

Exploring the Effects of Gratitude Voice Waves on Cellular Behavior: A Pilot Study in Affective Mechanotransduction

 ,
,  ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Design
2.3. Set-Up
2.3.1. Audio Recording System
2.3.2. EEG Data Recordings
2.3.3. Passive Vibro-Acoustic Treatment
2.3.4. Electro-Acoustic Radiation System
2.3.5. Noise and Vibration Measurements
2.4. Data Analyses
2.4.1. Voice Synthesis Method
2.4.2. Audio Filtering and Processing
2.4.3. Audio Analysis Magnitude-Squared Coherence
2.4.4. Audio Spectrogram
2.4.5. Affective Text Algorithm
2.4.6. EEG Pre-Processing and Filtering
2.4.7. Estimating the Power Spectral Density (PSD) for Each Frequency Band
2.4.8. EEG Statistical Analysis
2.5. Biological Assays
2.5.1. Crystal Violet Staining
2.5.2. Immunocytochemistry
2.5.3. Image Processing
2.5.4. Statistical Analysis
3. Results
3.1. Evaluation of the Affective State
3.2. Audio Analysis
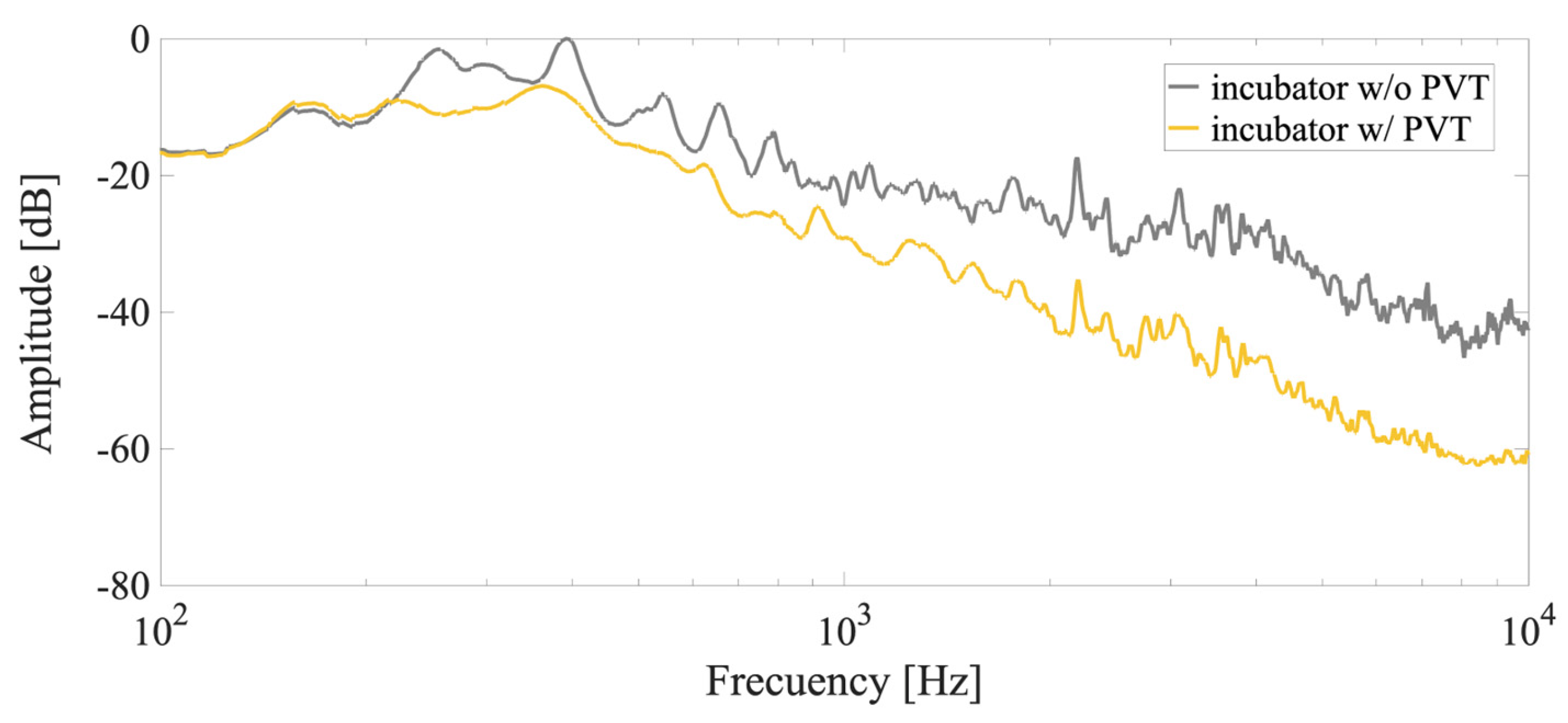
3.3. Characterization of the Environment and Electro-Acoustic Radiation System
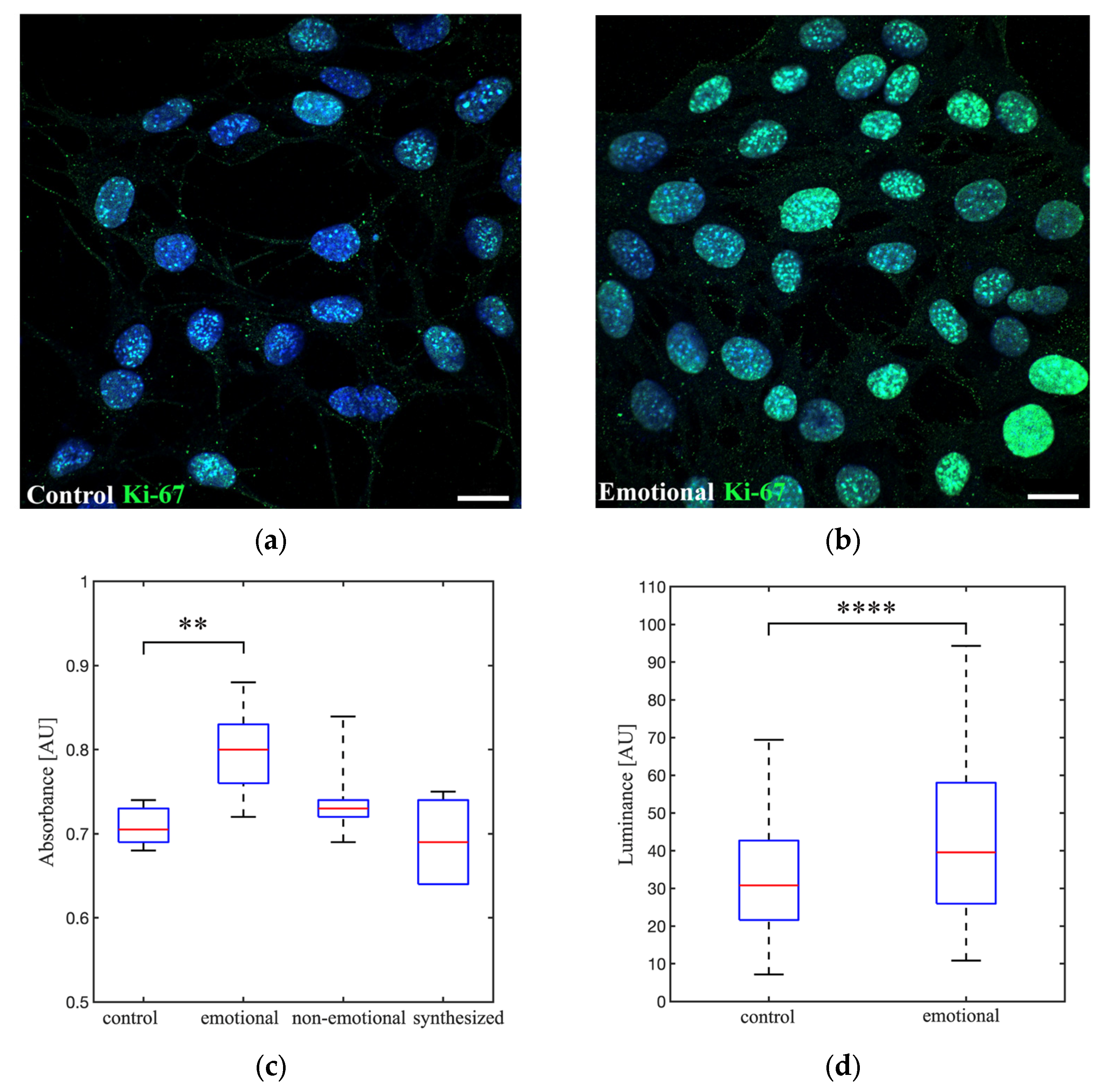
3.4. Effects of Emotional, Non-Emotional and Synthesized AAW on Cell Proliferation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Gratitude Letter
- Gracias por mostrarme que aquello que soy en realidad va mucho más allá de lo que incluso puedo llegar a imaginar ahora. Gracias por estar siempre a mi lado ofreciéndome una mano para andar en esta realidad junto a ti. Gracias también, por andar frente a mí apartando cualquier obstáculo. Desde el primer momento que te reconocí sentí que un universo entero se abría dentro de mi mente y de mi corazón. En ese momento se abrió un reconocimiento de miles de años al servicio del amor, al servicio de la paz.
- Todo aquello que me enseñas me transforma de una forma tan profunda que ni tan siquiera puedo llegar a entender. Esta transformación que ha ido calando a través del tiempo me permite hoy en día mirar la vida y a mí mismo desde una perspectiva completamente distinta a la que lo hacía antes de conocerte.
- Esta transformación me permite compartir aquello que tú me enseñas con muchas otras personas por varios lugares del planeta. El agradecimiento que siento cuando pongo palabras a tu enseñanza es un espacio inconmensurable dentro de mi corazón. Gracias por tu paciencia, tu amor y tu entrega hacia mí.
Appendix A.2. Don Quixote from la Mancha
- En un lugar de La Mancha, del cual no quiero acordarme, no hace mucho tiempo vivía un hidalgo de los de lanza en astillero, escudo antiguo, rocín flaco y galgo corredor. Una olla de algo más vaca que carnero, salpicón la mayoría de las noches, duelos y quebrantos los sábados, lentejas los viernes, algún palomino de añadido los domingos, consumían las tres cuartas partes de su hacienda. El resto se completaba con un traje de velarte, calzas de terciopelo para las fiestas, con sus pantuflas del mismo material, y los días entre semana se honraba con su traje de paño fino. Tenía en su casa una ama que pasaba de los cuarenta años y una sobrina que no llegaba a los veinte, y un mozo de campo y plaza que tanto ensillaba el rocín como tomaba la podadera. La edad de nuestro hidalgo rondaba los cincuenta años. Era de complexión recia, delgado de carnes, enjuto de rostro, gran madrugador y amigo de la caza. Dicen que tenía el sobrenombre de «Quijada», o «Quesada», sobre esto hay alguna diferencia entre los autores que escriben de este caso, aunque por conjeturas verosímiles se entiende que se llamaba «Quijana». Pero esto importa poco a nuestro cuento: basta que en la narración no se salga un punto de la verdad.
- Es, pues, de saber que este mencionado hidalgo, en los ratos que estaba ocioso —que eran la mayor parte del año—, se daba a leer libros de caballerías, con tanta afición y gusto, que olvidó casi por completo el ejercicio de la caza y aun la administración de su hacienda; y llegó a tal punto su curiosidad y desatino en esto, que vendió muchas fanegas de tierra de sembradura para comprar libros de caballerías en que leer, y, así, llevó a su casa todos cuantos pudo encontrar; y, de todos, ningunos le parecían tan bien como los que compuso el famoso Feliciano de Silva, porque la claridad de su prosa y aquellas intrincadas razones suyas le parecían de perlas, y más cuando llegaba a leer aquellos requiebros y cartas de desafíos, donde en muchas partes hallaba escrito: «La razón de la sinrazón que a mi razón se hace, de tal manera mi razón enflaquece, que con razón me quejo de la vuestra hermosura». Y también cuando leía: «Los altos cielos que de vuestra divinidad divinamente con las estrellas os fortifican y os hacen merecedora del merecimiento que merece vuestra grandeza».
References
- Darwin, C. The Expression of the Emotions in Man and Animals; Cambridge University Press: Cambridge, UK, 2013; ISBN 9781108061834. [Google Scholar]
- Ekman, P. Are There Basic Emotions? Psychol. Rev. 1992, 99, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Scherer, K.; Johnstone, T.; Klasmeyer, G. Vocal Expression of Emotion. In Handbook of Affective Sciences; Davidson, R.J., Scherer, K.R., Goldsmith, H.H., Eds.; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Russell, J.A.; Bachorowski, J.A.; Fernández-Dols, J.M. Facial and Vocal Expressions of Emotion. Annu. Rev. Psychol. 2003, 54, 329–349. [Google Scholar] [CrossRef] [PubMed]
- Banse, R.; Scherer, K.R. Acoustic Profiles in Vocal Emotion Expression. J. Pers. Soc. Psychol. 1996, 70, 614–636. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, S.P. Acoustic Characteristics of Vocal Emotions Simulated by Actors. Percept. Mot. Ski. 1999, 89, 1195–1208. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, A.; Adolphs, R. Emotion Perception from Face, Voice, and Touch: Comparisons and Convergence. Trends Cogn. Sci. 2017, 21, 216–228. [Google Scholar] [CrossRef]
- Ekman, P. Facial Expressions of Emotion: An Old Controversy and New Findings. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1992, 335, 63–69. [Google Scholar] [CrossRef]
- Johnstone, T.; Scherer, K.R. Vocal Communication of Emotion. In Handbook of Emotions; Lewis, M., Haviland-Jones, J., Eds.; Guilford Press: New York, NY, USA, 2000. [Google Scholar]
- Scherer, K.R. Vocal Correlates of Emotional Arousal and Affective Disturbance. In Handbook of Social Psychophysiology; Wagner, H., Manstead, A., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 1989. [Google Scholar]
- Pietrowicz, M.; Hasegawa-Johnson, M.; Karahalios, K.G. Acoustic Correlates for Perceived Effort Levels in Male and Female Acted Voices. J. Acoust. Soc. Am. 2017, 142, 792–811. [Google Scholar] [CrossRef]
- Kadiri, S.R.; Alku, P. Glottal Features for Classification of Phonation Type from Speech and Neck Surface Accelerometer Signals. Comput. Speech Lang. 2021, 70, 101232. [Google Scholar] [CrossRef]
- Grichkovtsova, I.; Morel, M.; Lacheret, A. The Role of Voice Quality and Prosodic Contour in Affective Speech Perception. Speech Commun. 2012, 54, 414–429. [Google Scholar] [CrossRef]
- Gobl, C.; Ní Chasaide, A. The Role of Voice Quality in Communicating Emotion, Mood and Attitude. Speech Commun. 2003, 40, 189–212. [Google Scholar] [CrossRef]
- Birkholz, P.; Martin, L.; Willmes, K.; Kröger, B.J.; Neuschaefer-Rube, C. The Contribution of Phonation Type to the Perception of Vocal Emotions in German: An Articulatory Synthesis Study. J. Acoust. Soc. Am. 2015, 137, 1503–1512. [Google Scholar] [CrossRef] [PubMed]
- Kuang, J.; Liberman, M. Integrating Voice Quality Cues in the Pitch Perception of Speech and Non-Speech Utterances. Front. Psychol. 2018, 9, 2147. [Google Scholar] [CrossRef] [PubMed]
- Honorof, D.N.; Whalen, D.H. Perception of Pitch Location within a Speaker’s F0 Range. J. Acoust. Soc. Am. 2005, 117, 2193–2200. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-Y.; Dutton, L.; Ram, G. The Role of Speaker Gender Identification in Relative Fundamental Frequency Height Estimation from Multispeaker, Brief Speech Segments. J. Acoust. Soc. Am. 2010, 128, 384–388. [Google Scholar] [CrossRef]
- Lieberman, P.; Michaels, S.B. Some Aspects of Fundamental Frequency and Envelope Amplitude as Related to the Emotional Content of Speech. J. Acoust. Soc. Am. 1962, 34, 922–927. [Google Scholar] [CrossRef]
- Sobin, C.; Alpert, M. Emotion in Speech: The Acoustic Attributes of Fear, Anger, Sadness, and Joy. J. Psycholinguist. Res. 1999, 28, 347–365. [Google Scholar] [CrossRef]
- Juslin, P.N.; Laukka, P. Communication of Emotions in Vocal Expression and Music Performance: Different Channels, Same Code? Psychol. Bull. 2003, 129, 770–814. [Google Scholar] [CrossRef]
- Juslin, P.N.; Scherer, K.R. Vocal Expression of Affect. In The New Handbook of Methods in Nonverbal Behavior Research; Harrigan, J.A., Rosenthal, R., Scherer, K.R., Eds.; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Breitenstein, C.; Van Lancker, D.; Daum, I. The Contribution of Speech Rate and Pitch Variation to the Perception of Vocal Emotions in a German and an American Sample. Cogn. Emot. 2010, 15, 57–79. [Google Scholar] [CrossRef]
- Davletcharova, A.; Sugathan, S.; Abraham, B.; James, A.P. Detection and Analysis of Emotion from Speech Signals. Procedia Comput. Sci. 2015, 58, 91–96. [Google Scholar] [CrossRef]
- Wood, A.M.; Froh, J.J.; Geraghty, A.W.A. Gratitude and Well-Being: A Review and Theoretical Integration. Clin. Psychol. Rev. 2010, 30, 890–905. [Google Scholar] [CrossRef]
- Wood, A.M.; Maltby, J.; Gillett, R.; Linley, P.A.; Joseph, S. The Role of Gratitude in the Development of Social Support, Stress, and Depression: Two Longitudinal Studies. J. Res. Pers. 2008, 42, 854–871. [Google Scholar] [CrossRef]
- Fox, G.R.; Kaplan, J.; Damasio, H.; Damasio, A. Neural Correlates of Gratitude. Front. Psychol. 2015, 6, 1491. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Gao, X.; Zhou, Y.; Zhou, X. Decomposing Gratitude: Representation and Integration of Cognitive Antecedents of Gratitude in the Brain. J. Neurosci. 2018, 38, 4886–4898. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Yu, J.; Song, M.; Yu, C.; Wang, F.; Sun, P.; Wang, D.; Zhang, D. EEG Correlates of Ten Positive Emotions. Front. Hum. Neurosci. 2017, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Toepfer, S.M.; Cichy, K.; Peters, P. Letters of Gratitude: Further Evidence for Author Benefits. J. Happiness Stud. 2012, 13, 187–201. [Google Scholar] [CrossRef]
- Boehm, J.K.; Lyubomirsky, S.; Sheldon, K.M. A Longitudinal Experimental Study Comparing the Effectiveness of Happiness-Enhancing Strategies in Anglo Americans and Asian Americans. Cogn. Emot. 2011, 25, 1263. [Google Scholar] [CrossRef]
- Bradley, M.M.; Lang, P.J. Affective Norms for English Words (ANEW): Instruction Manual and Affective Ratings; University of Florida: Gainesville, FL, USA, 1999. [Google Scholar]
- Mohammad, S.M.; Turney, P.D. Crowdsourcing a Word-Emotion Association Lexicon. Comput. Intell. 2013, 29, 436–465. [Google Scholar] [CrossRef]
- Kumeta, M.; Takahashi, D.; Takeyasu, K.; Yoshimura, S.H. Cell Type-Specific Suppression of Mechanosensitive Genes by Audible Sound Stimulation. PLoS ONE 2018, 13, e0188764. [Google Scholar] [CrossRef]
- Kwak, D.; Combriat, T.; Wang, C.; Scholz, H.; Danielsen, A.; Jensenius, A.R. Music for Cells? A Systematic Review of Studies Investigating the Effects of Audible Sound Played Through Speaker-Based Systems on Cell Cultures. Music Sci. 2022, 5, 20592043221080965. [Google Scholar] [CrossRef]
- Alenghat, F.J.; Ingber, D.E. Mechanotransduction: All Signals Point to Cytoskeleton, Matrix, and Integrins. Sci. STKE 2002, 2002, pe6. [Google Scholar] [CrossRef]
- Ross, T.D.; Coon, B.G.; Yun, S.; Baeyens, N.; Tanaka, K.; Ouyang, M.; Schwartz, M.A. Integrins in Mechanotransduction. Curr. Opin. Cell Biol. 2013, 25, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in Mechanotransduction. Nature 2011, 474, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Toepfer, S.M.; Walker, K. Letters of Gratitude: Improving Well-Being through Expressive Writing. J. Writ. Res. 2009, 1, 181–198. [Google Scholar] [CrossRef]
- Rowan, A.J.; Tolunsky, E. Primer of EEG: With a Mini-Atlas; Butterworth-Heinemann: Oxford, UK, 2003. [Google Scholar]
- Gish, H. The Magnitude Squared Coherence Estimate: A Geometric View. In Proceedings of the ICASSP ’84. IEEE International Conference on Acoustics, Speech, and Signal Processing, San Diego, CA, USA, 19–21 March 1984; Volume 1. [Google Scholar] [CrossRef]
- Malekpour, S.; Gubner, J.A.; Sethares, W.A. Measures of Generalized Magnitude-Squared Coherence: Differences and Similarities. J. Frankl. Inst. 2018, 355, 2932–2950. [Google Scholar] [CrossRef]
- Mohammad, S.M. Obtaining Reliable Human Ratings of Valence, Arousal, and Dominance for 20,000 English Words. In Proceedings of the ACL 2018—56th Annual Meeting of the Association for Computational Linguistics, Proceedings of the Conference (Long. Papers), Melbourne, Australia, 15–20 July 2018; Volume 1, pp. 174–184. [Google Scholar] [CrossRef]
- Russell, J.A. A Circumplex Model of Affect. J. Pers. Soc. Psychol. 1980, 39, 1161–1178. [Google Scholar] [CrossRef]
- Cohen, M.X. Analyzing Neural Time Series Data: Theory and Practice; The MIT Press: Cambridge, MA, USA, 2014; ISBN 9780262319553. [Google Scholar]
- Welch, P.D. The Use of Fast Fourier Transform for the Estimation of Power Spectra: A Method Based on Time Averaging Over Short, Modified Periodograms. IEEE Trans. Audio Electroacoust. 1967, 15, 70–73. [Google Scholar] [CrossRef]
- Delorme, A.; Beischel, J.; Michel, L.; Boccuzzi, M.; Radin, D.; Mills, P. Electrocortical Activity Associated with Subjective Communication with the Deceased. Front. Psychol. 2013, 4, 834. [Google Scholar] [CrossRef]
- Yamada, T.; Meng, E. Guía Práctica Para Pruebas Neurofisiológicas Clínicas-EEG; Ovid Technologies: New York, NY, USA, 2020. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Martínez-Cagigal, V. Multiple Testing Toolbox-File Exchange-MATLAB Central. Available online: https://ch.mathworks.com/matlabcentral/fileexchange/70604-multiple-testing-toolbox (accessed on 28 July 2024).
- Meyer, M.; Lamers, D.; Kayhan, E.; Hunnius, S.; Oostenveld, R. Enhancing Reproducibility in Developmental EEG Research: BIDS, Cluster-Based Permutation Tests, and Effect Sizes. Dev. Cogn. Neurosci. 2021, 52, 101036. [Google Scholar] [CrossRef]
- Maris, E.; Oostenveld, R. Nonparametric Statistical Testing of EEG- and MEG-Data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef]
- Sayyad, Z.; Sirohi, K.; Radha, V.; Swarup, G. 661W Is a Retinal Ganglion Precursor-like Cell Line in Which Glaucoma-Associated Optineurin Mutants Induce Cell Death Selectively. Sci. Rep. 2017, 7, 16855. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.; Ding, X.Q.; Saadi, A.; Agarwal, N.; Naash, M.I.; Al-Ubaidi, M.R. Expression of Cone-Photoreceptor-Specific Antigens in a Cell Line Derived from Retinal Tumors in Transgenic Mice. Investig. Ophthalmol. Vis. Sci. 2004, 45, 764–768. [Google Scholar] [CrossRef] [PubMed]
- Mickuviene, I.; Kirveliene, V.; Juodka, B. Experimental Survey of Non-Clonogenic Viability Assays for Adherent Cells In Vitro. Toxicol. Vitro 2004, 18, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Kaufman, P.D. Ki-67: More than a Proliferation Marker. Chromosoma 2018, 127, 175. [Google Scholar] [CrossRef] [PubMed]
- Tursunov, A.; Kwon, S.; Pang, H.S. Discriminating Emotions in the Valence Dimension from Speech Using Timbre Features. Appl. Sci. 2019, 9, 2470. [Google Scholar] [CrossRef]
- Brück, C.; Kreifelts, B.; Wildgruber, D. Emotional Voices in Context: A Neurobiological Model of Multimodal Affective Information Processing. Phys. Life Rev. 2011, 8, 383–403. [Google Scholar] [CrossRef]
- Nussbaum, C.; Schirmer, A.; Schweinberger, S.R. Contributions of Fundamental Frequency and Timbre to Vocal Emotion Perception and Their Electrophysiological Correlates. Soc. Cogn. Affect. Neurosci. 2022, 17, 1145–1154. [Google Scholar] [CrossRef]
- Lin, C.D.; Radu, C.M.; Vitiello, G.; Romano, P.; Polcari, A.; Iliceto, S.; Simioni, P.; Tona, F. Sounds Stimulation on In Vitro HL1 Cells: A Pilot Study and a Theoretical Physical Model. Int. J. Mol. Sci. 2021, 22, 10156. [Google Scholar] [CrossRef]
- Kechagia, J.Z.; Ivaska, J.; Roca-Cusachs, P. Integrins as Biomechanical Sensors of the Microenvironment. Nat. Rev. Mol. Cell Biol. 2019, 20, 457–473. [Google Scholar] [CrossRef]
- Matthews, B.D.; Thodeti, C.K.; Ingber, D.E. Activation of Mechanosensitive Ion Channels by Forces Transmitted Through Integrins and the Cytoskeleton. Curr. Top. Membr. 2007, 58, 59–85. [Google Scholar] [CrossRef]
- Delling, M.; Indzhykulian, A.A.; Liu, X.; Li, Y.; Xie, T.; Corey, D.P.; Clapham, D.E. Primary Cilia Are Not Calcium-Responsive Mechanosensors. Nature 2016, 531, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Muhamed, I.; Chowdhury, F.; Maruthamuthu, V. Biophysical Tools to Study Cellular Mechanotransduction. Bioengineering 2017, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.M.; Wiesolek, H.L.; Sumagin, R. ICAM-1: A Master Regulator of Cellular Responses in Inflammation, Injury Resolution, and Tumorigenesis. J. Leukoc. Biol. 2020, 108, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Wu, J.W.; Wang, C.W.; Jang, A.C.C. Hippo Signaling-Mediated Mechanotransduction in Cell Movement and Cancer Metastasis. Front. Mol. Biosci. 2019, 6, 157. [Google Scholar] [CrossRef] [PubMed]
- Spindler, M.C.; Redolfi, J.; Helmprobst, F.; Kollmannsberger, P.; Stigloher, C.; Benavente, R. Electron Tomography of Mouse LINC Complexes at Meiotic Telomere Attachment Sites with and without Microtubules. Commun. Biol. 2019, 2, 376. [Google Scholar] [CrossRef]
- Chang, L.; Li, M.; Shao, S.; Li, C.; Ai, S.; Xue, B.; Hou, Y.; Zhang, Y.; Li, R.; Fan, X.; et al. Nuclear Peripheral Chromatin-Lamin B1 Interaction Is Required for Global Integrity of Chromatin Architecture and Dynamics in Human Cells. Protein Cell 2022, 13, 258–280. [Google Scholar] [CrossRef]
- Peric-Hupkes, D.; van Steensel, B. Role of the Nuclear Lamina in Genome Organization and Gene Expression. Cold Spring Harb. Symp. Quant. Biol. 2010, 75, 517–524. [Google Scholar] [CrossRef]
- Fu, M.; Hu, Y.; Lan, T.; Guan, K.L.; Luo, T.; Luo, M. The Hippo Signalling Pathway and Its Implications in Human Health and Diseases. Signal Transduct. Target. Ther. 2022, 7, 376. [Google Scholar] [CrossRef]
- Zhang, X.; Abdelrahman, A.; Vollmar, B.; Zechner, D. The Ambivalent Function of YAP in Apoptosis and Cancer. Int. J. Mol. Sci. 2018, 19, 3770. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
del Rosario-Gilabert, D.; Carbajo, J.; Valenzuela-Miralles, A.; Vigué-Guix, I.; Ruiz, D.; Esquiva, G.; Gómez-Vicente, V. Exploring the Effects of Gratitude Voice Waves on Cellular Behavior: A Pilot Study in Affective Mechanotransduction. Appl. Sci. 2024, 14, 9400. https://doi.org/10.3390/app14209400
del Rosario-Gilabert D, Carbajo J, Valenzuela-Miralles A, Vigué-Guix I, Ruiz D, Esquiva G, Gómez-Vicente V. Exploring the Effects of Gratitude Voice Waves on Cellular Behavior: A Pilot Study in Affective Mechanotransduction. Applied Sciences. 2024; 14(20):9400. https://doi.org/10.3390/app14209400
Chicago/Turabian Styledel Rosario-Gilabert, David, Jesús Carbajo, Antonio Valenzuela-Miralles, Irene Vigué-Guix, Daniel Ruiz, Gema Esquiva, and Violeta Gómez-Vicente. 2024. "Exploring the Effects of Gratitude Voice Waves on Cellular Behavior: A Pilot Study in Affective Mechanotransduction" Applied Sciences 14, no. 20: 9400. https://doi.org/10.3390/app14209400
APA Styledel Rosario-Gilabert, D., Carbajo, J., Valenzuela-Miralles, A., Vigué-Guix, I., Ruiz, D., Esquiva, G., & Gómez-Vicente, V. (2024). Exploring the Effects of Gratitude Voice Waves on Cellular Behavior: A Pilot Study in Affective Mechanotransduction. Applied Sciences, 14(20), 9400. https://doi.org/10.3390/app14209400