
Let’s Talk about Sex Hormone Receptors and Their Physical Interaction with Sonic Hedgehog Protein: A Computational Study with Emphasis on Progesterone Receptor




Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
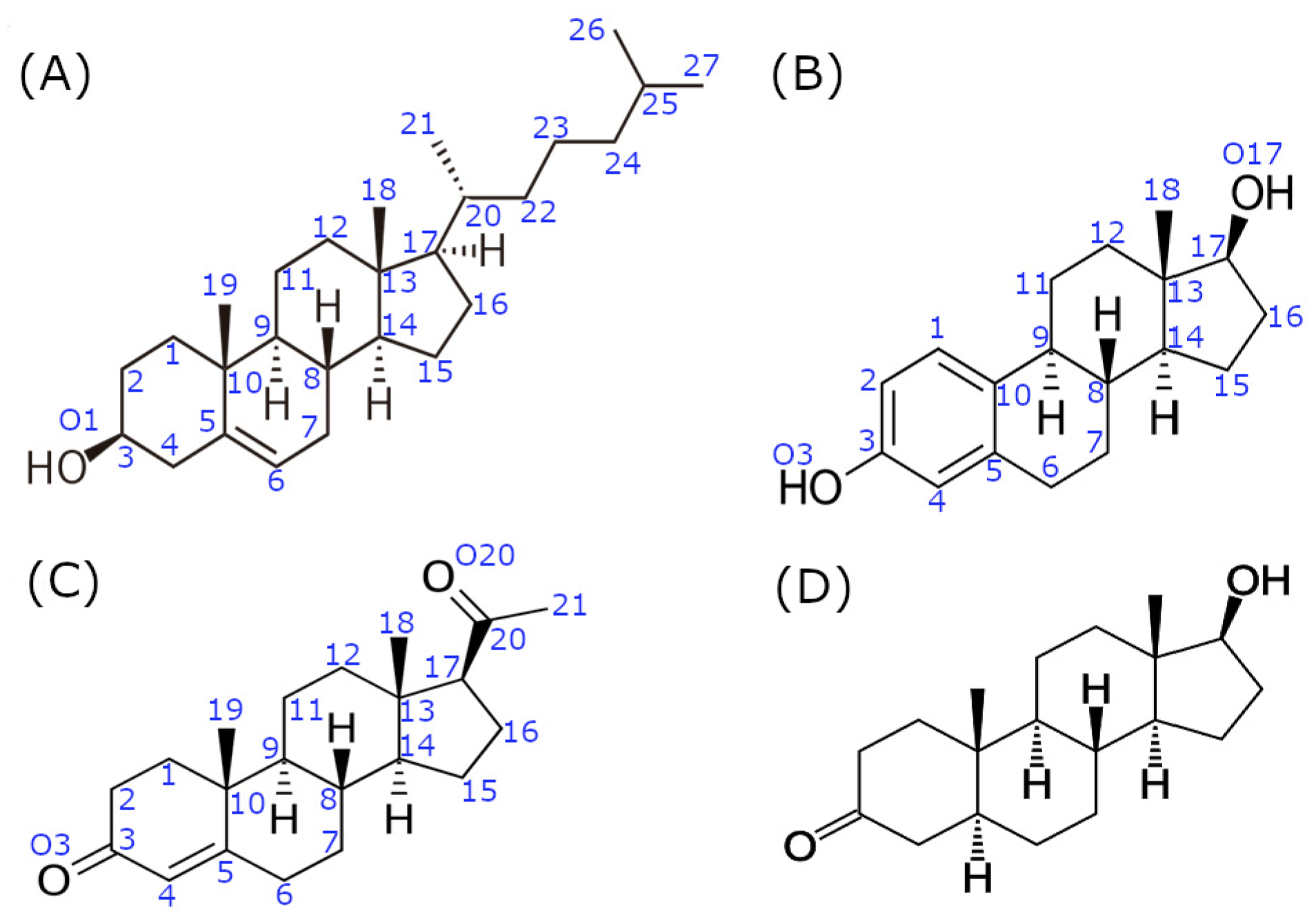
2.1. System Preparation
2.2. Molecular Docking and Molecular Modeling Details
2.3. MD Simulations Details
2.4. MM/G(P)BSA Calculations
2.5. Analysis
3. Results
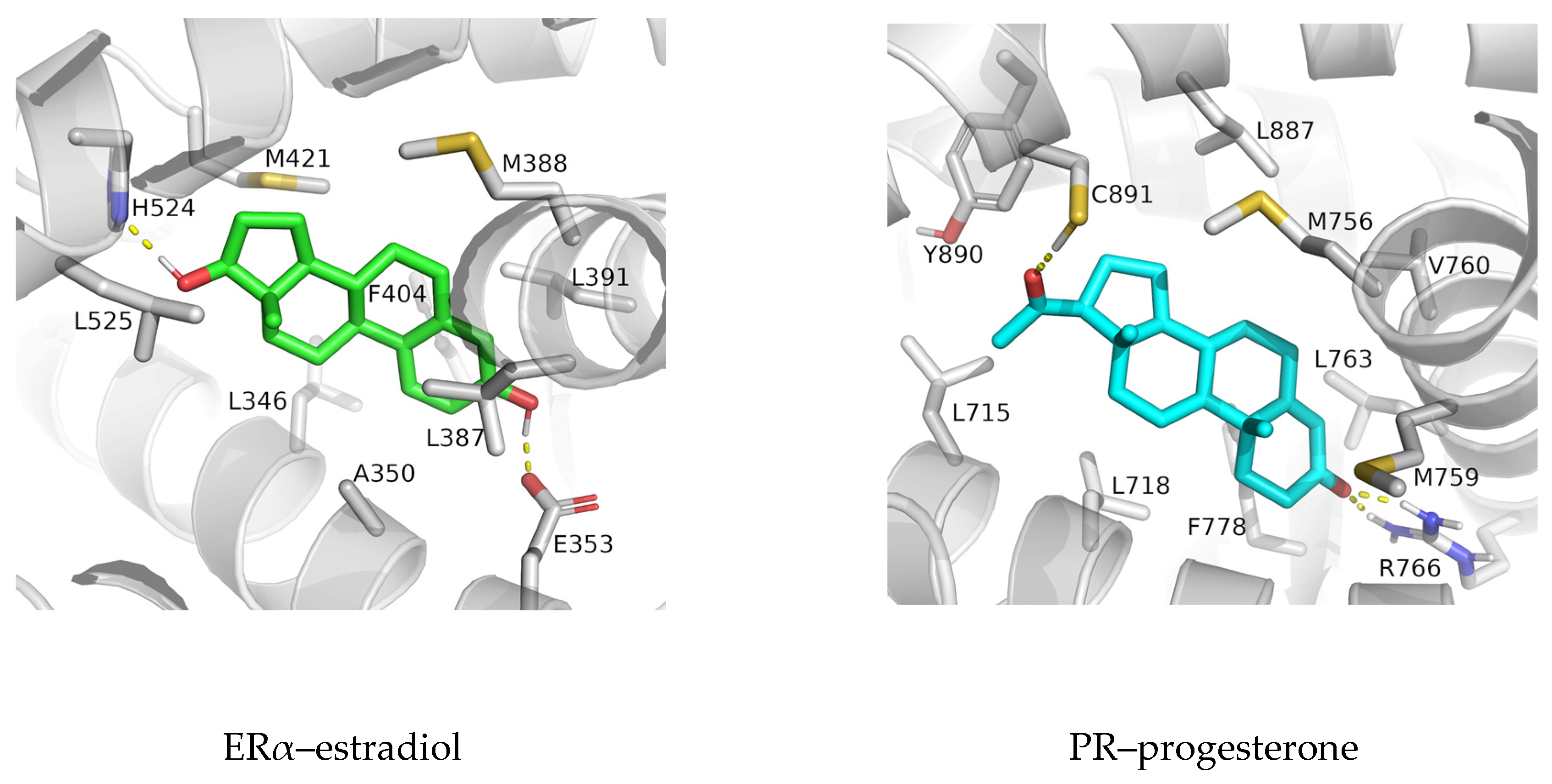
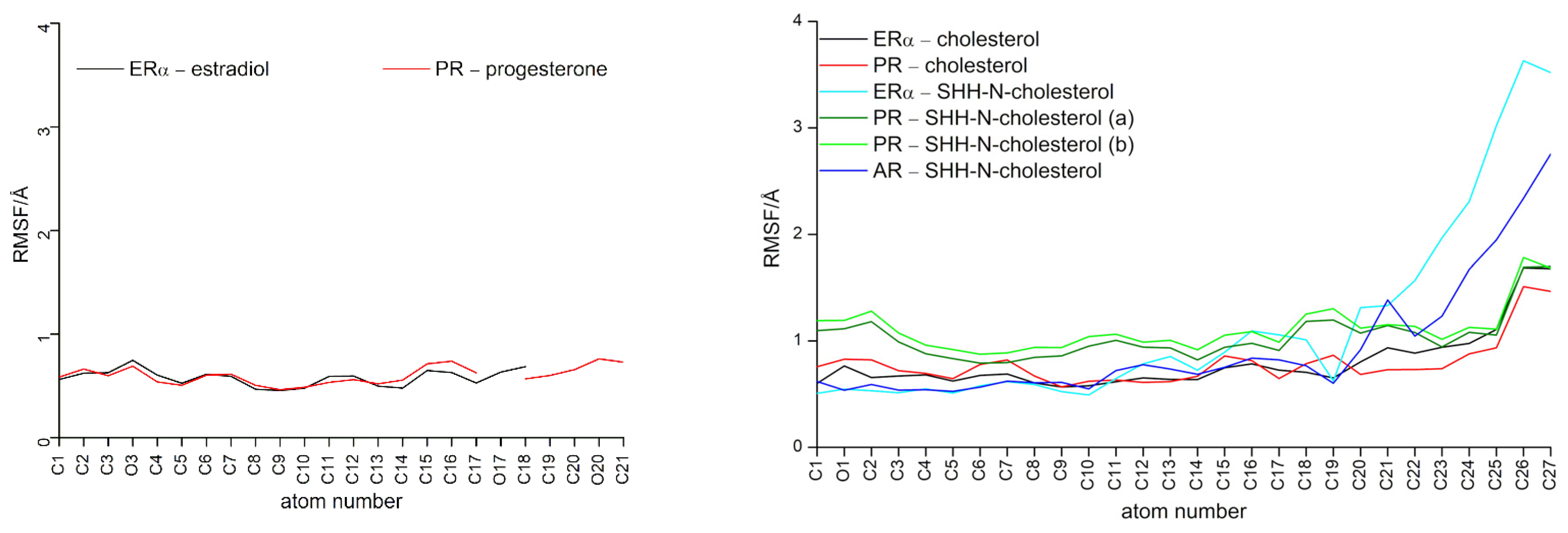
3.1. Complexes with Estradiol and Progesterone
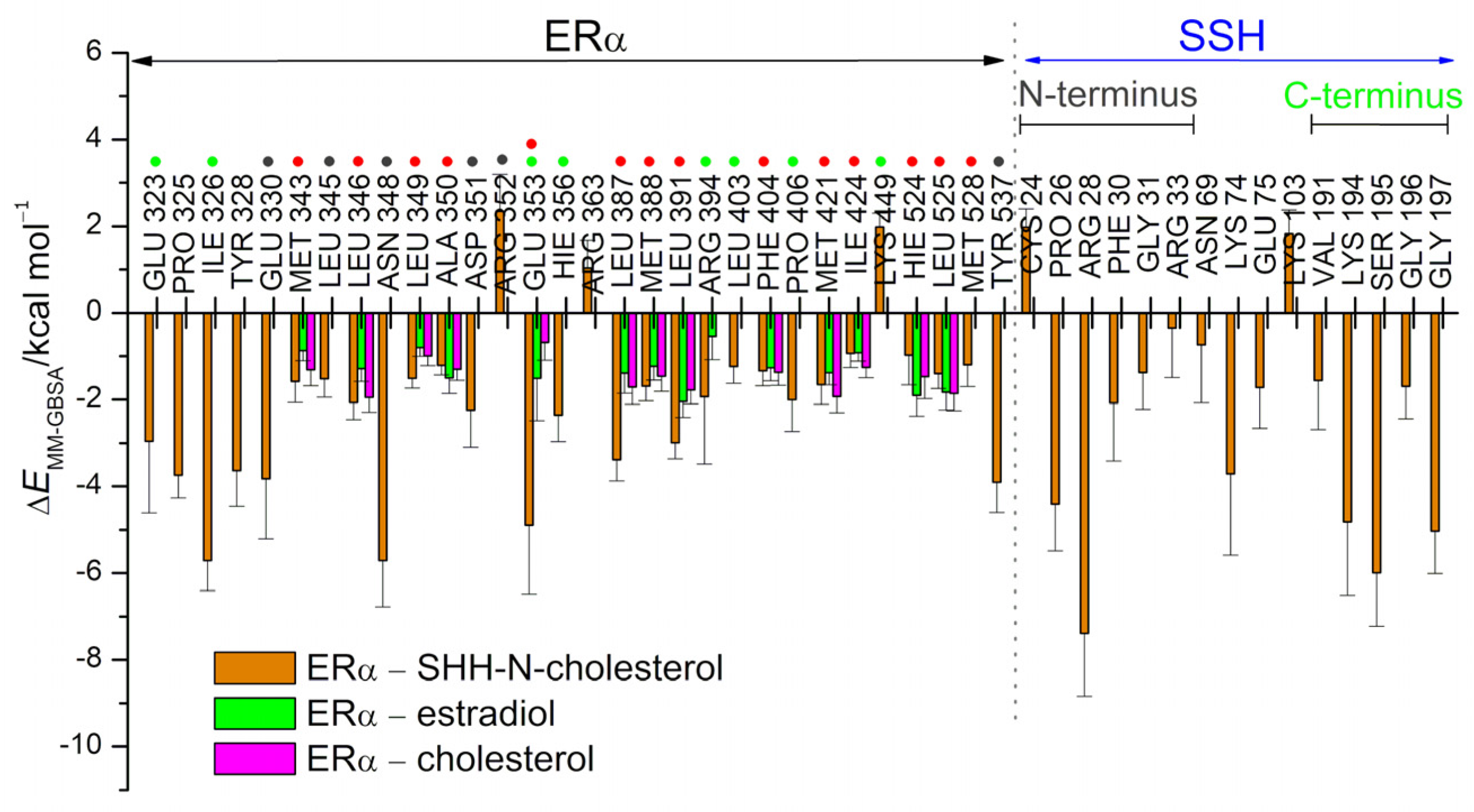
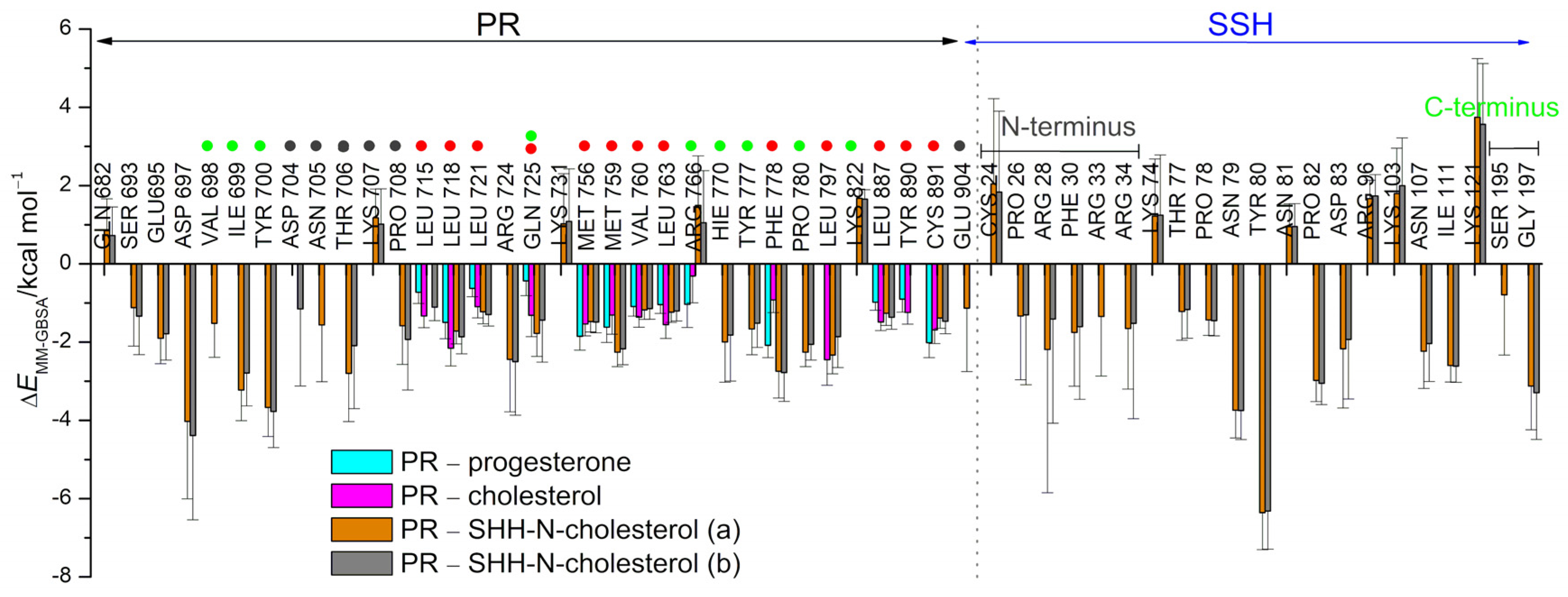
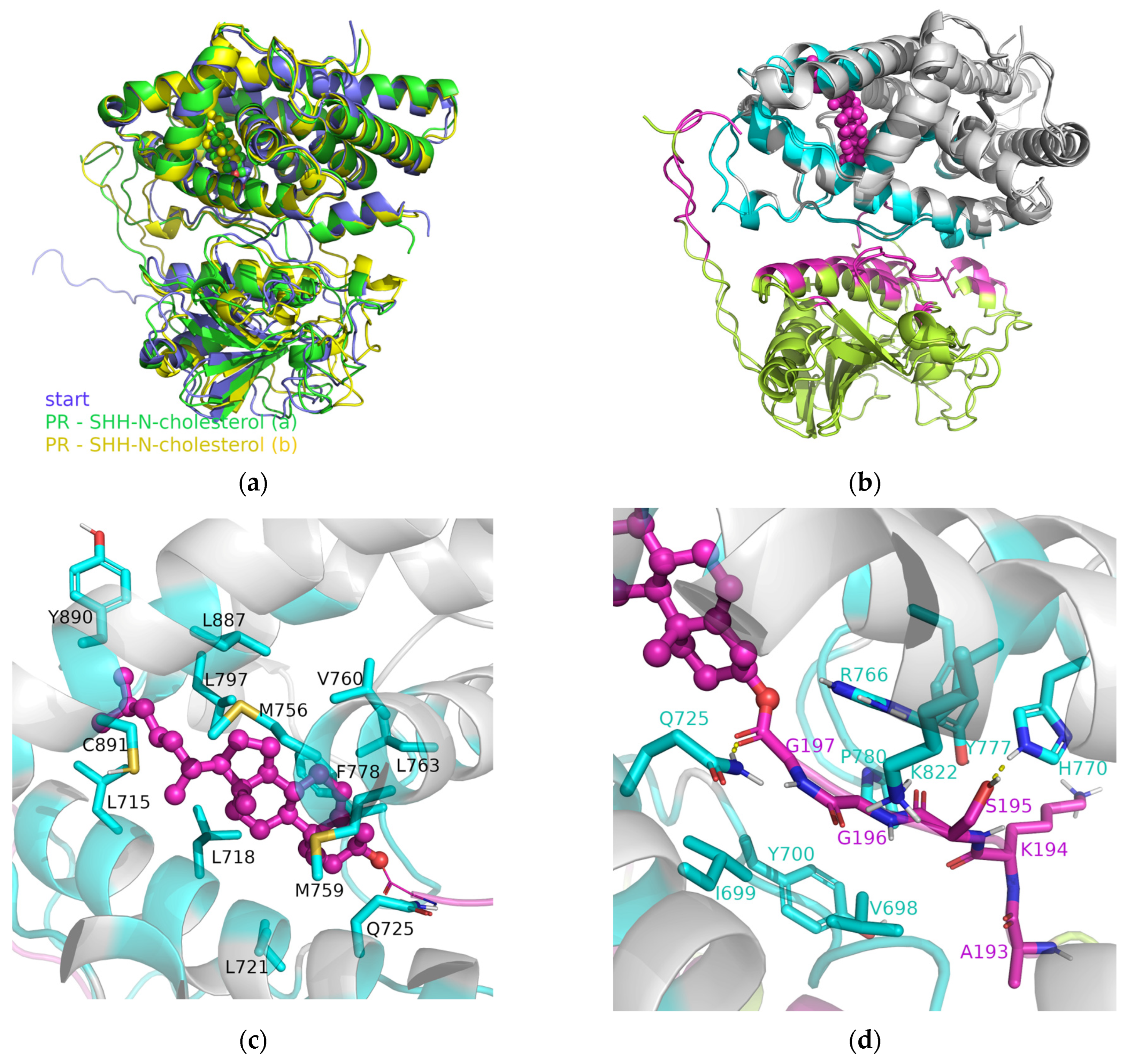
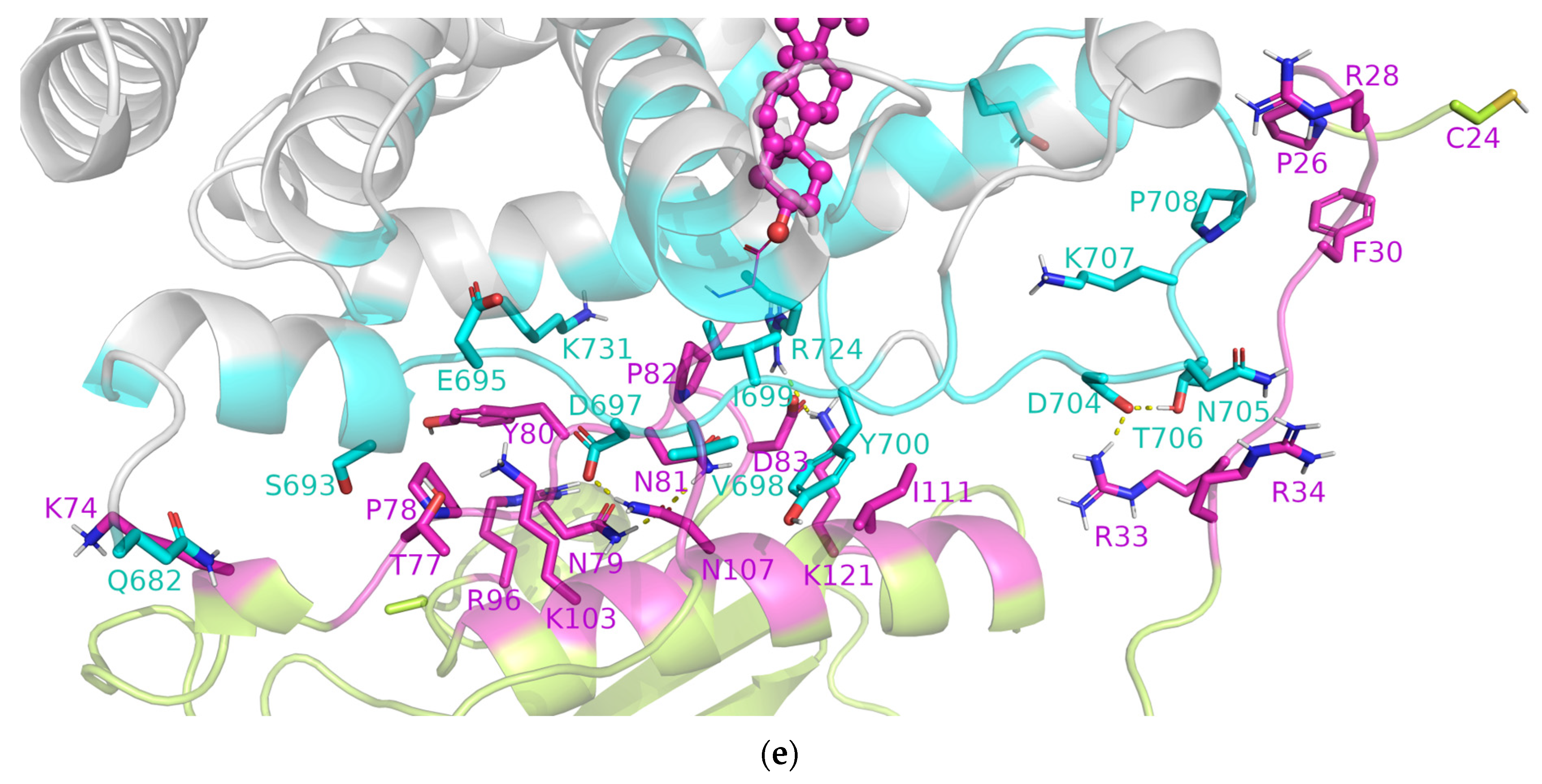
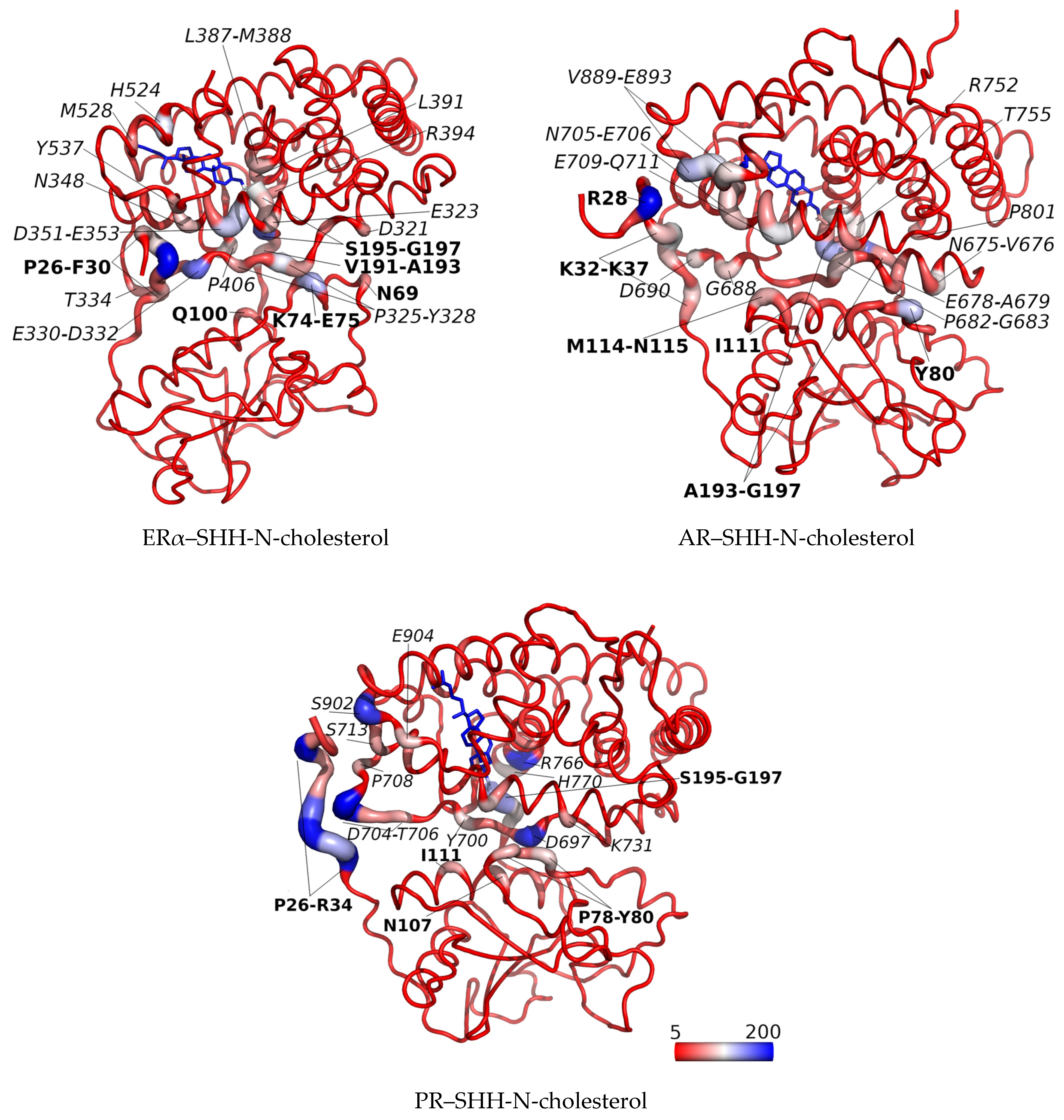
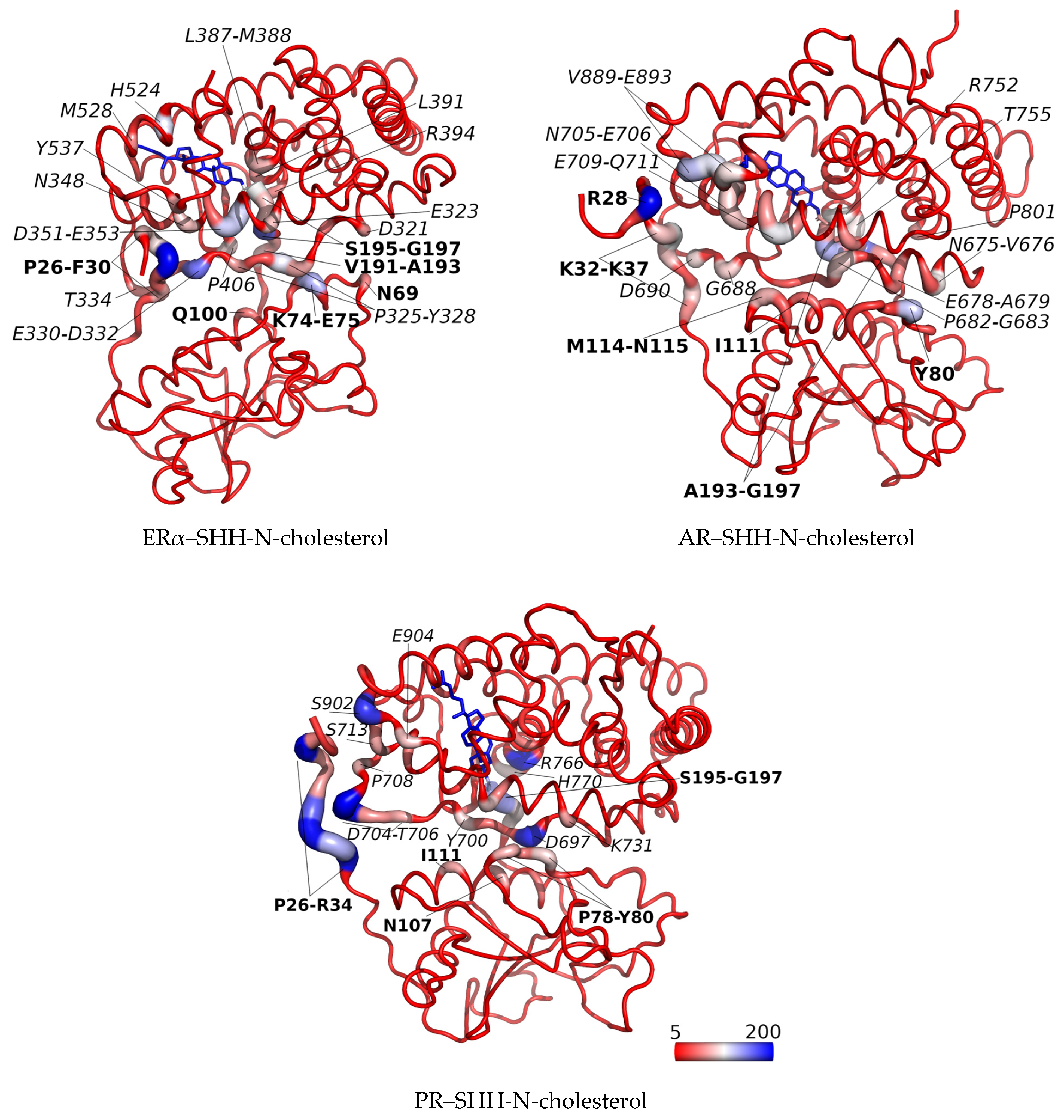
3.2. SHRs in Complex with SHH-N-Cholesterol (PP Complexes)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sigafoos, A.N.; Paradise, B.D.; Fernandez-Zapico, M.E. Hedgehog/GLI Signaling Pathway: Transduction, Regulation, and Implications for Disease. Cancers 2021, 13, 3410. [Google Scholar] [CrossRef] [PubMed]
- Varjosalo, M.; Li, S.-P.; Taipale, J. Divergence of Hedgehog Signal Transduction Mechanism between Drosophila and Mammals. Dev. Cell 2006, 10, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Kurtović, M.; Piteša, N.; Bartoniček, N.; Ozretić, P.; Musani, V.; Čonkaš, J.; Petrić, T.; King, C.; Sabol, M. RNA-Seq and ChIP-Seq Identification of Unique and Overlapping Targets of GLI Transcription Factors in Melanoma Cell Lines. Cancers 2022, 14, 4540. [Google Scholar] [CrossRef] [PubMed]
- Dawber, R.J.; Hebbes, S.; Herpers, B.; Docquier, F.; van den Heuvel, M. Differential Range and Activity of Various Forms of the Hedgehog Protein. BMC Dev. Biol. 2005, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Varjosalo, M.; Taipale, J. Hedgehog: Functions and Mechanisms. Genes Dev. 2008, 22, 2454–2472. [Google Scholar] [CrossRef] [PubMed]
- Ciepla, P.; Magee, A.I.; Tate, E.W. Cholesterylation: A Tail of Hedgehog. Biochem. Soc. Trans. 2015, 43, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Resh, M.D. Palmitoylation of Hedgehog Proteins by Hedgehog Acyltransferase: Roles in Signalling and Disease. Open Biol. 2021, 11, 200414. [Google Scholar] [CrossRef]
- Musani, V.; Ozretić, P.; Trnski, D.; Sabol, M.; Poduje, S.; Tošić, M.; Šitum, M.; Levanat, S. Potential Hot Spot for de Novo Mutations in PTCH1 Gene in Gorlin Syndrome Patients: A Case Report of Twins from Croatia. Croat. Med. J. 2018, 59, 20–24. [Google Scholar] [CrossRef]
- Car, D.; Sabol, M.; Musani, V.; Ozretić, P.; Levanat, S. Epigenetic Regulation of the Hedgehog-Gli Signaling Pathway in Cancer. Period. Biol. 2010, 112, 419–423. [Google Scholar]
- Budimir, I.; Tomasović-Lončarić, Č.; Kralik, K.; Čonkaš, J.; Eljuga, D.; Žic, R.; Gorjanc, B.; Tucaković, H.; Caktaš, D.; Jaman, J.; et al. Higher Expressions of SHH and AR Are Associated with a Positive Receptor Status and Have Impact on Survival in a Cohort of Croatian Breast Cancer Patients. Life 2022, 12, 1559. [Google Scholar] [CrossRef]
- Levanat, S.; Sabol, M.; Musani, V.; Ozretić, P.; Trnski, D. Hedgehog Signaling Pathway as Genetic and Epigenetic Target in Ovarian Tumors. Curr. Pharm. Des. 2016, 22, 73–94. [Google Scholar] [CrossRef]
- Liao, X.; Siu, M.K.; Au, C.W.; Chan, Q.K.; Chan, H.Y.; Wong, E.S.; Ip, P.P.; Ngan, H.Y.; Cheung, A.N. Aberrant Activation of Hedgehog Signaling Pathway Contributes to Endometrial Carcinogenesis through β-Catenin. Mod. Pathol. 2009, 22, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Bushman, W. Hedgehog Signaling in Prostate Development, Regeneration and Cancer. J. Dev. Biol. 2016, 4, 30. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.E.; Feigelson, H.S. Hormonal Carcinogenesis. Carcinogenesis 2000, 21, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Rochefort, H.; Chalbos, D. The Role of Sex Steroid Receptors on Lipogenesis in Breast and Prostate Carcinogenesis: A Viewpoint. Horm. Cancer 2010, 1, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Olefsky, J.M. Nuclear Receptor Minireview Series. J. Biol. Chem. 2001, 276, 36863–36864. [Google Scholar] [CrossRef] [PubMed]
- Sever, R.; Glass, C.K. Signaling by Nuclear Receptors. Cold Spring Harb. Perspect. Biol. 2013, 5, a016709. [Google Scholar] [CrossRef] [PubMed]
- Sabol, M.; Trnski, D.; Uzarevic, Z.; Ozretic, P.; Musani, V.; Rafaj, M.; Cindric, M.; Levanat, S. Combination of Cyclopamine and Tamoxifen Promotes Survival and Migration of MCF-7 Breast Cancer Cells—Interaction of Hedgehog-Gli and Estrogen Receptor Signaling Pathways. PLoS ONE 2014, 9, e114510. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L.; Auchus, R.J. The Molecular Biology, Biochemistry, and Physiology of Human Steroidogenesis and Its Disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef]
- Trnski, D.; Sabol, M.; Tomić, S.; Štefanac, I.; Mrčela, M.; Musani, V.; Rinčić, N.; Kurtović, M.; Petrić, T.; Levanat, S.; et al. SHH-N Non-Canonically Sustains Androgen Receptor Activity in Androgen-Independent Prostate Cancer Cells. Sci. Rep. 2021, 11, 14880. [Google Scholar] [CrossRef]
- Sartorius, C.A.; Melville, M.Y.; Hovland, A.R.; Tung, L.; Takimoto, G.S.; Horwitz, K.B. A Third Transactivation Function (AF3) of Human Progesterone Receptors Located in the Unique N-Terminal Segment of the B-Isoform. Mol. Endocrinol. 1994, 8, 1347–1360. [Google Scholar] [CrossRef] [PubMed]
- Hopp, T.A.; Weiss, H.L.; Hilsenbeck, S.G.; Cui, Y.; Allred, D.C.; Horwitz, K.B.; Fuqua, S.A.W. Breast Cancer Patients with Progesterone Receptor PR-A-Rich Tumors Have Poorer Disease-Free Survival Rates. Clin. Cancer Res. 2004, 10, 2751–2760. [Google Scholar] [CrossRef] [PubMed]
- Jaffrain-Rea, M.L.; Petrangeli, E.; Ortolani, F.; Fraioli, B.; Lise, A.; Esposito, V.; Spagnoli, L.G.; Tamburrano, G.; Frati, L.; Gulino, A. Cellular Receptors for Sex Steroids in Human Pituitary Adenomas. J. Endocrinol. 1996, 151, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Liu, D. Gene Signatures of Estrogen and Progesterone Receptor Pathways Predict the Prognosis of Colorectal Cancer. FEBS J. 2016, 283, 3115–3133. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, G.; De Leo, S.; Perrino, M.; Rossi, S.; Tosi, D.; Cirello, V.; Colombo, C.; Bulfamante, G.; Vicentini, L.; Fugazzola, L. Impact of Estrogen and Progesterone Receptor Expression on the Clinical and Molecular Features of Papillary Thyroid Cancer. Eur. J. Endocrinol. 2015, 173, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Elmaogod, E.A.; Khairy, D.; Mahmoud, A. Clinicopathological Role of Progesterone Hormone in IDH-Mutant Astrocytoma. J. Microsc. Ultrastruct. 2023. [Google Scholar] [CrossRef]
- Deli, T.; Orosz, M.; Jakab, A. Hormone Replacement Therapy in Cancer Survivors—Review of the Literature. Pathol. Oncol. Res. 2020, 26, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Lan, K.-C.; Ou, Y.-C.; Wu, C.-H.; Kang, H.-Y.; Chuang, I.-C.; Fu, H.-C. Highly Expressed Progesterone Receptor B Isoform Increases Platinum Sensitivity and Survival of Ovarian High-Grade Serous Carcinoma. Cancers 2021, 13, 5578. [Google Scholar] [CrossRef]
- Yang, S.; Thiel, K.W.; Leslie, K.K. Progesterone: The Ultimate Endometrial Tumor Suppressor. Trends Endocrinol. Metab. 2011, 22, 145–152. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.; Das, C.; Kumari, M.; Sen, A.; Mukhopadhyay, B.; Mukhopadhyay, B. Spectrum of Meningioma with Special Reference to Prognostic Utility of ER, PR and Ki67 Expression. J. Lab. Physicians 2017, 9, 308–313. [Google Scholar] [CrossRef]
- Brzozowski, A.M.; Pike, A.C.W.; Dauter, Z.; Hubbard, R.E.; Bonn, T.; Engström, O.; Öhman, L.; Greene, G.L.; Gustafsson, J.-Å.; Carlquist, M. Molecular Basis of Agonism and Antagonism in the Oestrogen Receptor. Nature 1997, 389, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.P.; Sigler, P.B. Atomic Structure of Progesterone Complexed with Its Receptor. Nature 1998, 393, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, A.F.; Kinnebrew, M.; Kowatsch, C.; Ansell, T.B.; El Omari, K.; Bishop, B.; Pardon, E.; Schwab, R.A.; Malinauskas, T.; Qian, M.; et al. The Morphogen Sonic Hedgehog Inhibits Its Receptor Patched by a Pincer Grasp Mechanism. Nat. Chem. Biol. 2019, 15, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Pereira de Jésus-Tran, K.; Côté, P.; Cantin, L.; Blanchet, J.; Labrie, F.; Breton, R. Comparison of Crystal Structures of Human Androgen Receptor Ligand-binding Domain Complexed with Various Agonists Reveals Molecular Determinants Responsible for Binding Affinity. Protein Sci. 2006, 15, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Tao, H.; He, J.; Huang, S.-Y. The HDOCK Server for Integrated Protein–Protein Docking. Nat. Protoc. 2020, 15, 1829–1852. [Google Scholar] [CrossRef] [PubMed]
- Izadi, S.; Anandakrishnan, R.; Onufriev, A.V. Building Water Models: A Different Approach. J. Phys. Chem. Lett. 2014, 5, 3863–3871. [Google Scholar] [CrossRef]
- Tian, C.; Kasavajhala, K.; Belfon, K.A.A.; Raguette, L.; Huang, H.; Migues, A.N.; Bickel, J.; Wang, Y.; Pincay, J.; Wu, Q.; et al. Ff19SB: Amino-Acid-Specific Protein Backbone Parameters Trained against Quantum Mechanics Energy Surfaces in Solution. J. Chem. Theory Comput. 2020, 16, 528–552. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General Amber Force Field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Tomić, A.; Abramić, M.; Spoljarić, J.; Agić, D.; Smith, D.M.; Tomić, S. Human Dipeptidyl Peptidase III: Insights into Ligand Binding from a Combined Experimental and Computational Approach. J. Mol. Recognit. 2011, 24, 804–814. [Google Scholar] [CrossRef]
- Loncharich, R.J.; Brooks, B.R.; Pastor, R.W. Langevin Dynamics of Peptides: The Frictional Dependence of Isomerization Rates OfN-Acetylalanyl-N?-Methylamide. Biopolymers 1992, 32, 523–535. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Salomon-Ferrer, R.; Götz, A.W.; Poole, D.; Le Grand, S.; Walker, R.C. Routine Microsecond Molecular Dynamics Simulations with AMBER on GPUs. 2. Explicit Solvent Particle Mesh Ewald. J. Chem. Theory Comput. 2013, 9, 3878–3888. [Google Scholar] [CrossRef] [PubMed]
- Götz, A.W.; Williamson, M.J.; Xu, D.; Poole, D.; Le Grand, S.; Walker, R.C. Routine Microsecond Molecular Dynamics Simulations with AMBER on GPUs. 1. Generalized Born. J. Chem. Theory Comput. 2012, 8, 1542–1555. [Google Scholar] [CrossRef] [PubMed]
- Salomon-Ferrer, R.; Case, D.A.; Walker, R.C. An Overview of the Amber Biomolecular Simulation Package. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2013, 3, 198–210. [Google Scholar] [CrossRef]
- Swanson, J.M.J.; Henchman, R.H.; McCammon, J.A. Revisiting Free Energy Calculations: A Theoretical Connection to MM/PBSA and Direct Calculation of the Association Free Energy. Biophys. J. 2004, 86, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Connolly, M.L. Analytical Molecular Surface Calculation. J. Appl. Crystallogr. 1983, 16, 548–558. [Google Scholar] [CrossRef]
- Siebold, C.; Rohatgi, R. The Inseparable Relationship Between Cholesterol and Hedgehog Signaling. Annu. Rev. Biochem. 2023, 92, 273–298. [Google Scholar] [CrossRef]
- Tukachinsky, H.; Kuzmickas, R.P.; Jao, C.Y.; Liu, J.; Salic, A. Dispatched and Scube Mediate the Efficient Secretion of the Cholesterol-Modified Hedgehog Ligand. Cell Rep. 2012, 2, 308–320. [Google Scholar] [CrossRef]
- Rozeboom, B.; Dey, N.; De, P. ER+ Metastatic Breast Cancer: Past, Present, and a Prescription for an Apoptosis-Targeted Future. Am. J. Cancer Res. 2019, 9, 2821–2831. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex | Time Interval/ns | (ΔHbind ± SD)/(kcal/mol) |
|---|---|---|
| ERα–estradiol | 0–500 | −12.87 ± 2.68 |
| ERα–cholesterol | −17.50 ± 3.42 | |
| PR–progesterone | −19.02 ± 2.97 | |
| PR–cholesterol | −19.48 ± 3.63 | |
| PR–SHH-N–cholesterol | 0–500 | −23.53 ± 8.33 |
| 300–500 | −20.99 ± 7.14 | |
| 500–1000 (a) * | −27.95 ± 7.80 | |
| 500–1000 (b) * | −23.25 ± 7.66 | |
| AR–SHH-N–cholesterol | 0–500 | −33.03 ± 10.27 |
| 300–500 | −29.73 ± 11.09 | |
| ERα–SHH-N–cholesterol | 0–300 | −20.70 ± 7.28 |
| 300–500 | −40.12 ± 9.25 | |
| 600–800 | −45.56 ± 9.5 | |
| 800–1000 | −36.29 ± 8.66 | |
| 1100–1350 | −42.40 ± 8.45 | |
| 1400–2000 | −51.51 ± 7.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomić, A.; Čonkaš, J.; Ozretić, P. Let’s Talk about Sex Hormone Receptors and Their Physical Interaction with Sonic Hedgehog Protein: A Computational Study with Emphasis on Progesterone Receptor. Appl. Sci. 2024, 14, 562. https://doi.org/10.3390/app14020562
Tomić A, Čonkaš J, Ozretić P. Let’s Talk about Sex Hormone Receptors and Their Physical Interaction with Sonic Hedgehog Protein: A Computational Study with Emphasis on Progesterone Receptor. Applied Sciences. 2024; 14(2):562. https://doi.org/10.3390/app14020562
Chicago/Turabian StyleTomić, Antonija, Josipa Čonkaš, and Petar Ozretić. 2024. "Let’s Talk about Sex Hormone Receptors and Their Physical Interaction with Sonic Hedgehog Protein: A Computational Study with Emphasis on Progesterone Receptor" Applied Sciences 14, no. 2: 562. https://doi.org/10.3390/app14020562
APA StyleTomić, A., Čonkaš, J., & Ozretić, P. (2024). Let’s Talk about Sex Hormone Receptors and Their Physical Interaction with Sonic Hedgehog Protein: A Computational Study with Emphasis on Progesterone Receptor. Applied Sciences, 14(2), 562. https://doi.org/10.3390/app14020562