
Involvement of Vasoactive Intestinal Peptide Family Members in Diabetic Keratopathy





Abstract
1. Introduction
2. Overview of Diabetic Keratopathy
3. Expression of Neuropeptides in the Cornea
- Gs signaling is involved in adenylyl cyclase (AC) activity, which regulates intracellular adenosine 3′,5′-cyclic monophosphate (cAMP). cAMP is an intracellular signal transmitter that, in turn, acts as a second messenger and activator of cAMP-dependent protein kinase A (PKA).
- Gi/o signaling is involved in the inhibition of AC activity, resulting in decreased intracellular cAMP production.
- Gq/11 signaling activates phospholipase Cβ (PLCβ), which hydrolyzes phosphatidylinositol 4,5-bisphosphate (PIP2), releasing diacylglycerol (DAG) and 1,4,5-inositol trisphosphate (IP3). DAG activates protein kinase C (PKC), whereas IP3 diffuses to the endoplasmic reticulum (ER) and binds to IP3 receptors on ligand-gated calcium channels on the surface of ER leading to the release of calcium ions.
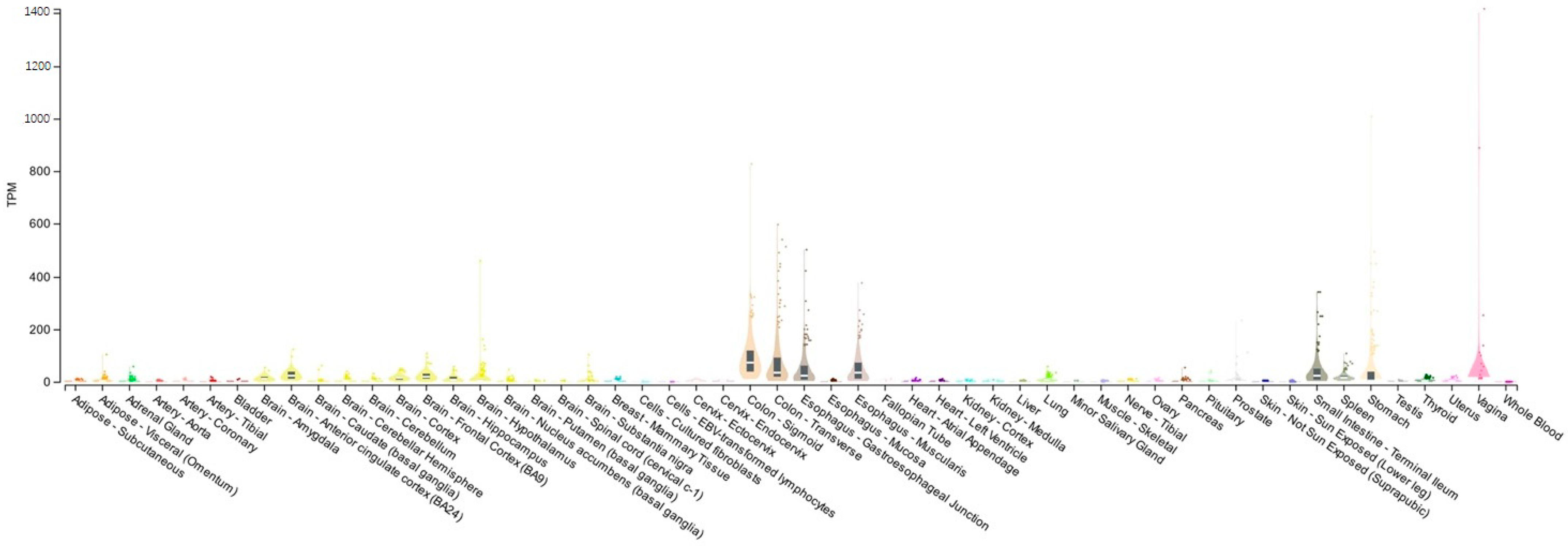
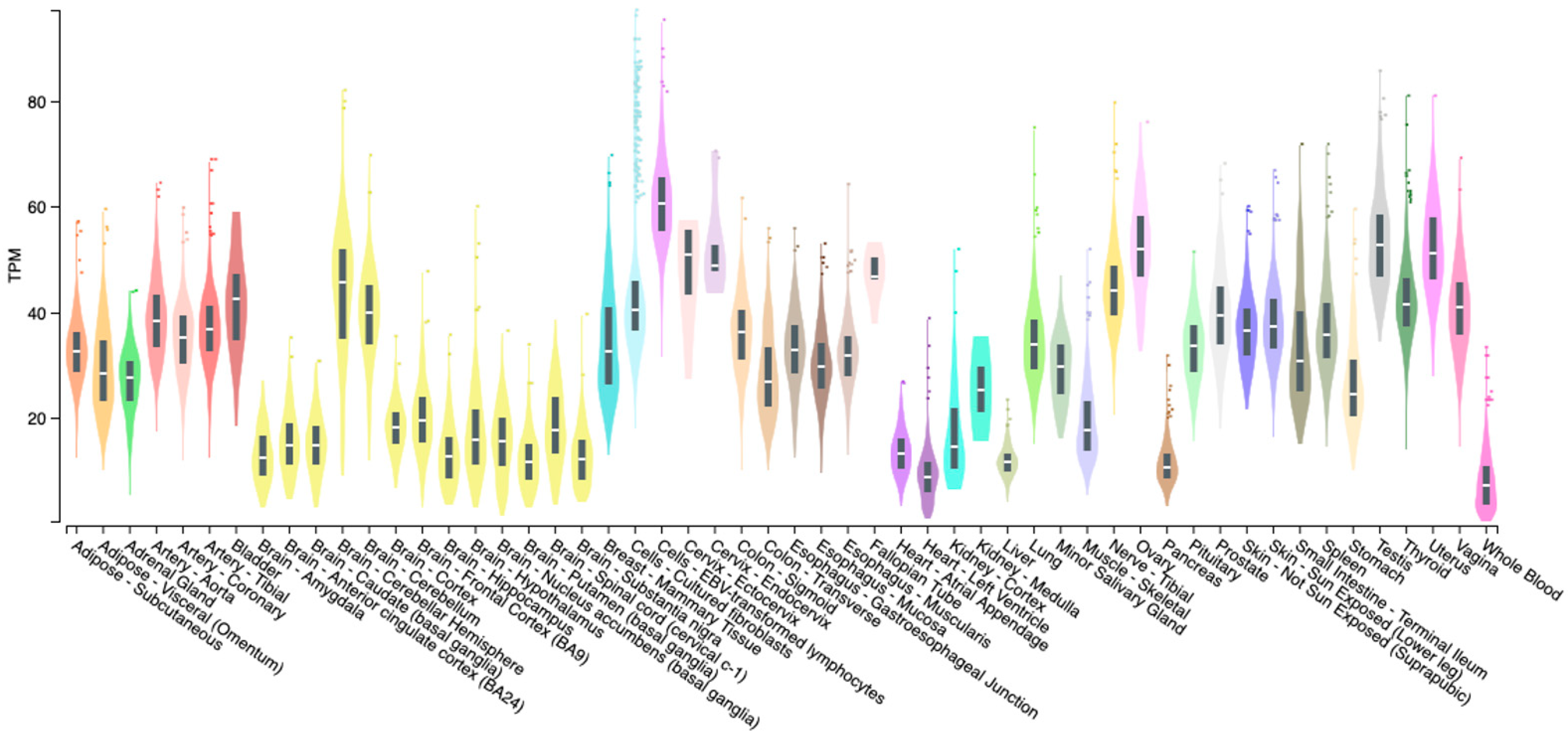
3.1. Vasoactive Intestinal Peptide (VIP)
3.2. Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP)
3.3. Activity-Dependent Neuroprotective Protein (ADNP)
4. Discussion
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef]
- Purushothaman, I.; Zagon, I.S.; Sassani, J.W.; McLaughlin, P.J. Ocular surface complications in diabetes: The interrelationship between insulin and enkephalin. Biochem. Pharmacol. 2021, 192, 114712. [Google Scholar] [CrossRef]
- Priyadarsini, S.; Whelchel, A.; Nicholas, S.; Sharif, R.; Riaz, K.; Karamichos, D. Diabetic keratopathy: Insights and challenges. Surv. Ophthalmol. 2020, 65, 513–529. [Google Scholar] [CrossRef]
- Singer, M.; O’Brien, P.; Mein, L.; Olvera, A. Corneal Sensitivity Is Inversely Correlated with Severity of Diabetic Retinopathy in a Predominantly Underrepresented Population. Am. J. Ophthalmol. 2023, 259, 53–61. [Google Scholar] [CrossRef]
- Salami, M.O.; Aribaba, O.T.; Musa, K.O.; Rotimi-Samuel, A.; Onakoya, A.O. Relationship between corneal sensitivity and diabetic retinopathy among diabetics attending a Nigerian Teaching Hospital. Int. Ophthalmol. 2020, 40, 2707–2716. [Google Scholar] [CrossRef] [PubMed]
- Tey, K.Y.; Teo, K.; Tan, A.C.S.; Devarajan, K.; Tan, B.; Tan, J.; Schmetterer, L.; Ang, M. Optical coherence tomography angiography in diabetic retinopathy: A review of current applications. Eye Vis. 2019, 6, 37. [Google Scholar] [CrossRef]
- D’Aloisio, R.; Giglio, R.; Di Nicola, M.; De Giacinto, C.; Pastore, M.R.; Tognetto, D.; Peto, T. Diagnostic Accuracy of Digital Retinal Fundus Image Analysis in Detecting Diabetic Maculopathy in Type 2 Diabetes Mellitus. Ophthalmic Res. 2019, 61, 100–106. [Google Scholar] [CrossRef]
- Ang, M.; Tan, A.C.S.; Cheung, C.M.G.; Keane, P.A.; Dolz-Marco, R.; Sng, C.C.A.; Schmetterer, L. Optical coherence tomography angiography: A review of current and future clinical applications. Graefes Arch. Clin. Exp. Ophthalmol. 2018, 256, 237–245. [Google Scholar] [CrossRef]
- Ferrara, M.; Loda, A.; Coco, G.; Grassi, P.; Cestaro, S.; Rezzola, S.; Romano, V.; Semeraro, F. Diabetic Retinopathy: Soluble and Imaging Ocular Biomarkers. J. Clin. Med. 2023, 12, 912. [Google Scholar] [CrossRef]
- Matulevičiūtė, I.; Sidaraitė, A.; Tatarūnas, V.; Veikutienė, A.; Dobilienė, O.; Žaliūnienė, D. Retinal and Choroidal Thinning-A Predictor of Coronary Artery Occlusion? Diagnostics 2022, 12, 2016. [Google Scholar] [CrossRef]
- Ong, C.J.T.; Wong, M.Y.Z.; Cheong, K.X.; Zhao, J.; Teo, K.Y.C.; Tan, T.E. Optical Coherence Tomography Angiography in Retinal Vascular Disorders. Diagnostics 2023, 13, 1620. [Google Scholar] [CrossRef] [PubMed]
- Palma, F.; Camacho, P. The role of Optical Coherence Tomography Angiography to detect early microvascular changes in Diabetic Retinopathy: A systematic review. J. Diabetes Metab. Disord. 2021, 20, 1957–1974. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.L.; Craig, J.P.; Patel, D.V.; McGhee, C.N.; Pradhan, M.; Ellyett, K.; Kilfoyle, D.; Braatvedt, G.D. In Vivo Confocal Microscopy of Corneal Nerves: An Ocular Biomarker for Peripheral and Cardiac Autonomic Neuropathy in Type 1 Diabetes Mellitus. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5060–5065. [Google Scholar] [CrossRef] [PubMed]
- Cosmo, E.; Midena, G.; Frizziero, L.; Bruno, M.; Cecere, M.; Midena, E. Corneal Confocal Microscopy as a Quantitative Imaging Biomarker of Diabetic Peripheral Neuropathy: A Review. J. Clin. Med. 2022, 11, 5130. [Google Scholar] [CrossRef]
- Mansoor, H.; Tan, H.C.; Lin, M.T.; Mehta, J.S.; Liu, Y.C. Diabetic Corneal Neuropathy. J. Clin. Med. 2020, 9, 3956. [Google Scholar] [CrossRef]
- Dewitt, E.N. The Histopathology of Bowman’s Membrane. Trans. Am. Ophthalmol. Soc. 1931, 29, 461–485. [Google Scholar]
- Bron, A.J. The architecture of the corneal stroma. Br. J. Ophthalmol. 2001, 85, 379–381. [Google Scholar] [CrossRef]
- Espana, E.M.; Birk, D.E. Composition, structure and function of the corneal stroma. Exp. Eye Res. 2020, 198, 108137. [Google Scholar] [CrossRef]
- Joyce, N.C. Proliferative capacity of the corneal endothelium. Prog. Retin. Eye Res. 2003, 22, 359–389. [Google Scholar] [CrossRef]
- Imanishi, J.; Kamiyama, K.; Iguchi, I.; Kita, M.; Sotozono, C.; Kinoshita, S. Growth factors: Importance in wound healing and maintenance of transparency of the cornea. Prog. Retin. Eye Res. 2000, 19, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Puri, S.; Kenyon, B.M.; Hamrah, P. Immunomodulatory Role of Neuropeptides in the Cornea. Biomedicines 2022, 10, 1985. [Google Scholar] [CrossRef]
- Giri, B.; Dey, S.; Das, T.; Sarkar, M.; Banerjee, J.; Dash, S.K. Chronic hyperglycemia mediated physiological alteration and metabolic distortion leads to organ dysfunction, infection, cancer progression and other pathophysiological consequences: An update on glucose toxicity. Biomed. Pharmacother. 2018, 107, 306–328. [Google Scholar] [CrossRef]
- Lane, J.D.; Krumholz, D.M.; Sack, R.A.; Morris, C. Tear glucose dynamics in diabetes mellitus. Curr. Eye Res. 2006, 31, 895–901. [Google Scholar] [CrossRef]
- Kim, J.; Kim, C.S.; Sohn, E.; Jeong, I.H.; Kim, H.; Kim, J.S. Involvement of advanced glycation end products, oxidative stress and nuclear factor-kappaB in the development of diabetic keratopathy. Graefes Arch. Clin. Exp. Ophthalmol. 2011, 249, 529–536. [Google Scholar] [CrossRef]
- Zhu, L.; Titone, R.; Robertson, D.M. The impact of hyperglycemia on the corneal epithelium: Molecular mechanisms and insight. Ocul. Surf. 2019, 17, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Lutty, G.A. Effects of diabetes on the eye. Investig. Ophthalmol. Vis. Sci. 2013, 54, Orsf81–Orsf87. [Google Scholar] [CrossRef]
- Yeung, A.; Dwarakanathan, S. Diabetic keratopathy. Dis. Mon. 2021, 67, 101135. [Google Scholar] [CrossRef]
- Shi, L.; Chen, H.; Yu, X.; Wu, X. Advanced glycation end products delay corneal epithelial wound healing through reactive oxygen species generation. Mol. Cell. Biochem. 2013, 383, 253–259. [Google Scholar] [CrossRef]
- Gekka, M.; Miyata, K.; Nagai, Y.; Nemoto, S.; Sameshima, T.; Tanabe, T.; Maruoka, S.; Nakahara, M.; Kato, S.; Amano, S. Corneal epithelial barrier function in diabetic patients. Cornea 2004, 23, 35–37. [Google Scholar] [CrossRef]
- Dan, J.; Zhou, Q.; Zhai, H.; Cheng, J.; Wan, L.; Ge, C.; Xie, L. Clinical analysis of fungal keratitis in patients with and without diabetes. PLoS ONE 2018, 13, e0196741. [Google Scholar] [CrossRef]
- Simpson, R.G.; Moshirfar, M.; Edmonds, J.N.; Christiansen, S.M. Laser in-situ keratomileusis in patients with diabetes mellitus: A review of the literature. Clin. Ophthalmol. 2012, 6, 1665–1674. [Google Scholar] [CrossRef] [PubMed]
- Javadi, M.A.; Zarei-Ghanavati, S. Cataracts in diabetic patients: A review article. J. Ophthalmic. Vis. Res. 2008, 3, 52–65. [Google Scholar] [PubMed]
- Sitompul, R. Corneal Sensitivity as a Potential Marker of Diabetic Neuropathy. Acta Med. Indones 2017, 49, 166–172. [Google Scholar] [PubMed]
- Müller, L.J.; Marfurt, C.F.; Kruse, F.; Tervo, T.M. Corneal nerves: Structure, contents and function. Exp. Eye Res. 2003, 76, 521–542. [Google Scholar] [CrossRef]
- Shaheen, B.S.; Bakir, M.; Jain, S. Corneal nerves in health and disease. Surv. Ophthalmol. 2014, 59, 263–285. [Google Scholar] [CrossRef]
- Labetoulle, M.; Baudouin, C.; Calonge, M.; Merayo-Lloves, J.; Boboridis, K.G.; Akova, Y.A.; Aragona, P.; Geerling, G.; Messmer, E.M.; Benítez-Del-Castillo, J. Role of corneal nerves in ocular surface homeostasis and disease. Acta Ophthalmol. 2019, 97, 137–145. [Google Scholar] [CrossRef]
- Zhou, T.; Lee, A.; Lo, A.C.Y.; Kwok, J. Diabetic Corneal Neuropathy: Pathogenic Mechanisms and Therapeutic Strategies. Front. Pharmacol. 2022, 13, 816062. [Google Scholar] [CrossRef]
- Bu, Y.; Shih, K.C.; Tong, L. The ocular surface and diabetes, the other 21st Century epidemic. Exp. Eye Res. 2022, 220, 109099. [Google Scholar] [CrossRef]
- Lewis, E.J.H.; Lovblom, L.E.; Ferdousi, M.; Halpern, E.M.; Jeziorska, M.; Pacaud, D.; Pritchard, N.; Dehghani, C.; Edwards, K.; Srinivasan, S.; et al. Rapid Corneal Nerve Fiber Loss: A Marker of Diabetic Neuropathy Onset and Progression. Diabetes Care 2020, 43, 1829–1835. [Google Scholar] [CrossRef]
- Zhou, Q.; Yang, L.; Wang, Q.; Li, Y.; Wei, C.; Xie, L. Mechanistic investigations of diabetic ocular surface diseases. Front. Endocrinol. 2022, 13, 1079541. [Google Scholar] [CrossRef]
- Zou, C.; Wang, S.; Huang, F.; Zhang, Y.A. Advanced glycation end products and ultrastructural changes in corneas of long-term streptozotocin-induced diabetic monkeys. Cornea 2012, 31, 1455–1459. [Google Scholar] [CrossRef]
- Inoue, K.; Kato, S.; Inoue, Y.; Amano, S.; Oshika, T. The corneal endothelium and thickness in type II diabetes mellitus. Jpn. J. Ophthalmol. 2002, 46, 65–69. [Google Scholar] [CrossRef]
- Abdelkader, H.; Patel, D.V.; McGhee, C.; Alany, R.G. New therapeutic approaches in the treatment of diabetic keratopathy: A review. Clin. Exp. Ophthalmol. 2011, 39, 259–270. [Google Scholar] [CrossRef]
- Strand, F.L. Neuropeptides: General characteristics and neuropharmaceutical potential in treating CNS disorders. Prog. Drug Res. 2003, 61, 1–37. [Google Scholar] [CrossRef]
- Amram, N.; Hacohen-Kleiman, G.; Sragovich, S.; Malishkevich, A.; Katz, J.; Touloumi, O.; Lagoudaki, R.; Grigoriadis, N.C.; Giladi, E.; Yeheskel, A.; et al. Sexual divergence in microtubule function: The novel intranasal microtubule targeting SKIP normalizes axonal transport and enhances memory. Mol. Psychiatry 2016, 21, 1467–1476. [Google Scholar] [CrossRef]
- Li, C.; Kim, K. Neuropeptides. WormBook 2008, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Koh, S.W.; Waschek, J.A. Corneal endothelial cell survival in organ cultures under acute oxidative stress: Effect of VIP. Investig. Ophthalmol. Vis. Sci. 2000, 41, 4085–4092. [Google Scholar]
- Sacchetti, M.; Lambiase, A. Neurotrophic factors and corneal nerve regeneration. Neural Regen. Res. 2017, 12, 1220–1224. [Google Scholar] [CrossRef]
- Koh, S.W. Corneal endothelial autocrine trophic factor VIP in a mechanism-based strategy to enhance human donor cornea preservation for transplantation. Exp. Eye Res. 2012, 95, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Alm, P.; Håkanson, R. Distribution and effects of pituitary adenylate cyclase-activating peptide in the rabbit eye. Neuroscience 1995, 69, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Giunta, S.; Giallongo, C.; Tibullo, D.; Bucolo, C.; Saccone, S.; Federico, C.; Scollo, D.; Longo, A.; et al. Activity-Dependent Neuroprotective Protein (ADNP)-Derived Peptide (NAP) Counteracts UV-B Radiation-Induced ROS Formation in Corneal Epithelium. Antioxidants 2022, 11, 128. [Google Scholar] [CrossRef]
- Beckers, H.J.; Klooster, J.; Vrensen, G.F.; Lamers, W.P. Substance P in rat corneal and iridal nerves: An ultrastructural immunohistochemical study. Ophthalmic Res. 1993, 25, 192–200. [Google Scholar] [CrossRef]
- Watanabe, M.; Nakayasu, K.; Iwatsu, M.; Kanai, A. Endogenous substance P in corneal epithelial cells and keratocytes. Jpn. J. Ophthalmol. 2002, 46, 616–620. [Google Scholar] [CrossRef]
- Lambiase, A.; Micera, A.; Sacchetti, M.; Cortes, M.; Mantelli, F.; Bonini, S. Alterations of tear neuromediators in dry eye disease. Arch Ophthalmol. 2011, 129, 981–986. [Google Scholar] [CrossRef]
- Jones, M.A.; Marfurt, C.F. Peptidergic innervation of the rat cornea. Exp. Eye Res. 1998, 66, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Kangawa, K.; Eto, T. Adrenomedullin and PAMP: Discovery, structures, and cardiovascular functions. Microsc. Res. Tech. 2002, 57, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Słoniecka, M.; Le Roux, S.; Boman, P.; Byström, B.; Zhou, Q.; Danielson, P. Expression Profiles of Neuropeptides, Neurotransmitters, and Their Receptors in Human Keratocytes In Vitro and In Situ. PLoS ONE 2015, 10, e0134157. [Google Scholar] [CrossRef] [PubMed]
- Patel, Y.C. Molecular pharmacology of somatostatin receptor subtypes. J. Endocrinol. Investig. 1997, 20, 348–367. [Google Scholar] [CrossRef] [PubMed]
- Tinsley, P.W.; Fridland, G.H.; Killmar, J.T.; Desiderio, D.M. Purification, characterization, and localization of neuropeptides in the cornea. Peptides 1988, 9, 1373–1379. [Google Scholar] [CrossRef]
- Schrödl, F.; Kaser-Eichberger, A.; Trost, A.; Strohmaier, C.; Bogner, B.; Runge, C.; Bruckner, D.; Motloch, K.; Holub, B.; Kofler, B.; et al. Distribution of galanin receptors in the human eye. Exp. Eye Res. 2015, 138, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Bourcier, T.; Rondeau, N.; Paquet, S.; Forgez, P.; Lombet, A.; Pouzaud, F.; Rostène, W.; Borderie, V.; Laroche, L. Expression of neurotensin receptors in human corneal keratocytes. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1765–1771. [Google Scholar]
- Lambiase, A.; Manni, L.; Bonini, S.; Rama, P.; Micera, A.; Aloe, L. Nerve growth factor promotes corneal healing: Structural, biochemical, and molecular analyses of rat and human corneas. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1063–1069. [Google Scholar]
- Zagon, I.S.; Verderame, M.F.; McLaughlin, P.J. The biology of the opioid growth factor receptor (OGFr). Brain Res. Brain Res. Rev. 2002, 38, 351–376. [Google Scholar] [CrossRef] [PubMed]
- Zagon, I.S.; Sassani, J.W.; Allison, G.; McLaughlin, P.J. Conserved expression of the opioid growth factor, [Met5]enkephalin, and the zeta (zeta) opioid receptor in vertebrate cornea. Brain Res. 1995, 671, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Said, S.I.; Mutt, V. Polypeptide with broad biological activity: Isolation from small intestine. Science 1970, 169, 1217–1218. [Google Scholar] [CrossRef] [PubMed]
- Henning, R.J.; Sawmiller, D.R. Vasoactive intestinal peptide: Cardiovascular effects. Cardiovasc. Res. 2001, 49, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Couvineau, A.; Laburthe, M. VPAC receptors: Structure, molecular pharmacology and interaction with accessory proteins. Br. J. Pharmacol. 2012, 166, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Laburthe, M.; Couvineau, A.; Tan, V. Class II G protein-coupled receptors for VIP and PACAP: Structure, models of activation and pharmacology. Peptides 2007, 28, 1631–1639. [Google Scholar] [CrossRef]
- Winzell, M.S.; Ahrén, B. Role of VIP and PACAP in islet function. Peptides 2007, 28, 1805–1813. [Google Scholar] [CrossRef]
- Fabricius, D.; Karacay, B.; Shutt, D.; Leverich, W.; Schafer, B.; Takle, E.; Thedens, D.; Khanna, G.; Raikwar, S.; Yang, B.; et al. Characterization of intestinal and pancreatic dysfunction in VPAC1-null mutant mouse. Pancreas 2011, 40, 861–871. [Google Scholar] [CrossRef]
- Bertrand, G.; Puech, R.; Maisonnasse, Y.; Bockaert, J.; Loubatières-Mariani, M.M. Comparative effects of PACAP and VIP on pancreatic endocrine secretions and vascular resistance in rat. Br. J. Pharmacol. 1996, 117, 764–770. [Google Scholar] [CrossRef]
- Hou, X.; Yang, D.; Yang, G.; Li, M.; Zhang, J.; Zhang, J.; Zhang, Y.; Liu, Y. Therapeutic potential of vasoactive intestinal peptide and its receptor VPAC2 in type 2 diabetes. Front. Endocrinol. 2022, 13, 984198. [Google Scholar] [CrossRef]
- Vu, J.P.; Larauche, M.; Flores, M.; Luong, L.; Norris, J.; Oh, S.; Liang, L.J.; Waschek, J.; Pisegna, J.R.; Germano, P.M. Regulation of Appetite, Body Composition, and Metabolic Hormones by Vasoactive Intestinal Polypeptide (VIP). J. Mol. Neurosci. 2015, 56, 377–387. [Google Scholar] [CrossRef]
- Gozes, I. VIP, from gene to behavior and back: Summarizing my 25 years of research. J. Mol. Neurosci. 2008, 36, 115–124. [Google Scholar] [CrossRef]
- Hill, J.M.; Hauser, J.M.; Sheppard, L.M.; Abebe, D.; Spivak-Pohis, I.; Kushnir, M.; Deitch, I.; Gozes, I. Blockage of VIP during mouse embryogenesis modifies adult behavior and results in permanent changes in brain chemistry. J. Mol. Neurosci. 2007, 31, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Said, S.I.; Mutt, V. Potent peripheral and splanchnic vasodilator peptide from normal gut. Nature 1970, 225, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Morell, M.; Souza-Moreira, L.; González-Rey, E. VIP in neurological diseases: More than a neuropeptide. Endocr. Metab. Immune Disord. Drug Targets 2012, 12, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Moody, T.W.; Nuche-Berenguer, B.; Jensen, R.T. Vasoactive intestinal peptide/pituitary adenylate cyclase activating polypeptide, and their receptors and cancer. Curr. Opin. Endocrinol. Diabetes Obes. 2016, 23, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Moody, T.W.; Gozes, I. Vasoactive intestinal peptide receptors: A molecular target in breast and lung cancer. Curr. Pharm. Des. 2007, 13, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Rasà, D.M.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. PACAP and VIP regulate hypoxia-inducible factors in neuroblastoma cells exposed to hypoxia. Neuropeptides 2018, 69, 84–91. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.G.; Maugeri, G.; Rasà, D.M.; Reitano, R.; Saccone, S.; Federico, C.; Magro, G.; D’Agata, V. Modulatory role of PACAP and VIP on HIFs expression in lung adenocarcinoma. Peptides 2021, 146, 170672. [Google Scholar] [CrossRef]
- Gonzalez-Rey, E.; Chorny, A.; Fernandez-Martin, A.; Ganea, D.; Delgado, M. Vasoactive intestinal peptide generates human tolerogenic dendritic cells that induce CD4 and CD8 regulatory T cells. Blood 2006, 107, 3632–3638. [Google Scholar] [CrossRef]
- Delgado, M.; Gonzalez-Rey, E.; Ganea, D. The neuropeptide vasoactive intestinal peptide generates tolerogenic dendritic cells. J. Immunol. 2005, 175, 7311–7324. [Google Scholar] [CrossRef] [PubMed]
- Dragich, J.M.; Loh, D.H.; Wang, L.M.; Vosko, A.M.; Kudo, T.; Nakamura, T.J.; Odom, I.H.; Tateyama, S.; Hagopian, A.; Waschek, J.A.; et al. The role of the neuropeptides PACAP and VIP in the photic regulation of gene expression in the suprachiasmatic nucleus. Eur. J. Neurosci. 2010, 31, 864–875. [Google Scholar] [CrossRef] [PubMed]
- Akrouh, A.; Kerschensteiner, D. Morphology and function of three VIP-expressing amacrine cell types in the mouse retina. J. Neurophysiol. 2015, 114, 2431–2438. [Google Scholar] [CrossRef] [PubMed]
- Tunçel, N.; Başmak, H.; Uzuner, K.; Tunçel, M.; Altiokka, G.; Zaimoğlu, V.; Ozer, A.; Gürer, F. Protection of rat retina from ischemia-reperfusion injury by vasoactive intestinal peptide (VIP): The effect of VIP on lipid peroxidation and antioxidant enzyme activity of retina and choroid. Ann. N. Y. Acad. Sci. 1996, 805, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Szabadfi, K.; Danyadi, B.; Kiss, P.; Tamas, A.; Fabian, E.; Gabriel, R.; Reglodi, D. Protective effects of vasoactive intestinal peptide (VIP) in ischemic retinal degeneration. J. Mol. Neurosci. 2012, 48, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Carion, T.W.; Jiang, Y.; Steinle, J.J.; Berger, E.A. VIP protects human retinal microvascular endothelial cells against high glucose-induced increases in TNF-α and enhances RvD1. Prostaglandins Other Lipid Mediat. 2016, 123, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. PACAP and VIP Inhibit HIF-1α-Mediated VEGF Expression in a Model of Diabetic Macular Edema. J. Cell. Physiol. 2017, 232, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Gagliano, C.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. VIP Family Members Prevent Outer Blood Retinal Barrier Damage in a Model of Diabetic Macular Edema. J. Cell. Physiol. 2017, 232, 1079–1085. [Google Scholar] [CrossRef]
- Troger, J.; Neyer, S.; Heufler, C.; Huemer, H.; Schmid, E.; Griesser, U.; Kralinger, M.; Kremser, B.; Baldissera, I.; Kieselbach, G. Substance P and vasoactive intestinal polypeptide in the streptozotocin-induced diabetic rat retina. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1045–1050. [Google Scholar]
- Taylor, A.W.; Streilein, J.W.; Cousins, S.W. Immunoreactive vasoactive intestinal peptide contributes to the immunosuppressive activity of normal aqueous humor. J. Immunol. 1994, 153, 1080–1086. [Google Scholar] [CrossRef]
- Sacchetti, M.; Micera, A.; Lambiase, A.; Speranza, S.; Mantelli, F.; Petrachi, G.; Bonini, S.; Bonini, S. Tear levels of neuropeptides increase after specific allergen challenge in allergic conjunctivitis. Mol. Vis. 2011, 17, 47–52. [Google Scholar]
- Berger, E.A.; Vistisen, K.S.; Barrett, R.P.; Hazlett, L.D. Effects of VIP on corneal reconstitution and homeostasis following Pseudomonas aeruginosa induced keratitis. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7432–7439. [Google Scholar] [CrossRef] [PubMed]
- Koh, S.M.; Coll, T.; Gloria, D.; Sprehe, N. Corneal Endothelial Cell Integrity in Precut Human Donor Corneas Enhanced by Autocrine Vasoactive Intestinal Peptide. Cornea 2017, 36, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Szliter, E.A.; Lighvani, S.; Barrett, R.P.; Hazlett, L.D. Vasoactive intestinal peptide balances pro- and anti-inflammatory cytokines in the Pseudomonas aeruginosa-infected cornea and protects against corneal perforation. J. Immunol. 2007, 178, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; McClellan, S.A.; Barrett, R.P.; Zhang, Y.; Hazlett, L.D. Vasoactive intestinal peptide downregulates proinflammatory TLRs while upregulating anti-inflammatory TLRs in the infected cornea. J. Immunol. 2012, 189, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef] [PubMed]
- Arimura, A.; Somogyvári-Vigh, A.; Miyata, A.; Mizuno, K.; Coy, D.H.; Kitada, C. Tissue distribution of PACAP as determined by RIA: Highly abundant in the rat brain and testes. Endocrinology 1991, 129, 2787–2789. [Google Scholar] [CrossRef] [PubMed]
- Arimura, A.; Shioda, S. Pituitary adenylate cyclase activating polypeptide (PACAP) and its receptors: Neuroendocrine and endocrine interaction. Front. Neuroendocrinol. 1995, 16, 53–88. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, T.; Grimaldi, M.; Eiden, L.E. The hop cassette of the PAC1 receptor confers coupling to Ca2+ elevation required for pituitary adenylate cyclase-activating polypeptide-evoked neurosecretion. J. Biol. Chem. 2007, 282, 8079–8091. [Google Scholar] [CrossRef] [PubMed]
- Dickson, L.; Finlayson, K. VPAC and PAC receptors: From ligands to function. Pharmacol. Ther. 2009, 121, 294–316. [Google Scholar] [CrossRef] [PubMed]
- Fabian, E.; Reglodi, D.; Mester, L.; Szabo, A.; Szabadfi, K.; Tamas, A.; Toth, G.; Kovacs, K. Effects of PACAP on intracellular signaling pathways in human retinal pigment epithelial cells exposed to oxidative stress. J. Mol. Neurosci. 2012, 48, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Bassan, M.; Zamostiano, R.; Davidson, A.; Pinhasov, A.; Giladi, E.; Perl, O.; Bassan, H.; Blat, C.; Gibney, G.; Glazner, G.; et al. Complete sequence of a novel protein containing a femtomolar-activity-dependent neuroprotective peptide. J. Neurochem. 1999, 72, 1283–1293. [Google Scholar] [CrossRef]
- Zamostiano, R.; Pinhasov, A.; Gelber, E.; Steingart, R.A.; Seroussi, E.; Giladi, E.; Bassan, M.; Wollman, Y.; Eyre, H.J.; Mulley, J.C.; et al. Cloning and characterization of the human activity-dependent neuroprotective protein. J. Biol. Chem. 2001, 276, 708–714. [Google Scholar] [CrossRef]
- Magrì, B.; D’Amico, A.G.; Maugeri, G.; Morello, G.; La Cognata, V.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. Neuroprotective effect of the PACAP-ADNP axis on SOD1G93A mutant motor neuron death induced by trophic factors deprivation. Neuropeptides 2023, 102, 102386. [Google Scholar] [CrossRef]
- D’Amico, A.G.; Maugeri, G.; Musumeci, G.; Reglodi, D.; D’Agata, V. PACAP and NAP: Effect of Two Functionally Related Peptides in Diabetic Retinopathy. J. Mol. Neurosci. 2021, 71, 1525–1535. [Google Scholar] [CrossRef]
- Moody, T.W.; Ramos-Alvarez, I.; Jensen, R.T. Bombesin, endothelin, neurotensin and pituitary adenylate cyclase activating polypeptide cause tyrosine phosphorylation of receptor tyrosine kinases. Peptides 2021, 137, 170480. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Reitano, R.; Magro, G.; Cavallaro, S.; Salomone, S.; D’Agata, V. PACAP and VIP Inhibit the Invasiveness of Glioblastoma Cells Exposed to Hypoxia through the Regulation of HIFs and EGFR Expression. Front. Pharmacol. 2016, 7, 139. [Google Scholar] [CrossRef]
- Moody, T.W.; Lee, L.; Jensen, R.T. The G Protein-Coupled Receptor PAC1 Regulates Transactivation of the Receptor Tyrosine Kinase HER3. J. Mol. Neurosci. 2021, 71, 1589–1597. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Bucolo, C.; D’Agata, V. Protective effect of PACAP-38 on retinal pigmented epithelium in an in vitro and in vivo model of diabetic retinopathy through EGFR-dependent mechanism. Peptides 2019, 119, 170108. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Rasà, D.M.; Federico, C.; Saccone, S.; Morello, G.; La Cognata, V.; Cavallaro, S.; D’Agata, V. Molecular mechanisms involved in the protective effect of pituitary adenylate cyclase-activating polypeptide in an in vitro model of amyotrophic lateral sclerosis. J. Cell. Physiol. 2019, 234, 5203–5214. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Castrogiovanni, P.; Saccone, S.; Federico, C.; Reibaldi, M.; Russo, A.; Bonfiglio, V.; Avitabile, T.; Longo, A.; et al. PACAP through EGFR transactivation preserves human corneal endothelial integrity. J. Cell. Biochem. 2019, 120, 10097–10105. [Google Scholar] [CrossRef]
- Farkas, J.; Kovács, L.; Gáspár, L.; Nafz, A.; Gaszner, T.; Ujvári, B.; Kormos, V.; Csernus, V.; Hashimoto, H.; Reglődi, D.; et al. Construct and face validity of a new model for the three-hit theory of depression using PACAP mutant mice on CD1 background. Neuroscience 2017, 354, 11–29. [Google Scholar] [CrossRef]
- Gupta, A.; Gargiulo, A.T.; Curtis, G.R.; Badve, P.S.; Pandey, S.; Barson, J.R. Pituitary Adenylate Cyclase-Activating Polypeptide-27 (PACAP-27) in the Thalamic Paraventricular Nucleus Is Stimulated by Ethanol Drinking. Alcohol. Clin. Exp. Res. 2018, 42, 1650–1660. [Google Scholar] [CrossRef]
- Han, P.; Tang, Z.; Yin, J.; Maalouf, M.; Beach, T.G.; Reiman, E.M.; Shi, J. Pituitary adenylate cyclase-activating polypeptide protects against β-amyloid toxicity. Neurobiol. Aging 2014, 35, 2064–2071. [Google Scholar] [CrossRef]
- King, S.B.; Lezak, K.R.; O’Reilly, M.; Toufexis, D.J.; Falls, W.A.; Braas, K.; May, V.; Hammack, S.E. The Effects of Prior Stress on Anxiety-Like Responding to Intra-BNST Pituitary Adenylate Cyclase Activating Polypeptide in Male and Female Rats. Neuropsychopharmacology 2017, 42, 1679–1687. [Google Scholar] [CrossRef][Green Version]
- Heppner, T.J.; Hennig, G.W.; Nelson, M.T.; May, V.; Vizzard, M.A. PACAP38-Mediated Bladder Afferent Nerve Activity Hyperexcitability and Ca(2+) Activity in Urothelial Cells from Mice. J. Mol. Neurosci. 2019, 68, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Parsons, R.L.; May, V. PACAP-Induced PAC1 Receptor Internalization and Recruitment of Endosomal Signaling Regulate Cardiac Neuron Excitability. J. Mol. Neurosci. 2019, 68, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Reglodi, D.; Illes, A.; Opper, B.; Schafer, E.; Tamas, A.; Horvath, G. Presence and Effects of Pituitary Adenylate Cyclase Activating Polypeptide Under Physiological and Pathological Conditions in the Stomach. Front. Endocrinol. 2018, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Bardosi, S.; Bardosi, A.; Nagy, Z.; Reglodi, D. Expression of PACAP and PAC1 Receptor in Normal Human Thyroid Gland and in Thyroid Papillary Carcinoma. J. Mol. Neurosci. 2016, 60, 171–178. [Google Scholar] [CrossRef]
- Egri, P.; Fekete, C.; Dénes, Á.; Reglődi, D.; Hashimoto, H.; Fülöp, B.D.; Gereben, B. Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) Regulates the Hypothalamo-Pituitary-Thyroid (HPT) Axis via Type 2 Deiodinase in Male Mice. Endocrinology 2016, 157, 2356–2366. [Google Scholar] [CrossRef]
- Prevost, G.; Arabo, A.; Jian, L.; Quelennec, E.; Cartier, D.; Hassan, S.; Falluel-Morel, A.; Tanguy, Y.; Gargani, S.; Lihrmann, I.; et al. The PACAP-regulated gene selenoprotein T is abundantly expressed in mouse and human β-cells and its targeted inactivation impairs glucose tolerance. Endocrinology 2013, 154, 3796–3806. [Google Scholar] [CrossRef]
- Sasaki, S.; Watanabe, J.; Ohtaki, H.; Matsumoto, M.; Murai, N.; Nakamachi, T.; Hannibal, J.; Fahrenkrug, J.; Hashimoto, H.; Watanabe, H.; et al. Pituitary adenylate cyclase-activating polypeptide promotes eccrine gland sweat secretion. Br. J. Dermatol. 2017, 176, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Lajko, A.; Meggyes, M.; Fulop, B.D.; Gede, N.; Reglodi, D.; Szereday, L. Comparative analysis of decidual and peripheral immune cells and immune-checkpoint molecules during pregnancy in wild-type and PACAP-deficient mice. Am. J. Reprod. Immunol. 2018, 80, e13035. [Google Scholar] [CrossRef]
- Reglodi, D.; Tamas, A.; Koppan, M.; Szogyi, D.; Welke, L. Role of PACAP in Female Fertility and Reproduction at Gonadal Level-Recent Advances. Front. Endocrinol. 2012, 3, 155. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.A.; Leon, S.; Madara, J.C.; Schafer, D.; Fergani, C.; Maguire, C.A.; Verstegen, A.M.; Brengle, E.; Kong, D.; Herbison, A.E.; et al. PACAP neurons in the ventral premammillary nucleus regulate reproductive function in the female mouse. Elife 2018, 7, e35960. [Google Scholar] [CrossRef]
- Fulop, B.D.; Sandor, B.; Szentleleky, E.; Karanyicz, E.; Reglodi, D.; Gaszner, B.; Zakany, R.; Hashimoto, H.; Juhasz, T.; Tamas, A. Altered Notch Signaling in Developing Molar Teeth of Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP)-Deficient Mice. J. Mol. Neurosci. 2019, 68, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Reglodi, D.; Jungling, A.; Longuespée, R.; Kriegsmann, J.; Casadonte, R.; Kriegsmann, M.; Juhasz, T.; Bardosi, S.; Tamas, A.; Fulop, B.D.; et al. Accelerated pre-senile systemic amyloidosis in PACAP knockout mice—A protective role of PACAP in age-related degenerative processes. J. Pathol. 2018, 245, 478–490. [Google Scholar] [CrossRef]
- Watanabe, J.; Nakamachi, T.; Matsuno, R.; Hayashi, D.; Nakamura, M.; Kikuyama, S.; Nakajo, S.; Shioda, S. Localization, characterization and function of pituitary adenylate cyclase-activating polypeptide during brain development. Peptides 2007, 28, 1713–1719. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Musumeci, G.; Reglodi, D.; D’Agata, V. Effects of Pacap on Schwann Cells: Focus on Nerve Injury. Int. J. Mol. Sci. 2020, 21, 8233. [Google Scholar] [CrossRef]
- D’Agata, V.; Cavallaro, S. Functional and molecular expression of PACAP/VIP receptors in the rat retina. Brain Res. Mol. Brain Res. 1998, 54, 161–164. [Google Scholar] [CrossRef]
- Patko, E.; Szabo, E.; Toth, D.; Tornoczky, T.; Bosnyak, I.; Vaczy, A.; Atlasz, T.; Reglodi, D. Distribution of PACAP and PAC1 Receptor in the Human Eye. J. Mol. Neurosci. 2022, 72, 2176–2187. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Shioda, S.; Ogino, D.; Nakai, Y.; Arimura, A.; Koide, R. Distribution and ultrastructural localization of a receptor for pituitary adenylate cyclase activating polypeptide and its mRNA in the rat retina. Neurosci. Lett. 1997, 238, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Izumi, S.; Shioda, S.; Zhou, C.J.; Arimura, A.; Koide, R. Gene expression for PACAP receptor mRNA in the rat retina by in situ hybridization and in situ RT-PCR. Ann. N. Y. Acad. Sci. 2000, 921, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Nakamachi, T.; Matkovits, A.; Seki, T.; Shioda, S. Distribution and protective function of pituitary adenylate cyclase-activating polypeptide in the retina. Front. Endocrinol. 2012, 3, 145. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.G.; Maugeri, G.; Magrì, B.; Lombardo, C.; Saccone, S.; Federico, C.; Cavallaro, P.; Giunta, S.; Bucolo, C.; D’Agata, V. Pacap-adnp axis prevents outer retinal barrier breakdown and choroidal neovascularization by interfering with vegf secreted from retinal pigmented epitelium cells. Peptides 2023, 168, 171065. [Google Scholar] [CrossRef] [PubMed]
- Elsås, T.; Uddman, R.; Sundler, F. Pituitary adenylate cyclase-activating peptide-immunoreactive nerve fibers in the cat eye. Graefes Arch. Clin. Exp. Ophthalmol. 1996, 234, 573–580. [Google Scholar] [CrossRef]
- Maugeri, G.; Longo, A.; D’Amico, A.G.; Rasà, D.M.; Reibaldi, M.; Russo, A.; Bonfiglio, V.; Avitabile, T.; D’Agata, V. Trophic effect of PACAP on human corneal endothelium. Peptides 2018, 99, 20–26. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Amenta, A.; Saccone, S.; Federico, C.; Reibaldi, M.; Russo, A.; Bonfiglio, V.; Avitabile, T.; Longo, A.; et al. Protective effect of PACAP against ultraviolet B radiation-induced human corneal endothelial cell injury. Neuropeptides 2020, 79, 101978. [Google Scholar] [CrossRef]
- Fukiage, C.; Nakajima, T.; Takayama, Y.; Minagawa, Y.; Shearer, T.R.; Azuma, M. PACAP induces neurite outgrowth in cultured trigeminal ganglion cells and recovery of corneal sensitivity after flap surgery in rabbits. Am. J. Ophthalmol. 2007, 143, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, J.; Chen, X.; Hong, A. Expression, identification and biological effects of the novel recombination protein, PACAP38-NtA, with high bioactivity. Int. J. Mol. Med. 2015, 35, 376–382. [Google Scholar] [CrossRef]
- Shioda, S.; Takenoya, F.; Hirabayashi, T.; Wada, N.; Seki, T.; Nonaka, N.; Nakamachi, T. Effects of PACAP on Dry Eye Symptoms, and Possible Use for Therapeutic Application. J. Mol. Neurosci. 2019, 68, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Nakamachi, T.; Ohtaki, H.; Seki, T.; Yofu, S.; Kagami, N.; Hashimoto, H.; Shintani, N.; Baba, A.; Mark, L.; Lanekoff, I.; et al. PACAP suppresses dry eye signs by stimulating tear secretion. Nat. Commun. 2016, 7, 12034. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Waldeck, K.; Grundemar, L.; Håkanson, R. Ocular inflammation induced by electroconvulsive treatment: Contribution of nitric oxide and neuropeptides mobilized from C-fibres. Br. J. Pharmacol. 1997, 120, 1491–1496. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Alm, P.; Håkanson, R. PACAP occurs in sensory nerve fibers and participates in ocular inflammation in the rabbit. Ann. N. Y. Acad. Sci. 1996, 805, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Danielsen, N.; Sundler, F.; Mulder, H. Pituitary adenylate cyclase-activating peptide is upregulated in sensory neurons by inflammation. Neuroreport 1998, 9, 2833–2836. [Google Scholar] [CrossRef]
- Hashimoto, H.; Shintani, N.; Baba, A. New insights into the central PACAPergic system from the phenotypes in PACAP- and PACAP receptor-knockout mice. Ann. N. Y. Acad. Sci. 2006, 1070, 75–89. [Google Scholar] [CrossRef]
- Mandel, S.; Gozes, I. Activity-dependent neuroprotective protein constitutes a novel element in the SWI/SNF chromatin remodeling complex. J. Biol. Chem. 2007, 282, 34448–34456. [Google Scholar] [CrossRef]
- Gozes, I. The ADNP Syndrome and CP201 (NAP) Potential and Hope. Front. Neurol. 2020, 11, 608444. [Google Scholar] [CrossRef]
- Mandel, S.; Rechavi, G.; Gozes, I. Activity-dependent neuroprotective protein (ADNP) differentially interacts with chromatin to regulate genes essential for embryogenesis. Dev. Biol. 2007, 303, 814–824. [Google Scholar] [CrossRef]
- Helsmoortel, C.; Vulto-van Silfhout, A.T.; Coe, B.P.; Vandeweyer, G.; Rooms, L.; van den Ende, J.; Schuurs-Hoeijmakers, J.H.; Marcelis, C.L.; Willemsen, M.H.; Vissers, L.E.; et al. A SWI/SNF-related autism syndrome caused by de novo mutations in ADNP. Nat. Genet. 2014, 46, 380–384. [Google Scholar] [CrossRef]
- Pascolini, G.; Agolini, E.; Majore, S.; Novelli, A.; Grammatico, P.; Digilio, M.C. Helsmoortel-Van der Aa Syndrome as emerging clinical diagnosis in intellectually disabled children with autistic traits and ocular involvement. Eur. J. Paediatr. Neurol. 2018, 22, 552–557. [Google Scholar] [CrossRef]
- Levine, J.; Cohen, D.; Herman, C.; Verloes, A.; Guinchat, V.; Diaz, L.; Cravero, C.; Mandel, A.; Gozes, I. Developmental Phenotype of the Rare Case of DJ Caused by a Unique ADNP Gene De Novo Mutation. J. Mol. Neurosci. 2019, 68, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Gale, M.J.; Titus, H.E.; Harman, G.A.; Alabduljalil, T.; Dennis, A.; Wilson, J.L.; Koeller, D.M.; Finanger, E.; Blasco, P.A.; Chiang, P.W.; et al. Longitudinal ophthalmic findings in a child with Helsmoortel-Van der Aa Syndrome. Am. J. Ophthalmol. Case Rep. 2018, 10, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Gozes, I.; Ivashko-Pachima, Y. ADNP: In search for molecular mechanisms and innovative therapeutic strategies for frontotemporal degeneration. Front. Aging Neurosci. 2015, 7, 205. [Google Scholar] [CrossRef] [PubMed]
- Magen, I.; Gozes, I. Davunetide: Peptide therapeutic in neurological disorders. Curr. Med. Chem. 2014, 21, 2591–2598. [Google Scholar] [CrossRef]
- Oz, S.; Kapitansky, O.; Ivashco-Pachima, Y.; Malishkevich, A.; Giladi, E.; Skalka, N.; Rosin-Arbesfeld, R.; Mittelman, L.; Segev, O.; Hirsch, J.A.; et al. The NAP motif of activity-dependent neuroprotective protein (ADNP) regulates dendritic spines through microtubule end binding proteins. Mol. Psychiatry 2014, 19, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Sayas, C.L.; Tortosa, E.; Bollati, F.; Ramírez-Ríos, S.; Arnal, I.; Avila, J. Tau regulates the localization and function of End-binding proteins 1 and 3 in developing neuronal cells. J. Neurochem. 2015, 133, 653–667. [Google Scholar] [CrossRef]
- Ivashko-Pachima, Y.; Sayas, C.L.; Malishkevich, A.; Gozes, I. ADNP/NAP dramatically increase microtubule end-binding protein-Tau interaction: A novel avenue for protection against tauopathy. Mol. Psychiatry 2017, 22, 1335–1344. [Google Scholar] [CrossRef]
- D’Incal, C.P.; Van Rossem, K.E.; De Man, K.; Konings, A.; Van Dijck, A.; Rizzuti, L.; Vitriolo, A.; Testa, G.; Gozes, I.; Vanden Berghe, W.; et al. Chromatin remodeler Activity-Dependent Neuroprotective Protein (ADNP) contributes to syndromic autism. Clin. Epigenetics 2023, 15, 45. [Google Scholar] [CrossRef]
- Teuchner, B.; Dimmer, A.; Humpel, C.; Amberger, A.; Fischer-Colbrie, R.; Nemeth, J.; Waschek, J.A.; Kieselbach, G.; Kralinger, M.; Schmid, E.; et al. VIP, PACAP-38, BDNF and ADNP in NMDA-induced excitotoxicity in the rat retina. Acta Ophthalmol. 2011, 89, 670–675. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Magrì, B.; Musumeci, G.; D’Agata, V. Activity-Dependent Neuroprotective Protein (ADNP): An Overview of Its Role in the Eye. Int. J. Mol. Sci. 2022, 23, 3654. [Google Scholar] [CrossRef] [PubMed]
- Lagrèze, W.A.; Pielen, A.; Steingart, R.; Schlunck, G.; Hofmann, H.D.; Gozes, I.; Kirsch, M. The peptides ADNF-9 and NAP increase survival and neurite outgrowth of rat retinal ganglion cells in vitro. Investig. Ophthalmol. Vis. Sci. 2005, 46, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Jehle, T.; Dimitriu, C.; Auer, S.; Knoth, R.; Vidal-Sanz, M.; Gozes, I.; Lagrèze, W.A. The neuropeptide NAP provides neuroprotection against retinal ganglion cell damage after retinal ischemia and optic nerve crush. Graefes Arch. Clin. Exp. Ophthalmol. 2008, 246, 1255–1263. [Google Scholar] [CrossRef]
- Belokopytov, M.; Shulman, S.; Dubinsky, G.; Gozes, I.; Belkin, M.; Rosner, M. Ameliorative effect of NAP on laser-induced retinal damage. Acta Ophthalmol. 2011, 89, e126–e131. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zeng, H.; She, H.; Liu, H.; Sun, N. Expression of peptide NAP in rat retinal Müller cells prevents hypoxia-induced retinal injuries and promotes retinal neurons growth. Biomed. Pharmacother. 2010, 64, 417–423. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.G.; Maugeri, G.; Rasà, D.; Federico, C.; Saccone, S.; Lazzara, F.; Fidilio, A.; Drago, F.; Bucolo, C.; D’Agata, V. NAP modulates hyperglycemic-inflammatory event of diabetic retina by counteracting outer blood retinal barrier damage. J. Cell. Physiol. 2019, 234, 5230–5240. [Google Scholar] [CrossRef]
- D’Amico, A.G.; Maugeri, G.; Rasà, D.M.; La Cognata, V.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. NAP counteracts hyperglycemia/hypoxia induced retinal pigment epithelial barrier breakdown through modulation of HIFs and VEGF expression. J. Cell. Physiol. 2018, 233, 1120–1128. [Google Scholar] [CrossRef]
- D’Amico, A.G.; Maugeri, G.; Bucolo, C.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. Nap Interferes with Hypoxia-Inducible Factors and VEGF Expression in Retina of Diabetic Rats. J. Mol. Neurosci. 2017, 61, 256–266. [Google Scholar] [CrossRef]
- Ebrahim, A.S.; Carion, T.W.; Ebrahim, T.; Win, J.; Kani, H.; Wang, Y.; Stambersky, A.; Ibrahim, A.S.; Sosne, G.; Berger, E.A. A Novel Combination Therapy Tβ4/VIP Protects against Hyperglycemia-Induced Changes in Human Corneal Epithelial Cells. Biosensors 2023, 13, 974. [Google Scholar] [CrossRef]
- Martínez, C.; Juarranz, Y.; Gutiérrez-Cañas, I.; Carrión, M.; Pérez-García, S.; Villanueva-Romero, R.; Castro, D.; Lamana, A.; Mellado, M.; González-Álvaro, I.; et al. A Clinical Approach for the Use of VIP Axis in Inflammatory and Autoimmune Diseases. Int. J. Mol. Sci. 2019, 21, 65. [Google Scholar] [CrossRef] [PubMed]
- Gomariz, R.P.; Martinez, C.; Abad, C.; Leceta, J.; Delgado, M. Immunology of VIP: A review and therapeutical perspectives. Curr. Pharm. Des. 2001, 7, 89–111. [Google Scholar] [CrossRef] [PubMed]
- Saika, S.; Muragaki, Y.; Okada, Y.; Miyamoto, T.; Ohnishi, Y.; Ooshima, A.; Kao, W.W. Sonic hedgehog expression and role in healing corneal epithelium. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2577–2585. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, L.; Li, Y.; Sun, D.; Chen, R.; Dou, S.; Liu, T.; Zhang, S.; Zhou, Q.; Xie, L. Interference of sympathetic overactivation restores limbal stem/progenitor cells function and accelerates corneal epithelial wound healing in diabetic mice. Biomed. Pharmacother. 2023, 161, 114523. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Magrì, B.; Giunta, S.; Saccone, S.; Federico, C.; Bucolo, C.; Musumeci, G.; D’Agata, V. Protective effect of pituitary adenylate cyclase activating polypeptide in diabetic keratopathy. Peptides 2023, 170, 171107. [Google Scholar] [CrossRef] [PubMed]
- Sibilia, M.; Kroismayr, R.; Lichtenberger, B.M.; Natarajan, A.; Hecking, M.; Holcmann, M. The epidermal growth factor receptor: From development to tumorigenesis. Differentiation 2007, 75, 770–787. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.P.; Li, Y.; Ljubimov, A.V.; Yu, F.S. High glucose suppresses epidermal growth factor receptor/phosphatidylinositol 3-kinase/Akt signaling pathway and attenuates corneal epithelial wound healing. Diabetes 2009, 58, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Yu, F.S. Impaired epithelial wound healing and EGFR signaling pathways in the corneas of diabetic rats. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3301–3308. [Google Scholar] [CrossRef]
- Jiang, Q.W.; Kaili, D.; Freeman, J.; Lei, C.Y.; Geng, B.C.; Tan, T.; He, J.F.; Shi, Z.; Ma, J.J.; Luo, Y.H.; et al. Diabetes inhibits corneal epithelial cell migration and tight junction formation in mice and human via increasing ROS and impairing Akt signaling. Acta Pharmacol. Sin. 2019, 40, 1205–1211. [Google Scholar] [CrossRef]
- Ljubimov, A.V. Diabetic complications in the cornea. Vision Res. 2017, 139, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Magrì, B.; Giunta, S.; Musumeci, G.; Saccone, S.; Federico, C.; Scollo, D.; Longo, A.; Avitabile, T.; et al. Regulation of UV-B-Induced Inflammatory Mediators by Activity-Dependent Neuroprotective Protein (ADNP)-Derived Peptide (NAP) in Corneal Epithelium. Int. J. Mol. Sci. 2023, 24, 6895. [Google Scholar] [CrossRef] [PubMed]
- Hamrah, P.; Huq, S.O.; Liu, Y.; Zhang, Q.; Dana, M.R. Corneal immunity is mediated by heterogeneous population of antigen-presenting cells. J. Leukoc. Biol. 2003, 74, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Blanco, T.; Musayeva, A.; Singh, R.B.; Nakagawa, H.; Lee, S.; Alemi, H.; Gonzalez-Nolasco, B.; Ortiz, G.; Wang, S.; Kahale, F.; et al. The impact of donor diabetes on corneal transplant immunity. Am. J. Transplant. 2023, 23, 1345–1358. [Google Scholar] [CrossRef] [PubMed]
- Idan-Feldman, A.; Schirer, Y.; Polyzoidou, E.; Touloumi, O.; Lagoudaki, R.; Grigoriadis, N.C.; Gozes, I. Davunetide (NAP) as a preventative treatment for central nervous system complications in a diabetes rat model. Neurobiol. Dis. 2011, 44, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.; Verma, A.; Alagorie, A.R. Association of severity of diabetic retinopathy with corneal endothelial and thickness changes in patients with diabetes mellitus. Eye 2022, 36, 1202–1208. [Google Scholar] [CrossRef]
- Saif, P.S.; Salman, A.E.G.; Omran, N.A.H.; Farweez, Y.A.T. Assessment of Diabetic Retinopathy Vascular Density Maps. Clin. Ophthalmol. 2020, 14, 3941–3953. [Google Scholar] [CrossRef]
- Buonfiglio, F.; Wasielica-Poslednik, J.; Pfeiffer, N.; Gericke, A. Diabetic Keratopathy: Redox Signaling Pathways and Therapeutic Prospects. Antioxidants 2024, 13, 120. [Google Scholar] [CrossRef]
- Valdehita, A.; Bajo, A.M.; Schally, A.V.; Varga, J.L.; Carmena, M.J.; Prieto, J.C. Vasoactive intestinal peptide (VIP) induces transactivation of EGFR and HER2 in human breast cancer cells. Mol. Cell. Endocrinol. 2009, 302, 41–48. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neuropeptide | Abbreviation | Corneal Distribution | References |
|---|---|---|---|
| Vasoactive intestinal peptide | VIP | Nerve endings; endothelium | [45,47,48,49] |
| Pituitary adenylate cyclase-activating polypeptide | PACAP | Nerve endings; stroma | [50] |
| Activity-dependent Neuroprotective Protein | ADNP | Epithelium; stroma | [51] |
| Substance P | SP | Epithelium; stroma | [52,53,54] |
| Calcitonin gene-related peptide | CGRP | Nerve endings | [55] |
| Adrenomedullin | ADM | Nerve endings | [56] |
| Neuropeptide Y | NPY | Stroma | [57] |
| Somatostatin | SST | Whole cornea | [58] |
| Alpha melanocyte-stimulating hormone | α-MSH | Whole cornea | [59] |
| Galanin | GALP | Epithelium, stroma, and endothelium | [57,60] |
| Neurotensin | NT | Stroma | [57,61] |
| Brain natriuretic peptide | BNP | Epithelium | [57] |
| Nerve growth factor | NGF | Epithelium | [62] |
| Opioid growth factor (OGF)/met-enkephalin | OGF/MENK | Epithelium | [63,64] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maugeri, G.; D’Amico, A.G.; Magrì, B.; D’Agata, V. Involvement of Vasoactive Intestinal Peptide Family Members in Diabetic Keratopathy. Appl. Sci. 2024, 14, 1754. https://doi.org/10.3390/app14051754
Maugeri G, D’Amico AG, Magrì B, D’Agata V. Involvement of Vasoactive Intestinal Peptide Family Members in Diabetic Keratopathy. Applied Sciences. 2024; 14(5):1754. https://doi.org/10.3390/app14051754
Chicago/Turabian StyleMaugeri, Grazia, Agata Grazia D’Amico, Benedetta Magrì, and Velia D’Agata. 2024. "Involvement of Vasoactive Intestinal Peptide Family Members in Diabetic Keratopathy" Applied Sciences 14, no. 5: 1754. https://doi.org/10.3390/app14051754
APA StyleMaugeri, G., D’Amico, A. G., Magrì, B., & D’Agata, V. (2024). Involvement of Vasoactive Intestinal Peptide Family Members in Diabetic Keratopathy. Applied Sciences, 14(5), 1754. https://doi.org/10.3390/app14051754