-19. Front. Pharmacol. 2022, 13, 952192. [Google Scholar] [CrossRef]
- Sadybekov, A.V.; Katritch, V. Computational approaches streamlining drug discovery. Nature 2023, 616, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Agu, P.; Afiukwa, C.; Orji, O.; Ezeh, E.; Ofoke, I.; Ogbu, C.; Ugwuja, E.; Aja, P. Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in diseases management. Sci. Rep. 2023, 13, 13398. [Google Scholar] [CrossRef]
- Sakhawat, A.; Khan, M.U.; Rehman, R.; Khan, S.; Shan, M.A.; Batool, A.; Javed, M.A.; Ali, Q. Natural compound targeting BDNF V66M variant: Insights from in silico docking and molecular analysis. AMB Express 2023, 13, 134. [Google Scholar] [CrossRef]
- Jia, C.-Y.; Li, J.-Y.; Hao, G.-F.; Yang, G.-F. A drug-likeness toolbox facilitates ADMET study in drug discovery. Drug Discov. Today 2020, 25, 248–258. [Google Scholar] [CrossRef]
- Abdullahi, M.; Uzairu, A.; Shallangwa, G.A.; Mamza, P.A.; Ibrahim, M.T.; Ahmad, I.; Patel, H. Structure-based drug design, molecular dynamics simulation, ADMET, and quantum chemical studies of some thiazolinones targeting influenza neuraminidase. J. Biomol. Struct. Dyn. 2023, 41, 13829–13843. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed]
- Stein, R.; Berger, M.; Santana de Cecco, B.; Mallmann, L.P.; Terraciano, P.B.; Driemeier, D.; Rodrigues, E.; Beys-da-Silva, W.O.; Konrath, E.L. Chymase inhibition: A key factor in the anti-inflammatory activity of ethanolic extracts and spilanthol isolated from Acmella oleracea. J. Ethnopharmacol. 2021, 270, 113610. [Google Scholar] [CrossRef] [PubMed]
- Nomura, E.C.O.; Rodrigues, M.R.A.; da Silva, C.F.; Hamm, L.A.; Nascimento, A.M.; de Souza, L.M.; Cipriani, T.R.; Baggio, C.H.; Werner, M.F.d.P. Antinociceptive effects of ethanolic extract from the flowers of Acmella oleracea (L.) R.K. Jansen in mice. J. Ethnopharmacol. 2013, 150, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.T.; Peretti, P.; Bezerra, R.M.; Biancardi, M.F.; Sousa, F.F.; Mendes, E.P.; Dutra, J.B.; Silveira, C.C.; Castro, C.H.; Cruz, J.N. Pharmacological Characteristics of the Hydroethanolic Extract of Acmella oleracea (L) RK Jansen Flowers: ADME/Tox In Silico and In Vivo Antihypertensive and Chronic Toxicity Evaluation. Evid. -Based Complement. Altern. Med. 2023, 2023, 1278720. [Google Scholar] [CrossRef] [PubMed]
- Souza, G.; Silva, I.; Viana, M.D.; Melo, N.; Sánchez-Ortiz, B.; Oliveira, M.; Barbosa, W.; Ferreira, I.M.; Carvalho, J. Acute Toxicity of the Hydroethanolic Extract of the Flowers of Acmella oleracea L. in Zebrafish (Danio rerio): Behavioral and Histopathological Studies. Pharmaceuticals 2019, 12, 173. [Google Scholar]
- De Souza, G.; Viana, M.; Goés, L.; Sanchez-Ortiz, B.; Silva, G.d.; Pinheiro, W.d.S.; Santos, C.R.D.; Carvalho, J.T. Reproductive toxicity of the hydroethanolic extract of the flowers of Acmella oleracea and spilanthol in zebrafish: In vivo and in silico evaluation. Hum. Exp. Toxicol. 2020, 39, 127–146. [Google Scholar] [CrossRef] [PubMed]
- de Souza, G.C.; Matias Pereira, A.C.; Viana, M.D.; Ferreira, A.M.; da Silva, I.D.R.; de Oliveira, M.M.R.; Barbosa, W.L.R.; Silva, L.B.; Ferreira, I.M.; Dos Santos, C.B.R. Acmella oleracea (L) RK Jansen reproductive toxicity in zebrafish: An in vivo and in silico assessment. Evid.-Based Complement. Altern. Med. 2019, 2019, 1237301. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Zhao, Y.; Yang, T.; Gao, X.; Meng, C. Duhuo Jisheng decoction alleviates neuroinflammation and neuropathic pain by suppressing microglial M 1 polarization: A network pharmacology research. J. Orthop. Surg. Res. 2023, 18, 629. [Google Scholar] [CrossRef]
- Guo, S.; Wang, H.; Yin, Y. Microglia polarization from M1 to M2 in neurodegenerative diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Xu, Y.-R.; Lei, C.-Q. TAK1-TABs complex: A central signalosome in inflammatory responses. Front. Immunol. 2021, 11, 608976. [Google Scholar] [CrossRef]
- Okun, E.; Griffioen, K.J.; Lathia, J.D.; Tang, S.-C.; Mattson, M.P.; Arumugam, T.V. Toll-like receptors in neurodegeneration. Brain Res. Rev. 2009, 59, 278–292. [Google Scholar] [CrossRef]
- Azam, S.; Jakaria, M.; Kim, I.-S.; Kim, J.; Haque, M.E.; Choi, D.-K. Regulation of toll-like receptor (TLR) signaling pathway by polyphenols in the treatment of age-linked neurodegenerative diseases: Focus on TLR4 signaling. Front. Immunol. 2019, 10, 1000. [Google Scholar] [CrossRef]
- Su, Y.; Wang, D.; Liu, N.; Yang, J.; Sun, R.; Zhang, Z. Clostridium butyricum improves cognitive dysfunction in ICV-STZ-induced Alzheimer’s disease mice via suppressing TLR4 signaling pathway through the gut-brain axis. PLoS ONE 2023, 18, e0286086. [Google Scholar] [CrossRef]
- Cui, W.; Sun, C.; Ma, Y.; Wang, S.; Wang, X.; Zhang, Y. Inhibition of TLR4 induces M2 microglial polarization and provides neuroprotection via the NLRP3 inflammasome in Alzheimer’s disease. Front. Neurosci. 2020, 14, 444. [Google Scholar] [CrossRef]
- Kouli, A.; Horne, C.; Williams-Gray, C. Toll-like receptors and their therapeutic potential in Parkinson’s disease and α-synucleinopathies. BrainBehav. Immun. 2019, 81, 41–51. [Google Scholar] [CrossRef]
- Heidari, A.; Yazdanpanah, N.; Rezaei, N. The role of Toll-like receptors and neuroinflammation in Parkinson’s disease. J. Neuroinflammation 2022, 19, 135. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, W.-M.; Tang, P.-C.; Zhang, X.; Zhang, X.-Y.; Yu, B.-C.; Fan, Y.-Y.; Ge, X.-Q.; Zhang, X.-L. Neuroprotective effects of natural cordycepin on LPS-induced Parkinson’s disease through suppressing TLR4/NF-κB/NLRP3-mediated pyroptosis. J. Funct. Foods 2020, 75, 104274. [Google Scholar] [CrossRef]
- Sharma, V.K.; Singh, T.G.; Singh, S.; Garg, N.; Dhiman, S. Apoptotic Pathways and Alzheimer’s Disease: Probing Therapeutic Potential. Neurochem. Res. 2021, 46, 3103–3122. [Google Scholar] [CrossRef]
- Mohamed Asik, R.; Suganthy, N.; Aarifa, M.A.; Kumar, A.; Szigeti, K.; Mathe, D.; Gulyás, B.; Archunan, G.; Padmanabhan, P. Alzheimer’s disease: A molecular view of β-amyloid induced morbific events. Biomedicines 2021, 9, 1126. [Google Scholar] [CrossRef]
- Jagust, W. Imaging the evolution and pathophysiology of Alzheimer disease. Nat. Rev. Neurosci. 2018, 19, 687–700. [Google Scholar] [CrossRef]
- Abubakar, M.B.; Sanusi, K.O.; Ugusman, A.; Mohamed, W.; Kamal, H.; Ibrahim, N.H.; Khoo, C.S.; Kumar, J. Alzheimer’s disease: An update and insights into pathophysiology. Front. Aging Neurosci. 2022, 14, 742408. [Google Scholar] [CrossRef] [PubMed]
- Muzio, L.; Viotti, A.; Martino, G. Microglia in neuroinflammation and neurodegeneration: From understanding to therapy. Front. Neurosci. 2021, 15, 742065. [Google Scholar] [CrossRef]
- Rahimifard, M.; Maqbool, F.; Moeini-Nodeh, S.; Niaz, K.; Abdollahi, M.; Braidy, N.; Nabavi, S.M.; Nabavi, S.F. Targeting the TLR4 signaling pathway by polyphenols: A novel therapeutic strategy for neuroinflammation. Ageing Res. Rev. 2017, 36, 11–19. [Google Scholar] [CrossRef]
- Spinozzi, E.; Pavela, R.; Bonacucina, G.; Perinelli, D.R.; Cespi, M.; Petrelli, R.; Cappellacci, L.; Fiorini, D.; Scortichini, S.; Garzoli, S. Spilanthol-rich essential oil obtained by microwave-assisted extraction from Acmella oleracea (L.) RK Jansen and its nanoemulsion: Insecticidal, cytotoxic and anti-inflammatory activities. Ind. Crops Prod. 2021, 172, 114027. [Google Scholar] [CrossRef]
- Wu, L.-C.; Fan, N.-C.; Lin, M.-H.; Chu, I.-R.; Huang, S.-J.; Hu, C.-Y.; Han, S.-Y. Anti-inflammatory effect of spilanthol from Spilanthes acmella on murine macrophage by down-regulating LPS-induced inflammatory mediators. J. Agric. Food Chem. 2008, 56, 2341–2349. [Google Scholar] [CrossRef] [PubMed]
- Polley, S.; Passos, D.O.; Huang, D.-B.; Mulero, M.C.; Mazumder, A.; Biswas, T.; Verma, I.M.; Lyumkis, D.; Ghosh, G. Structural Basis for the Activation of IKK1/α. Cell Rep. 2016, 17, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.W.; Rey, F.A.; Sodeoka, M.; Verdine, G.L.; Harrison, S.C. Structure of the NF-κB p50 homodimer bound to DNA. Nature 1995, 373, 311–317. [Google Scholar] [CrossRef]
- Meng, E.C.; Goddard, T.D.; Pettersen, E.F.; Couch, G.S.; Pearson, Z.J.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Tools for structure building and analysis. Protein Sci. 2023, 32, e4792. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [PubMed]
- Dolinsky, T.J.; Czodrowski, P.; Li, H.; Nielsen, J.E.; Jensen, J.H.; Klebe, G.; Baker, N.A. PDB2PQR: Expanding and upgrading automated preparation of biomolecular structures for molecular simulations. Nucleic Acids Res. 2007, 35, W522–W525. [Google Scholar] [CrossRef]
- Case, D.A.; Aktulga, H.M.; Belfon, K.; Cerutti, D.S.; Cisneros, G.A.; Cruzeiro, V.W.D.; Forouzesh, N.; Giese, T.J.; Götz, A.W.; Gohlke, H.; et al. AmberTools. J. Chem. Inf. Model. 2023, 63, 6183–6191. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Kasavajhala, K.; Belfon, K.A.; Raguette, L.; Huang, H.; Migues, A.N.; Bickel, J.; Wang, Y.; Pincay, J.; Wu, Q. ff19SB: Amino-acid-specific protein backbone parameters trained against quantum mechanics energy surfaces in solution. J. Chem. Theory Comput. 2019, 16, 528–552. [Google Scholar] [CrossRef]
- He, X.; Man, V.H.; Yang, W.; Lee, T.-S.; Wang, J. A fast and high-quality charge model for the next generation general AMBER force field. J. Chem. Phys. 2020, 153, 114502. [Google Scholar] [CrossRef]
- Wang, J.; Cieplak, P.; Kollman, P.A. How well does a restrained electrostatic potential (RESP) model perform in calculating conformational energies of organic and biological molecules? J. Comput. Chem. 2000, 21, 1049–1074. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E., III. PTRAJ and CPPTRAJ: Software for processing and analysis of molecular dynamics trajectory data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- de Freitas Blanco, V.S.; Michalak, B.; Zelioli, Í.A.M.; de Oliveira, A.d.S.S.; Rodrigues, M.V.N.; Ferreira, A.G.; Garcia, V.L.; Cabral, F.A.; Kiss, A.K.; Rodrigues, R.A.F. Isolation of spilanthol from Acmella oleracea based on Green Chemistry and evaluation of its in vitro anti-inflammatory activity. J. Supercrit. Fluids 2018, 140, 372–379. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | PDB ID | Inhibitor | GOLD Fitness Score | |
|---|---|---|---|---|
| Inhibitor | Spilanthol | |||
| TLR-4 | 2Z65 | CID6912404 | 141.86 | 58.29 |
| NF-κB 1 | 1SVC | CID5280961 | 49.29 | 49.73 |
| IRAK4 | 6BFN | CID 44449334 | 81.29 | 59.38 |
| IKKα | 5EBZ | CID16048085 | 57.99 | 59.70 |
| MD2 | 2E56 | CID11005 | 58.78 | 58.64 |
| iNOS | 1DD7 | CID16116169 | 93.52 | 73.91 |
| COX-2 | 5IKR | CID5090 | 66.46 | 62.13 |
| MyD88 | 2JS7 | ST2825 | 61.55 | 44.96 |
| IKKα | NF-kB1 | |||
|---|---|---|---|---|
| Component (avg ± SD) | CID16048085 | Spilanthol | CID5280961 | Spilanthol |
| ΔEvdW | −8.64 ± 16.31 | −34.48 ± 2.62 | −1.24 ± 4.94 | −19.80 ± 16.24 |
| ΔEc | −1.37 ± 3.20 | −2.87 ± 2.67 | −0.44 ± 1.75 | −1.20 ± 1.43 |
| γΔMSA | 2.82 ± 5.50 | 7.63 ± 2.15 | 0.37 ± 1.49 | 3.54 ± 3.03 |
| ΔGR | −1.57 ± 2.96 | −6.15 ± 0.37 | −0.20 ± 0.78 | −3.27 ± 2.68 |
| C | −2.89 | |||
| α | 0.104758 | |||
| aΔGbind | −3.81 ± 1.74 | −6.65 ± 0.33 | −3.05 ± 0.62 | −5.06 ± 1.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayashan, S.S.; Darai, N.; Rungrotmongkol, T.; Dasuni Wasana, P.W.; Nwe, S.Y.; Thongphichai, W.; Suriyakala, G.; Towiwat, P.; Sukrong, S. Exploring the Therapeutic Potential of Spilanthol from Acmella paniculata (Wall ex DC.) R. K. Jansen in Attenuating Neurodegenerative Diseases: A Multi-Faceted Approach Integrating In Silico and In Vitro Methodologies. Appl. Sci. 2024, 14, 3755. https://doi.org/10.3390/app14093755
Jayashan SS, Darai N, Rungrotmongkol T, Dasuni Wasana PW, Nwe SY, Thongphichai W, Suriyakala G, Towiwat P, Sukrong S. Exploring the Therapeutic Potential of Spilanthol from Acmella paniculata (Wall ex DC.) R. K. Jansen in Attenuating Neurodegenerative Diseases: A Multi-Faceted Approach Integrating In Silico and In Vitro Methodologies. Applied Sciences. 2024; 14(9):3755. https://doi.org/10.3390/app14093755
Chicago/Turabian StyleJayashan, Sanith Sri, Nitchakan Darai, Thanyada Rungrotmongkol, Peththa Wadu Dasuni Wasana, San Yoon Nwe, Wisuwat Thongphichai, Gunasekaran Suriyakala, Pasarapa Towiwat, and Suchada Sukrong. 2024. "Exploring the Therapeutic Potential of Spilanthol from Acmella paniculata (Wall ex DC.) R. K. Jansen in Attenuating Neurodegenerative Diseases: A Multi-Faceted Approach Integrating In Silico and In Vitro Methodologies" Applied Sciences 14, no. 9: 3755. https://doi.org/10.3390/app14093755
APA StyleJayashan, S. S., Darai, N., Rungrotmongkol, T., Dasuni Wasana, P. W., Nwe, S. Y., Thongphichai, W., Suriyakala, G., Towiwat, P., & Sukrong, S. (2024). Exploring the Therapeutic Potential of Spilanthol from Acmella paniculata (Wall ex DC.) R. K. Jansen in Attenuating Neurodegenerative Diseases: A Multi-Faceted Approach Integrating In Silico and In Vitro Methodologies. Applied Sciences, 14(9), 3755. https://doi.org/10.3390/app14093755