Abstract
Background: Cognitive disorders are symptoms of degenerative neuronal diseases such as Alzheimer’s disease (AD). This study evaluated the effects of a mixture of Gastrodiae elata (GE) and Glycyrrhizae uralensis (GU) (GGW) on scopolamine-treated cognitive function disorders in vivo. Methods: To induce memory impairment and cognitive disorder, C57BL/6 mice were intraperitoneally administered scopolamine. In the experimental period, the Y-maze, passive avoidance, and water maze tests were carried out to measure the ameliorative effect of GGW on cognitive function disorder. The expression of amyloid-β (Aβ), brain-derived neurotrophic factor (BDNF), and acetylcholinesterase (AChE) in the hippocampal tissues were measured. In addition, to assess the effect of GGW on brain tissue, tissues were stained using hematoxylin and eosin (H&E). Results: GGW treatment improved latencies in the Y-maze, water maze, and passive avoidance tests compared with scopolamine treatment alone. GGW administration downregulated Aβ and AChE expression. In addition, BDNF expression was upregulated by GGW administration. Conclusion: These results suggest that GGW ameliorates impaired cognitive function in a scopolamine-induced mouse model. In addition, GGW could control the pathway of scopolamine-induced cognitive disorders by regulating Aβ, AChE, and BDNF. These results suggest that GGW could be used as a treatment agent for cognitive dysfunction.
1. Introduction
Cognitive impairment has a negative effect on cognition; that is, the ability to retrieve and store information [1]. In addition, cognitive impairment, including memory and learning failure, can be initial manifestations of neurodegenerative diseases [2] and hippocampal lesions [3]. Such cognitive dysfunction frequently presents as degenerative diseases with increasing age [4]. Many patients with cognitive impairments suffer from persistent symptoms and enormous medical expenses, which lead to social and financial problems [5]. Acetylcholinesterase inhibitors (AChEIs) (donepezil, rivastigmine, and galantamine, etc.) have been applied to remedy symptoms of mild cognitive decline [6,7]. Among AChEIs, donepezil is most frequently prescribed to patients with cognitive disorder [8] as it ameliorates acetylcholinesterase (AChE) activation in the hippocampus and cerebral cortex [9]. Despite having a clear mechanism of action, donepezil causes various adverse reactions such as diarrhea, nausea, chronic fatigue, vomiting, and insomnia [10]. Therefore, it is necessary to identify medicines of natural origin that can reduce adverse effects.
Among the various animal models of cognitive impairment, scopolamine is frequently used as a muscarinic receptor antagonist [11]. Scopolamine promotes learning delay and memory deficiency in dementia and Alzheimer’s disease (AD) models [12]. Scopolamine leads to neurocognitive disease by competitively blocking muscarinic acetylcholine (ACh) receptors and neuronal signal propagation [13]. Moreover, scopolamine causes increased ACh in the nervous system, resulting in damage to hippocampal tissues, leading to poor learning and cognitive impairment [14,15]. For this reason, the application of scopolamine is appropriate in an experimental mouse model of cognitive dysfunction.
The hippocampus, consisting of the dentate cornu ammonis (CA) 1–3 areas and gyrus, plays an important part in cognition processing, including the consolidation of memory and the learning and retrieval of information [16]. In cognitive disorders, memory loss is followed by a gradual decline of cholinergic function during hippocampal degeneration [17,18]. Amyloid β (Aβ) accumulation in the hippocampus occurs prior to cognitive decline and leads to neuronal cell inflammation, neurofibrillary tangles, and eventually, neuronal cell death [19,20]. For this reason, drug development has focused on ameliorating excess Aβ to protect neurons and the brain [21]. The cleaved Aβ precursor protein generates Aβ and self-aggregates into various oligomer sizes. Aggregated Aβ interrupts oxidative metabolism, mitochondrial activity, and cell apoptosis [22]. Aβ oligomer accumulation has also been found in degenerative neuronal diseases, such as cognitive disorders and AD [23].
Brain-derived neurotrophic factor (BDNF) performs as a role in memory learning, cognitive function, and regulation of synaptic plasticity [24]. BDNF is also related to neuronal cell growth and survival, and neurotransmitter release [25]. Elevated BDNF expression and ACh levels are essential for maintaining cognitive function, memory retrieval, and nerve growth in basal forebrain [26]. In the cholinergic system, decreased levels of ACh result in excessive AChE expression, causing cognitive disorders and memory dysfunction [27]. Upregulated AChE, found in the hippocampus and cortex of patients with AD, causes cognitive disorders [28]. The fragmentation and cleaved amyloid precursor protein (APP) is caused by β- and γ-secretase (BACE-1), a major enzyme in the nervous system [29]. Therefore, treatment of cognitive disorders by ameliorating BDNF, AChE, and BACE-1 is necessary.
Gastrodia elata Blume (GE), used as a traditional medicine and dietary supplement, has been used to cure dizziness, headaches, and rheumatic inflammation [30]. Some rhizomes of GE have been used in major medical applications, including polysaccharides, phenolic compounds, and organic acids [31]. Gastrodin is considered the main bioactive constituent of the rhizome of GE [32]. Previous studies have shown that GE has various beneficial effects for depression [33], diabetes [34], obesity [35], vascular inflammation [36], and colon cancer [37]. Moreover, GE has been announced to prevent against H2O2-induced neuronal cell cytotoxicity [38], alleviate Parkinson’s disease induced by L-DOPA administration [39], Alzheimer’s disease [40], and cognitive dysfunction [41]. Glycyrrhiza uralensis (GU) has been used in Asia as a herbal alternative medicine [42] and is known to remedy skin diseases, digestive disorders, and respiratory diseases [43]. Various pharmaceutical constituents of GU are flavonoids and triterpenoid saponins, such as liquiritin, licochalcones, liquiritigenin, glycyrrhizic acid, and glycyrrhetinic acid [43]. These constituents of GU have been reported to exhibit anti-inflammatory [44], antioxidant [45], antiviral [46], and antibacterial effects [47]. In addition, both GE and GU have been shown to have therapeutic effects on neuronal diseases, such as neurotoxicity [48] and cholinergic memory loss [49]. However, the effectiveness of a mixture of GE and GU for cognitive disorders in vivo has not yet been uncovered.
This study aimed to reveal the effect of a mixture of GE and GU on scopolamine-induced cognitive disorder in vivo. To establish their regulatory effects on cognitive function, behavioral tests such as the water maze, Y-maze, and passive avoidance tests were conducted. In addition, we measured AChE, BDNF, and BACE-1 expression levels to determine the pathway of such cognitive disorders.
2. Materials and Methods
2.1. Reagents
Primary antibodies (BDNF, p-APP, β-actin) for performing Western blot analysis were acquired from Cell Signaling Technology (Danvers, MA, USA). Primary BACE-1 antibody for Western blotting was acquired from Abcam (Cambridge, MA, USA). Secondary antibodies (anti-rabbit or anti-mouse monoclonal antibodies) were obtained from Jackson ImmunoResearch (West Grove, PA, USA). Additionally, 10× phosphate buffer saline (PBS) was obtained from ELPIS BIOTECH (Daejeon, Republic of Korea) and bovine serum albumin (BSA) was obtained from MPbio Chemicals (Irvine, CA, USA). The Total BDNF ELISA kit was obtained from R&D Systems (Minneapolis, MN, USA). AChE and Aβ ELISA kits were purchased from NOVUS Biologicals (Centennial, CO, USA). Donepezil (purity 98%) and scopolamine (purity 95%) were acquired from Sigma Aldrich (St. Louis, MO, USA).
2.2. GE and GU Extract Preparation
The gastrodia elata‘s rhizome (GER), cultivated in Muju-gun, 30 kg was obtained by MJ HEALTH FOOD Co., Ltd. (Muju-gun, Jeollabuk-do, Republic of Korea) and was decocted into 300 L of distilled water for 6 h at 90 °C. A 25 μm polyethersulfone syringe filter was used to filter the GER extract. The vacuum evaporation was performed at 65 °C. The concentrated substrate was deep-frozen over a 24 h period at −75 °C before being freeze-dried at −85 °C at a chamber pressure of 5 mTorr for 60 h (Yield of GER; 14.6%). The glycyrrhiza uralensis‘s radix (GUR) was obtained by MJ HEALTH FOOD Co., Ltd. (Muju-gun, Jeollabuk-do, Republic of Korea) and 15 kg was decocted with 150 L of sterile distilled water for 3 h at 100 °C. The GUR extract was filtrated through a 25 μm polyethersulfone filter, and then vacuum evaporation was performed at 65 °C. Afterward, the concentrated substrate was deep-frozen for 24 h at −75 °C, and then freeze-dried for 48 h at a chamber pressure of 5 mTorr condition at −85 °C (Yield of GUR; 19.2%). The ratio of the mixture of GER and GUR was 7:3. This mixture is referred to as GGW73.
2.3. Animals
Male C57BL/6 eight-week-old mice (DBL, Eumseong, Republic of Korea) were acclimated for one week to the absence of specific pathogens before the behavioral experiment. A temperature of 21 ± 2 °C and a humidity of 45–55% were maintained for experimental mice under a light and dark cycle for 12 h and placed in a cage. Tap water and feed were provided ad libitum. All experiments were conducted in accordance with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals, and the Institutional Animal Care and Use Committee of Woosuk University (approval ID: WS2020-08).
Mice were randomly divided into eight groups (group 1, control 0.9% saline; group 2, scopolamine (SCP) 1 mg/kg; group 3, SCP + GER 100 mg/kg; group 4, SCP + GUR 100 mg/kg; group 5, SCP + GGW73 10 mg/kg; group 6, SCP + GGW73 100 mg/kg; group 7, SCP + GGW73 200 mg/kg, and; group 8, SCP + Donepezil 5 mg/kg). GER, GUR, and GGW73 extracts were orally administered daily for 12 days. The extracts were orally administered 1 h before behavioral tests. Scopolamine was orally administered 30 min prior to the behavioral test. On the last day of the experiment, brain tissues were collected and used for analysis. Serum blood urea nitrogen (BUN), aspartate aminotransferase (AST), alanine aminotransferase (ALT), and creatinine levels were quantified.
To measure the toxicity of GGW73 in mice, we orally administered various concentrations (312.5, 625, 1250, 2500, 5000 mg/kg) of GGW73 to 6-week-old ICR mice for 2 weeks. On the final experimental day, mouse blood was isolated from the orbital venous plexus. The serum was then separated by centrifugation (12,879× g, 10 min). Serum ALT, AST, creatinine, and BUN levels were measured using SCL (Yongin, Gyeonggi-do, Republic of Korea).
2.4. Y-Maze Test
A Y-shaped maze with three arms at 120 °C from each other (35 × 10 × 15) was used to conduct the Y-maze test. Each arm was identified as A, B, or C. Mice were allowed to freely search the three arms. The locomotion activities were quantified using the total number of arm entries during a 5 min session. Spontaneous alternation was defined as entering all three arms on consecutive occasions. The following calculation is defined as the percentage of alternation:
2.5. Passive Avoidance Test
The passive avoidance test was carried out in two chambers that were juxtaposed as illuminated and dark chambers. Each test involved a training trial and a test trial. To test the training trial, an illuminated chamber was prepared to place mice initially placed in it. When the mouse entered the dark chamber, an electrical shock (0.3 mA) was delivered through stainless steel rods for 3 s. Once the mouse entered the dark compartment, latency times were measured using a stopwatch. 24 h after the training trial, a test trial was conducted, and latency times to enter the dark chamber again were checked for up to 180 s.
2.6. Water Maze Test
To test the water maze, a tank (45 cm height and 85 cm in diameter) filled with water at approximately 25 °C was used. One of quadrants contained a 8-cm diameter circular escape platform placed at a fixed position 1 cm below the water surface. Visual cues were observed around the maze. The training trials were tested twice daily for 5 days. To search for the platform, the mice were randomly placed in the water in a quadrant and were given 120 s. The mice were permitted to stay on the platform for 10 s, if the mice successfully escaped onto the platform within the given time. The mice were led to the platform and allowed to stay there for 10 s, if the mice did not find the platform within the given time. The test trials measured the time taken to reach the platform.
2.7. Enzyme-Linked Immunosorbent Assay
To determine Aβ, AChE, BDNF, and BACE-1 levels, mice hippocampal tissues were used. On the last day of the experiment, the brains of each group of mice were obtained. The hippocampus (20 mg) was homogenized in 1 × PBS at 5500× g for 30 min using a Precellys 24 tissue homogenizer (Bertin Technologies, Rockville, MD, USA). The homogenized tissues were centrifuged for 10 min at 12,879× g and the obtained supernatants were used in the experiment. Each well of a 96-well immunoplate was coated with primary antibodies (100 μL) and then incubated overnight in refrigerator (4 °C). The 96-well immuno plates were washed thrice with wash buffer 300 μL (1 × PBS including 0.05% (v/v) tween-20). After each well was blocked with 1 × PBS containing 1% (w/v) bovine serum albumin (BSA) for 60 min at room temperature (RT), samples and standards (each volme 100 μL) were added to appropriate wells for 2 h. Then, the secondary antibodies (100 μL) were dispensed into each well and left for 2 h. After washing thrice, 3,3′,5,5′-tetramethylbenzidine (TMB) substrate 100 μL was applied. Absorbance levels were measured at 405 nm and were recorded to analyze the effect of GGW73.
2.8. Western Blot Analysis
As mentioned previously, hippocampal tissue was obtained at the end of the experiment. The hippocampus (10 mg) was homogenized with PRO-PREP solution (iNtRON Biotech, Seongnam, Gyeonggi-do, Republic of Korea) at 5500× g for 1 h using a Precellys 24 tissue homogenizer. The homogenized tissues were centrifuged for 10 min at 4 °C, 12,879× g and the supernatants were frozen in a deep freezer (−75 °C) before the experiment. The supernatant was then moved into a 1.7 mL sterile tube. To quantify proteins, the DC Protein Assay Kit Ⅱ (Bio-Rad Laboratories, Hercules, CA, USA) was used.
For electrophoresis, the protein samples (8 μg) were loaded onto SDS-PAGE. The transferred membranes, made of polyvinylidene fluoride, were blocked with 5% TBSB (BSA in 1 xTBS at 5% (w/v)) for 2 h, then membranes were washed with wash buffer (1 xTBS containing tween-20 at 0.05% (v/v)) at 30 min intervals for 1.5 h. The transferred membranes were incubated with specific antibodies in a refrigerator at 4 °C overnight. The antibody-coated membranes were washed with wash buffer at intervals of 10 min for 2 h. The secondary antibodies and original antibody were diluted with sterile distilled water ten times, and were then added to the membranes and reacted for 1.5 h. Then, the membranes were washed with wash buffer at 10-min intervals for 2 h. Finally, the membranes were coated with Western blotting luminol reagent (Santa Cruz, CA, USA) and measured with a LuminoGraph III Lite (ATTO, Tokyo, Japan).
2.9. Statistical Analysis
GraphPad Prism (ver. 5.0) was utilized for statistical analysis of all the experimental values. The experimental values were presented as the mean ± standard error of the mean (S.E.M.). A statistical assessment was conducted using Tukey’s post hoc analysis and one-way analysis of variance analysis (ANOVA). p-values of <0.05 were regarded as significant.
3. Results
3.1. Effect of GGW73 on the Levels of Serum AST, ALT, BUN and Creatinine
To evaluate the effect of GGW73 on metabolic toxicity, serum ALT, AST, BUN, and creatinine levels were detected. As a result of the administration of various concentrations of GGW73 for 14 days, no significant changes were found in serum AST, ALT, BUN, or creatinine levels compared to the non-treatment group (Figure 1A–D). However, oral administration of 5000 mg/kg GGW73 slightly increased serum BUN.
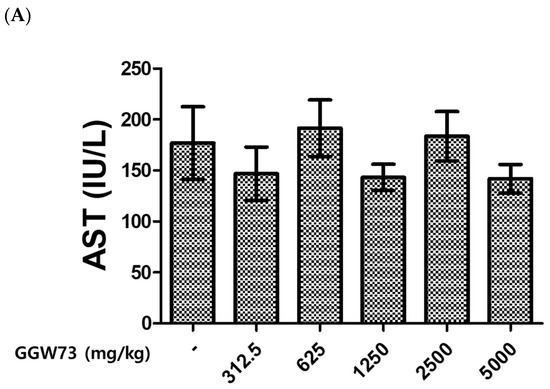
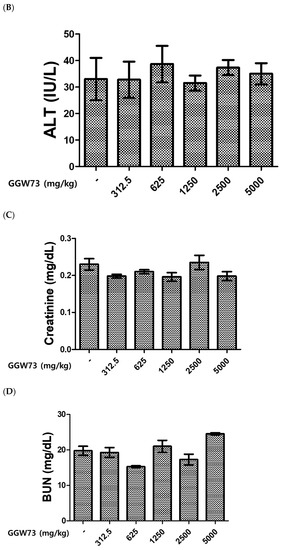
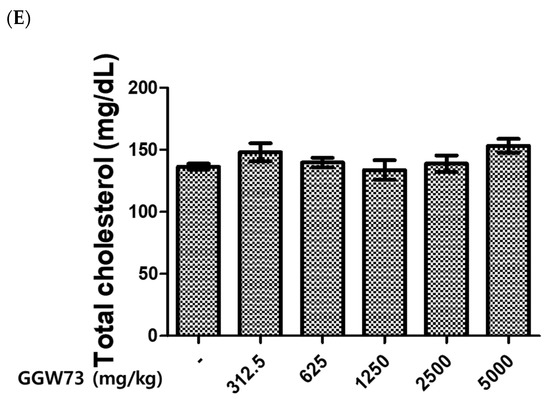
Figure 1.
Effect of GGW73 on the levels of serum AST, ALT, BUN, and creatinine. We analyzed the serum of mice (n = 5) which were administered GGW73 for 14 days. Bars represent (A) AST, (B) ALT, (C) BUN, (D) creatinine, (E) total cholesterol in serum. All values are represented as the mean ± S.E.M.
3.2. Effect of GGW73 on Y-Maze Test Performance in Scopolamine-Induced Cognitive Disorder Mice
The efficacy of GGW73 in scopolamine-induced cognitive disorders was evaluated through the quantification of total arm entries and alternation triplets of the Y-maze test. Total arm entries were significantly increased by the intraperitoneally scopolamine injection (1 mg/kg) (Figure 2A). Increased entries, extracts, and donepezil tended to slightly decrease as a positive control. In particular, the administration of GGW 73 100 mg/kg significantly reduced the total arm entries (Figure 2A). In addition, 100 mg/kg GGW73 administration significantly upregulated the percentage of alternation arm triplets compared with the scopolamine-only treated group (Figure 2B).
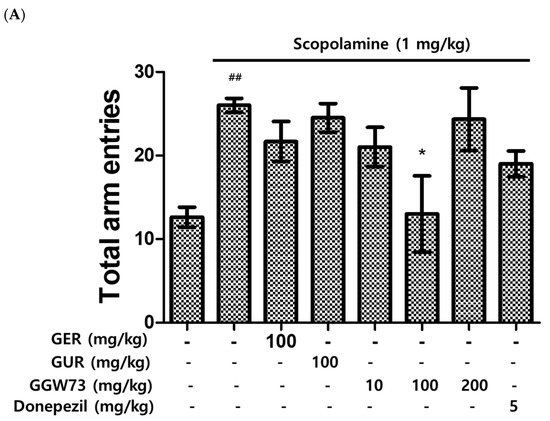
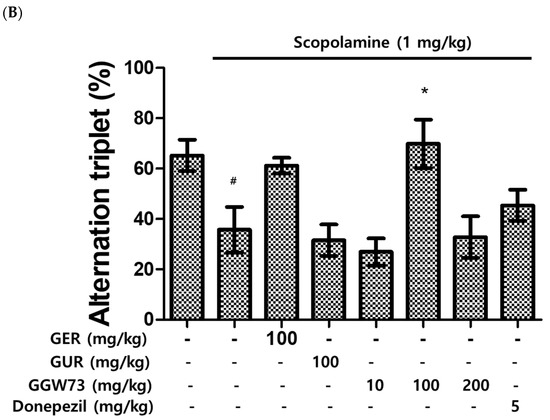
Figure 2.
Effect of GGW73 on Y-maze test performance in scopolamine-induced cognitive disorder mice. Shown are the (A) total arm entries, and (B) alternation triplets. All values are represented as the mean ± S.E.M. # p < 0.05 compared with only saline treatment group; ## p < 0.005 only saline treatment group; * p < 0.05 compared with only scopolamine treatment group.
3.3. Effect of GGW73 on Water-Maze Test Performance in Scopolamine-Induced Cognitive Disorder Mice
The water maze test was carried out to measure the influence of scopolamine and GGW73 in vivo. The scopolamine (1 mg/kg) intraperitoneal injection elevated the latency to locate the platform. However, 100 mg/kg GUR extract ameliorated the latency to target scopolamine-induced cognitive function disorder in mice. With GGW73 administration, a decreasing trend was observed, but the difference was not statistically significant (Figure 3A). The swimming percentage of time spent in the pool was increased by the scopolamine injection. GUR extract (100 mg/kg) significantly decreased the time spent percentage in the pool. GGW73 and donepezil administration had no effect on the scopolamine-induced time spent in the pool (Figure 3B). Similarly, the percentage of time spent on the platform following scopolamine injection was significantly increased by GUR or 10 mg/kg GGW73 treatments (Figure 3C). However, GGW73 (100 mg/kg) and donepezil administration did not show any significant changes.
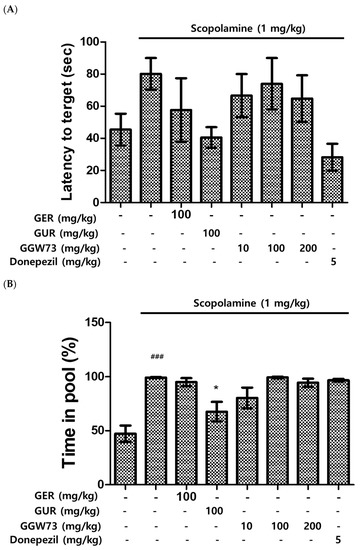
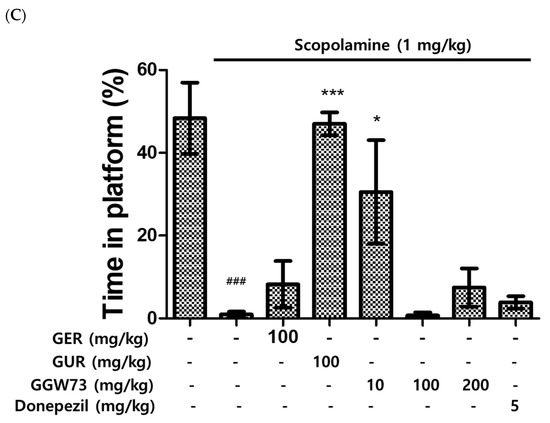
Figure 3.
Effect of GGW73 on water-maze test scores in scopolamine-induced cognitive disorder mice. The water maze test was performed to evaluate cognitive function. (A) latency to target, (B) time in pool, (C) time in platform. All values are represented as the mean ± S.E.M. ### p < 0.001 compared with only saline treatment group; * p < 0.05 compared with only scopolamine-treatment group; *** p < 0.001 compared with only scopolamine treatment group.
3.4. Effect of GGW73 on the Passive Avoidance Test in Scopolamine-Induced Cognitive Disorder Mice
To further examine the effect of GGW73 on scopolamine-induced cognitive disorders, we performed a passive avoidance test in vivo. Latency, which indicates impairment of cognition, was significantly shorter in the scopolamine injection group than in the non-treatment group. The experimental groups administered GER 100 mg/kg, GUR 100 mg/kg, GGW 10, and GGW 100 mg/kg showed improved latency compared with the scopolamine-only injection group (Figure 4). These results indicated that GGW regulates scopolamine-induced cognitive function disorders.
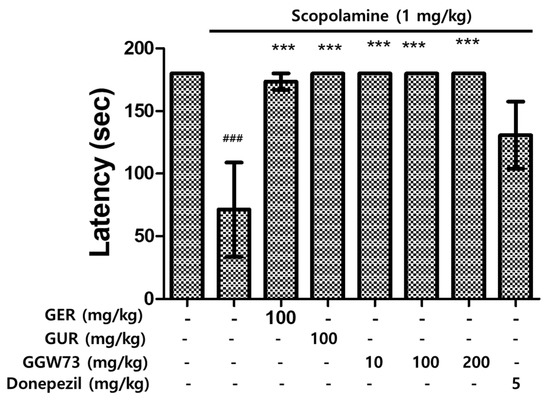
Figure 4.
Effect of GGW73 on the passive avoidance test in scopolamine-induced cognitive disorder mice. Shown is the latency of avoidance time. All values are represented as the mean ± S.E.M. ### p < 0.001 compared with only saline treatment group; *** p < 0.001 compared with only scopolamine treatment group.
3.5. Effect of GGW73 on the Hippocampal Amyloid-Beta Pathway in Scopolamine-Induced Cognitive Disorder Mice
To estimate the effect of GGW73 on the Aβ-related pathway, Western blot analysis was performed using the total hippocampal protein. We found that scopolamine (1 mg/kg) diminished BDNF expression. However, oral administration of extracts containing GGW73 (100 mg/kg) restored the diminished BDNF expression. In addition, APP phospholyration and BACE-1 exression were slightly increased following scopolamine injection. GGW73 100 mg/kg administration downregulated the enhanced APP phospholyration and BACE-1 expression in the hippocampus (Figure 5).
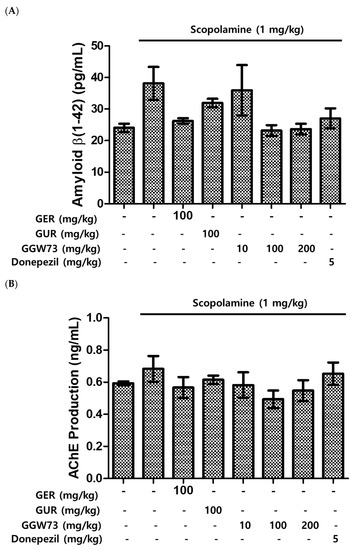
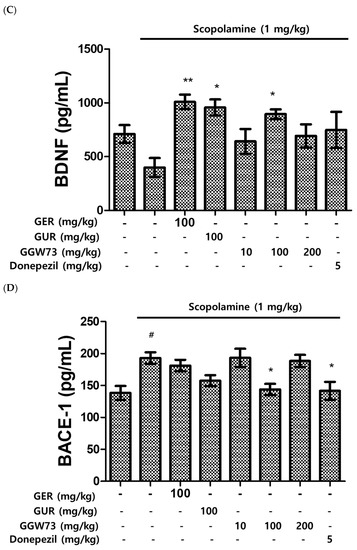
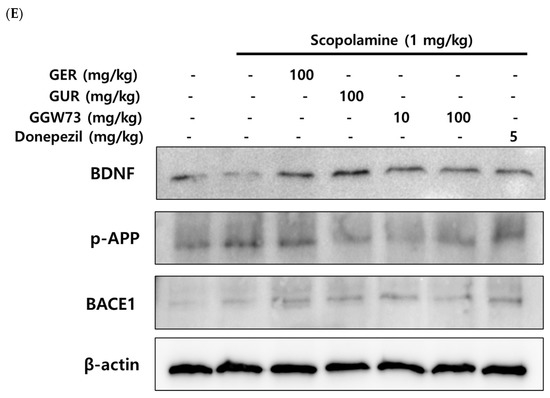
Figure 5.
Effect of GGW73 on the amyloid-beta pathway in the hippocampus of scopolamine-induced cognitive disorder mice. The total hippocampus was used to measure protein concentrations. Shown are the expressions of (A) β-amyloid, (B) acetylcholinesterase, (C) BDNF, (D) BACE-1, and (E) the of BDNF, p-APP, and BACE1, β-actin, as the loading control, protein expressions. All values are represented as the mean ± S.E.M. # p < 0.05 compared with only saline treatment group; * p < 0.05 compared with only scopolamine-treatment group; ** p < 0.005 compared with only scopolamine treatment group.
3.6. Effect of GGW73 on Hippocampal Injury in Scopolamine-Induced Cognitive Disorder Mice
To observe the influence of scopolamine and GGW73, H&E staining was performed on hippocampal tissues. Only the scopolamine-injected group showed condensed neuronal cells (red arrows) in the hippocampal tissue compared with the non-treatment group, which had a distinctly round nucleus and maintained its structure. However, GGW73 100 mg/kg administration slightly recovered the condensed neurons in CA (1–3) regions of the hippocampus (Figure 6). These results indicate that GGW73 administration can prevent hippocampal injury.
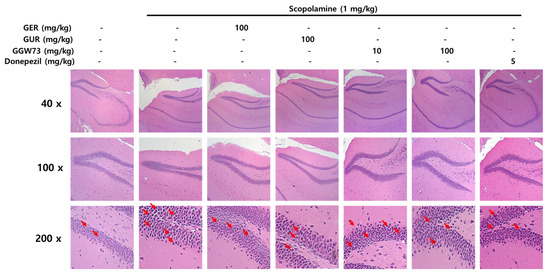
Figure 6.
Effect of GGW73 on hippocampal injury in scopolamine-induced cognitive disorder mice. Slices were stained with hematoxylin and eosin. The hippocampus was observed with a standard microscope at 40×, 100×, 200× magnification.
4. Discussion
GE, belonging to the Orchidaceae family [50], has been shown to be effective in the cardiovascular system [51], convulsions [52], skin aging [53], and cancer cell invasion [54]. Above all, the beneficial neurological effects of GE extract and its constituents, especially gastrodin, have been revealed in Aβ-induced neuronal apoptosis [23], mice with pharmacologically induced memory impairments [55], and in a mouse model of AD [56]. GU belongs to the Leguminosae family [57] and has been determined to have memory enhancing [58], neuroprotective [48], and cerebral infarction modulative [59] effects. Our previous study demonstrated that GGW73 in mixtures of various proportion ratios (GER: GUR = 5:5, 6:4, 7:3, 8:2, and 9:1) had neuroprotective effects on Aβ stimulation in vitro [60]. Based on our previous results, we chose to use GGW73 for scopolamine-induced cognitive disorders in vivo.
The Y-maze test is an experimental tool for measuring cognitive function in scopolamine-treated mice in vivo [61]. The Y-maze test, which uses the propensity of rodents to enter an unfamiliar place, is a widely used method to estimate the extent of memory impairment in cognitive disorders, such as AD and memory deficiency [62]. The total arm entries were significantly decreased by GGW73 100 mg/kg oral administration compared with scopolamine 1 mg/kg peritoneal injection (non-treatment group:12.60 ± 1.21, scopolamine group: 26.00 ± 0.84, GGW73 100 mg/kg group: 13.00 ± 4.56, donepezil group: 19.00 ± 1.53). Additionally, we observed decreased alternation triplets after scopolamine treatment in the Y-maze test and elevated alternation triplets by GEW 100 mg/kg and GGW73 100 mg/kg administration (non-treatment group: 65.22 ± 6.22%, scopolamine 1 mg/kg group: 35.72 ± 9.06%, GER 100 mg/kg group: 61.21 ± 3.11%, GGW73 100 mg/kg group: 69.84 ± 9.65%, donepezil group: 45.39 ± 6.19%). These results indicated that oral administration of GGW73 (100 mg/kg) attenuated scopolamine-induced cognitive disorder. The water maze test has been representatively performed to evaluate memory and cognition in behavioral neuroscience [63]. The latency to reach the platform was reduced by GUR 100 mg/kg administration compared to scopolamine injection. Oral administration of GGW73 tended to shorten the latency, but no significant difference was observed (non-treatment group: 45.42 ± 9.99 s, scopolamine group: 80.16 ± 9.85 s, GUR 100 mg/kg group: 40.55 ± 6.43 s, GGW73 100 mg/kg group: 64.79 ± 14.49 s, donepezil group: 25.28 ± 8.363 s) (Figure 3A). The time spent swimming in the maze and platform ratios were also significantly regulated by GUR 100 mg/kg; however, oral administration of GGW73 100 mg/kg also showed a slight regulatory effect (Figure 3B,C). The mice learned to recognize aversive stimuli, thereby enhancing cognitive function in the passive avoidance test [64]. Therefore, the passive avoidance test is the most widely used in vivo experiment for evaluating cognitive function disorder, memory deficient [65]. The scopolamine 1 mg/kg intraperitoneal injection decreased the latency to target (non-treatment group: 180.0 ± 0.0 s, scopolamine group: 71.4 ± 37.6 s); however, GGW73 administration significantly recovered the latency to target (Figure 4). The results showed that administration of GGW73 has the potential to remedy scopolamine-induced cognitive impairment.
The accumulation of Aβ peptides is related to cognitive function. In cognitive disorders, the expression level of Aβ is elevated by a decrease in amyloid precursor protein (APP) [66]. Scopolamine contributes to increased Aβ release, which is influenced by APP [67]. The level of Aβ peptide was elevated by scopolamine injection (non-treatment group:24.06 ± 1.30 pg/mL, scopolamine group:38.14 ± 5.23 pg/mL). However, the increased expression of Aβ was downregulated by oral administration of GGW73 (100 and 200 mg/kg) in hippocampal tissues (GGW73 100 mg/kg group: 23.17 ± 1.74 pg/mL, GGW73 200 mg/kg group: 23.67 ± 1.69 pg/mL) (Figure 5A). The experimental results indicate that GGW73 could regulate cognitive disorders by diminishing excessive Aβ release in hippocampal tissues.
Loss of cognitive function causes ACh deficiency in the hippocampus and cortex [12]. For this reason, a strategy to inhibit AChE in the hippocampus improves cognitive function disorders [68]. The increased AChE expression in hippocampal tissue by scopolamine was reduced by oral administration of GGW73 100 mg/kg (non-treatment group: 0.593 ± 0.012 ng/mL, scopolamine group: 0.683 ± 0.080 ng/mL, GGW73 100 mg/kg group: 0.493 ± 0.054 ng/mL, donepezil group:0.653 ± 0.069 ng/mL) (Figure 5B). Among various neuronal enzymes, β- and γ-secretase sequentially lyse APP proteins, which generate Aβ [69]. Furthermore, over-expression of β-secretase (BACE-1) and Aβ deposition has been detected in degenerative neuronal diseases such as AD and cognitive disorders [70]. As a muscarinic antagonist, scopolamine influences Aβ peptide release [67] and BACE-1 expression in the hippocampus [71,72]. GGW73 100 mg/kg oral administration ameliorated BACE-1 expression in scopolamine-injected mice hippocampus tissue (non-treatment group: 138.4 ± 10.9 pg/mL, scopolamine group: 192.9 ± 9.1 pg/mL, GGW73 100 mg/kg group: 143.9 ± 8.8 pg/mL, GGW73 200 mg/kg group: 188.4 ± 9.6 pg/mL, donepezil group: 141.6 ± 14.2 pg/mL) (Figure 5D). These results indicate that GGW73 has a regulatory effect on scopolamine-induced cognitive disorders.
Synaptic plasticity is modulated by BDNF in the human brain [73]. Scopolamine influences neuronal plasticity by reducing BDNF levels in the mouse cerebrum [74] and hippocampus [75]. The decreased BDNF expression by scopolamine injection was significantly recovered by GGW73 oral administration in the hippocampus (non-treatment group: 710.5 ± 82.1 pg/mL, scopolamine group: 398.7 ± 88.0 pg/mL, GER 100 mg/kg group: 1009.1 ± 67.54 pg/mL, GGW73 100 mg/kg group: 895.3 ± 44.68 pg/mL, donepezil group: 748.3 ± 168.0 pg/mL) (Figure 5C). The results showed that GGW73 restored scopolamine-induced hippocampal injury. In Western blot analysis, GGW73 oral administration slightly regulated BDNF and p-APP, but not in a dose-dependent manner (Figure 5E). However, these results showed GGW73 could modulate the signaling pathway in scopolamine-induced cognitive disorder. In addition, neurons of the hippocampus were condensed by scopolamine injection compared to those in the non-treatment group, which had a distinctly round nucleus and a constant structure. However, condensed neurons in the hippocampus were slightly recovered by GGW73 (100 mg/kg) administration (Figure 6).
5. Conclusions
The present results showed an ameliorative effect of GGW73 on behavioral tests, such as the Y-maze, water maze, and passive avoidance test in mice with scopolamine-induced cognitive disorder. We also investigated whether GGW73 administration attenuated the Aβ-related pathway by regulating the expression levels of Aβ peptide, AChE, BACE-1, and BDNF in the hippocampus. These results indicate that GGW73 could be used as an alternative medical remedy for cognitive disorders.
Author Contributions
J.-H.L., Y.-D.J., D.-K.K. and J.-W.L. conceptualized and designed the experiments. S.-W.K., S.-H.L., S.-H.H., J.-Y.C. and W.-W.L. performed the experiments. J.-H.L., Y.-D.J., S.-W.K. and S.-H.L. analyzed the experimental data. J.-H.L., Y.-D.J. and D.-K.K. prepared the manuscript. J.-W.L. and D.-K.K. edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the Ministry of Trade, Industry, and Energy (MOTIE) and the Korea Institute for Advancement of Technology (KIAT) through the National Innovation Cluster R&D Program (P0016236). This research was also supported by a grant from the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health and Welfare, Republic of Korea (grant number: HF22C0106).
Institutional Review Board Statement
All experiments were conducted in accordance with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals, and the Institutional Animal Care and Use Committee at Woosuk University approved this study (approval ID: WS2020-08).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Thanks for contribution of M.-R.P., CEO of MJ HEALTH FOOD Co., Ltd. (Muju-gun, Jeollabuk-do, Republic of Korea). M.-R.P. provided GE and GU to research team and conceptualized in vivo experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Im, H.I.; Lee, E.S.; Hwang, Y.J.; Kim, Y.S. Baicalein protects 6-OHDA-induced neuronal damage by suppressing oxidative stress. Korean J. Physiol. Pharmacol. 2006, 10, 309–315. [Google Scholar]
- More, S.V.; Kumar, H.; Cho, D.-Y.; Yun, Y.-S.; Choi, D.-K. Toxin-Induced Experimental Models of Learning and Memory Impairment. Int. J. Mol. Sci. 2016, 17, 1447. [Google Scholar] [CrossRef]
- Asuntha, G.; Prasannaraju, Y.; Prasad, K.V.S.R.G. Effect of ethanol extract of Indigofera tinctoria Linn. (Fabaceae) on lithium/pilocarpine-induced status epilepticus and oxidative stress in Wistar rats. Trop. J. Pharm. Res. 2010, 9, 149–156. [Google Scholar] [CrossRef]
- Di Paolo, G.; Kim, T.-W. Linking lipids to Alzheimer ’s disease: Cholesterol and beyond. Nat. Rev. Neurosci. 2011, 12, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, L.H.; Langa, K.M.; Bynum, J.P.; Hsu, J.W. Financial presentation of Alzheimer disease and related dementias. JAMA Intern Med. 2021, 181, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chang, L.; Song, Y.; Li, H.; Wu, Y. The role of NMDA receptors in Alzheimer’s disease. Front. Neurosci. 2019, 13, 43. [Google Scholar] [CrossRef]
- Hashimoto, M.; Kazui, H.; Matsumoto, K.; Nakano, Y.; Yasuda, M.; Mori, E. Does Donepezil Treatment Slow the Progression of Hippocampal Atrophy in Patients With Alzheimer’s Disease? Am. J. Psychiatry 2005, 162, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Modrego, P.J.; Fayed, N.; Rios, C.; Pina, M.A.; Sarasa, M. Memantine versus donepezil in mild to moderate Alzheimer’s disease: A randomized trial with magnetic resonance spectroscopy. Eur. J. Neurol. 2010, 17, 405–412. [Google Scholar] [CrossRef]
- Kasa, P.; Papp, H.; Kasa, P., Jr.; Torok, I. Donepezil dose-dependently inhibits acetylcholinesterase activity in various areas and in the presynaptic cholinergic and the postsynaptic cholinoceptive enzyme-positive structures in the human and rat brain. Neuroscience 2000, 101, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Almeida, S.; Filipe, A.; Neves, R.; Desjardins, I.; Shink, E.; Castillo, A. Bioequivalence study of two different tablet formulations of donepezil using truncated areas under the curve. A single-center, single-dose, randomized, open-label, 2-way crossover study under fasting conditions. Arzneimnittelforschung 2010, 60, 116–123. [Google Scholar]
- Aryannejad, A.; Gandominejad, A.; Tabary, M.; Noroozi, N.; Abbasi, A.; Araghi, F.; Jafari, R.M.; Dehpour, A.R. Protective effect of modafinil on skin flap survival in the experimental random-pattern skin flap model in rats: The role of ATP-sensitive potassium channels and nitric oxide pathway. J. Plast. Reconstr. Aesthet. Surg. 2021, 74, 1346–1354. [Google Scholar] [CrossRef]
- Schliebs, R.; Arendt, T. The cholinergic system in aging and neuronal degeneration. Behav. Brain Res. 2011, 221, 555–563. [Google Scholar] [CrossRef]
- Aykac, A.; Ozbeyli, D.; Uncu, M.; Ertas, B.; Kılınc, O.; Sen, A.; Orun, O.; Sener, G. Evaluation of the protective effect of Myrtus communis in scopolamine-induced Alzheimer model through cholinergic receptors. Gene 2019, 689, 194–201. [Google Scholar] [CrossRef]
- Karthivashan, G.; Park, S.-Y.; Kweon, M.-H.; Kim, J.; Haque, M.; Cho, D.-Y.; Kim, I.-S.; Cho, E.-A.; Ganesan, P.; Choi, D.-K. Ameliorative potential of desalted Salicornia europaea L. extract in multifaceted Alzheimer ’s-like scopolamine-induced amnesic mice model. Sci. Rep. 2018, 8, 7174. [Google Scholar] [CrossRef]
- Richetti, S.; Blank, M.; Capiotti, K.; Piato, A.; Bogo, M.; Vianna, M.; Bonan, C. Quercetin and rutin prevent scopolamine-induced memory impairment in zebrafish. Behav. Brain Res. 2011, 217, 10–15. [Google Scholar] [CrossRef]
- Zhao., L.; Sun, C.; Xiong, L.; Yang, Y.; Gao, Y.; Wang, L.; Zuo, H.; Xu, X.; Dong, J.; Zhou, H.; et al. MicroRNAs: Novel mechanism involved in the pathogenesis of microwave exposure on rats’ hippocampus. J. Mol. Neurosci. 2014, 53, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Terry, A.V., Jr.; Callahan, P.M.; Hall, B.; Webster, S.J. Alzheimer’s disease and age-related memory decline (preclinical). Pharmacol. Biochem. Behav. 2011, 99, 190–210. [Google Scholar] [CrossRef] [PubMed]
- Gold, P.E. Acetylcholine modulation of neural systems involved in learning and memory. Neurobiol. Learn Mem. 2003, 80, 194–210. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.J.; Shou, H.; Benzinger, T.; Marcus, D.; Durbin, T.; Morris, J.C.; Sheline, Y.I. Amyloid burden in cognitively normal elderly is associated with preferential hippocampal subfield volume loss. J. Alzheimer’s Dis. 2015, 45, 27–33. [Google Scholar] [CrossRef]
- Kaur, R.; Parveen, S.; Mehan, S.; Khanna, D.; Kalra, S. Neuroprotective effect of ellagic acid against chronically scopolamine induced Alzheimer’s type memory and cognitive dysfunctions: Possible behavioural and biochemical evidences. Int. J. Preven. Med. Res. 2015, 1, 45–64. [Google Scholar]
- Mohs, R.C. A perspective on risks that impede development of drugs to modify the course of Alzheimer’s disease: Can they be reduced? Alzheimer’s Dement. 2008, 4 (Suppl. S1), S85–S87. [Google Scholar] [CrossRef] [PubMed]
- Green, K.N.; Billings, L.M.; Roozendaal, B.; McGaugh, J.L.; LaFerla, F.M. Glucocorticoids increase amyloid-beta and tau pathology in a mouse model of Alzheimer’s disease. J. Neurosci. 2006, 26, 9047–9056. [Google Scholar] [CrossRef]
- Park, Y.M.; Lee, B.G.; Park, S.H.; Oh, H.G.; Kang, Y.G.; Kim, O.J.; Kwon, L.S.; Kim, Y.P.; Choi, M.H.; Jeong, Y.S.; et al. Prolonged oral administration of Gastrodia elata extract improves spatial learning and memory of scopolamine-treated rats. Lab. Anim. Res. 2015, 31, 69–77. [Google Scholar] [CrossRef] [PubMed]
- El Hayek, L.; Khalifeh, M.; Zibara, V.; Abi Assaad, R.; Emmanuel, N.; Karnib, N.; El-Chandour, R.; Nasrallah, P.; Bilen, M.; Ibrahim, P.; et al. Lactate mediates the effects of exercise on learning and memory through sirt1- dependent activation of hippocampal brain-derived neurotrophic factor (BDNF). J. Neurosci. 2019, 39, 2369–2382. [Google Scholar] [CrossRef]
- Dong, B.E.; Chen, H.; Sakata, K. BDNF deficiency and enriched environment treatment affect neurotransmitter gene expression differently across ages. J. Neurochem. 2020, 154, 41–55. [Google Scholar] [CrossRef]
- Auld, D.S.; Mennicken, F.; Day, J.C.; Quirion, R. Neurotrophins differentially enhance acetylcholine release, acetylcholine content and choline acetyltransferase activity in basal forebrain neurons. J. Neurochem. 2001, 77, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Hritcu, L.; Cioanca, O.; Hancianu, M. Effects of lavender oil inhalation on improving scopolamine-induced spatial memory impairment in laboratory rats. Phytomedicine 2012, 19, 529–534. [Google Scholar] [CrossRef]
- Aarsland, D.; Mosimann, U.P.; McKeith, I.G. Role of cholinesterase inhibitors in Parkinson’s disease and dementia with Lewy bodies. J. Geriatr. Psychiatr. Neurol. 2004, 17, 164–171. [Google Scholar] [CrossRef]
- Roberds, S.L.; Anderson, J.; Basi, G.; Bienkowski, M.J.; Branstetter, D.G.; Chen, K.S.; Freedman, S.B.; Frigon, N.L.; Games, D.; Hu, K.; et al. BACE knockout mice are healthy despite lacking the primary beta-secretase activity in brain: Implications for Alzheimer’s disease therapeutics. Hum. Mol. Genet. 2001, 10, 1317–1324. [Google Scholar] [CrossRef]
- Zhan, H.-D.; Zhou, H.-Y.; Sui, Y.-P.; Du, X.-L.; Wang, W.-H.; Dai, L.; Sui, F.; Huo, H.R.; Jiang, T.L. The rhizome of Gastrodia elata Blume—An ethnopharmacological review. J. Ethnopharmacol. 2016, 189, 361–385. [Google Scholar] [CrossRef]
- Duan, X.H.; Li, Z.L.; Yang, D.S.; Zhang, F.L.; Lin, Q.; Dai, R. Study on the chemical constituents of Gastrodia elata. Zhong Yao Cai 2013, 36, 1608–1611. [Google Scholar]
- Tao, J.; Luo, Z.; Msangi, C.I.; Shu, X.; Wen, L.; Liu, S.; Zhou, C.; Liu, R.; Hu, W. Relationships among genetic makeup, active ingredient content, and place of origin of the medicinal plant Gastrodia tuber. Biochem. Genet. 2009, 47, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Choong, L.C.; Panyod, S.; Lin, Y.E.; Huang, H.S.; Lu, K.H.; Wu, W.K.; Sheen, L.Y. Gastrodia elata Blume water extract modulates neurotransmitters and alters the gut microbiota in a mild social defeat stress-induced depression mouse model. Phytother. Res. 2021, 35, 5133–5142. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, J.X.; Yan, C.; Liu, Y.; Liu, H.; Li, D.; Zhu, J.; Luo, Z.B.; Han, S.Z.; Jin, Z.Y.; et al. Gastrodia elata Blume extract improves high-fat diet-induced type 2 diabetes by regulating gut microbiota and bile acid profile. Front. Microbiol. 2022, 13, 1091712. [Google Scholar] [CrossRef]
- Park, S.; Kim, D.S.; Kang, S. Gastrodia elata Blume water extracts improve insulin resistance by decreasing body fat in diet-induced obese rats: Vanillin and 4-hydroxybenzaldehyde are the bioactive candidates. Eur. J. Nutr. 2011, 50, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.M.; Lee, Y.J.; Kang, D.G.; Lee, H.S. Anti-inflammatory effect of Gastrodia elata rhizome in human umbilical vein endothelial cells. Am. J. Chin. Med. 2009, 37, 395–406. [Google Scholar] [CrossRef]
- Kim, N.H.; Xin, M.J.; Cha, J.Y.; Ji, S.J.; Kwon, S.U.; Jee, H.K.; Park, M.R.; Park, Y.S.; Kim, C.T.; Kim, D.K.; et al. Antitumor and Immunomodulatory Effect of Gastrodia elata on Colon Cancer In Vitro and In Vivo. Am. J. Chin. Med. 2017, 45, 319–335. [Google Scholar] [CrossRef]
- Tan, W.; Zheng, Q.; Feng, K.; Feng, X.; Zhong, W.; Liao, C.; Li, S.; Liu, Y.; Hu, W. Neuroprotection of Gastrodia elata polyphenols against H2O2-induced PC12 cell cytotoxicity by reducing oxidative stress. Front. Pharmacol. 2022, 10, 1050775. [Google Scholar] [CrossRef] [PubMed]
- Doo, A.R.; Kim, S.N.; Hahm, D.H.; Yoo, H.H.; Park, J.Y.; Lee, H.; Jeon, S.; Kim, J.; Park, S.U.; Park, H.J. Gastrodia elata Blume alleviates L-DOPA-induced dyskinesia by normalizing FosB and ERK activation in a 6-OHDA-lesioned Parkinson’s disease mouse model. BMC Complement Altern. Med. 2014, 20, 107. [Google Scholar] [CrossRef]
- Huang, G.B.; Zhao, T.; Muna, S.S.; Jin, H.M.; Park, J.I.; Jo, K.S.; Lee, B.H.; Chae, S.W.; Kim, S.Y.; Park, S.H.; et al. Therapeutic potential of Gastrodia elata Blume for the treatment of Alzheimer’s disease. Neural. Regen. Res. 2013, 8, 1061–1070. [Google Scholar]
- Mishra, M.; Huang, J.; Lee, Y.Y.; Chua, D.S.K.; Lin, X.; Hu, J.M.; Heese, K. Gastrodia elata modulates amyloid precursor protein cleavage and cognitive functions in mice. Biosci. Trends 2011, 5, 129–138. [Google Scholar] [CrossRef]
- Mochida, K.; Sakurai, T.; Seki, H.; Yoshida, T.; Takahagi, K.; Sawai, S.; Uchiyama, H.; Muranaka, T.; Saito, K. Draft genome assembly and annotation of Glycyrrhiza uralensis, a medicinal legume. Plant J. 2017, 89, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Asl, M.N.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother. Res. 2008, 22, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Oh, S.M.; Kwon, H.S.; Oh, Y.S.; Lim, S.S.; Shin, H.K. Anti-inflammatory effect of roasted licorice extracts on lipopolysaccharide-induced inflammatory responses in murine macrophages. Biochem. Biophys. Res. Commun. 2006, 345, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Oganesyan, K.R. Antioxidant effect of licorice root on blood catalase activity in vibration stress. Bull. Exp. Biol. Med. 2002, 134, 135–136. [Google Scholar] [CrossRef]
- Fiore, C.; Eisenhut, M.; Krausse, R.; Ragazzi, E.; Pellati, D.; Armanini, D.; Bielenberg, J. Antiviral effects of glycyrrhiza species. Phytother. Res. 2008, 22, 141–148. [Google Scholar] [CrossRef]
- Ramachandran, B.; Srinivasadesikan, V.; Chou, T.M.; Jeyakanthan, J.; Lee, S.L. Atomistic simulation on flavonoids derivatives as potential inhibitors of bacterial gyrase of Staphylococcus aureus. J. Biomol. Struct. Dyn. 2022, 40, 4314–4327. [Google Scholar] [CrossRef]
- Lee, H.K.; Yang, E.J.; Kim, J.Y.; Song, K.; Seong, Y.H. Inhibitory effects of Glycyrrhizae radix and its active component, isoliquiritigenin, on Aβ(25-35)-induced neurotoxicity in cultured rat cortical neurons. Arch. Pharm. Res. 2012, 35, 897–904. [Google Scholar] [CrossRef]
- Dhingra, D.; Parle, M.; Kulkarni, S.K. Memory enhancing activity of Glycyrrhiza glabra in mice. J. Ethnopharmacol. 2004, 91, 361–365. [Google Scholar] [CrossRef]
- Ding, X.; Liu, Z.; Liu, Y.; Xu, B.; Chen, J.; Pu, Z.; Wu, D.; Yu, H.; Jin, C.; Wang, X. Comprehensive evaluation of the mechanism of Gastrodia elata Blume in ameliorating cerebral ischemia-reperfusion injury based on integrating fecal metabonomics and 16S rDNA sequencing. Front. Cell Infect. Microbiol. 2022, 12, 1026627. [Google Scholar]
- Kim, M.J.; Yang, H.J.; Moon, B.R.; Kim, J.E.; Kim, K.S.; Park, S. Gastrodia elata Blume Rhizome Aqueous Extract Improves Arterial Thrombosis, Dyslipidemia, and Insulin Response in Testosterone-Deficient Rats. Evid. Based Complem. Altern. Med. 2017, 2017, 2848570. [Google Scholar] [CrossRef]
- Hsieh, C.L.; Chiang, S.Y.; Cheng, K.S.; Lin, Y.H.; Tang, N.Y.; Lee, C.J.; Pon, C.Z.; Hsieh, C.T. Anticonvulsive and free radical scavenging activities of Gastrodia elata Bl. in kainic acid-treated rats. Am. J. Chin. Med. 2001, 29, 331–341. [Google Scholar] [CrossRef]
- Song, E.; Chung, H.; Shim, E.; Jeong, J.K.; Han, B.K.; Choi, H.J.; Hwang, J. Gastrodia elata Blume Extract Modulates Antioxidant Activity and Ultraviolet A-Irradiated Skin Aging in Human Dermal Fibroblast Cells. J. Med. Food 2016, 19, 1057–1064. [Google Scholar] [CrossRef]
- Heo, J.C.; Woo, S.U.; Son, M.; Park, J.Y.; Choi, W.S.; Chang, K.T.; Kim, S.U.; Yoon, E.K.; Kim, Y.H.; Shin, H.M.; et al. Anti-tumor activity of Gastrodia elata Blume is closely associated with a GTP-Ras-dependent pathway. Oncol. Rep. 2007, 18, 849–853. [Google Scholar] [CrossRef]
- Huang, H.; Jiang, N.; Zhang, Y.W.; Lv, J.W.; Wang, H.X.; Lu, C.; Liu, X.M.; Lu, G.H. Gastrodia elata blume ameliorates circadian rhythm disorder-induced mice memory impairment. Life Sci. Space Res. 2021, 31, 51–58. [Google Scholar] [CrossRef]
- Ding, Y.; Bao, X.; Lao, L.; Ling, Y.; Wang, Q.; Xu, S. p-Hydroxybenzyl alcohol prevents memory deficits by increasing neurotrophic factors and decreasing inflammatory factors in a mice model of Alzheimer’s disease. J. Alzheimer’s Dis. 2019, 67, 1007–1019. [Google Scholar] [CrossRef]
- Sato, Y.; He, J.X.; Nagai, H.; Tani, T.; Akao, T. Isoliquiritigenin, one of the antispasmodic principles of Glycyrrhiza ularensis roots, acts in the lower part of intestine. Biol. Pharm. Bull. 2007, 30, 145–149. [Google Scholar] [CrossRef]
- Ahn, J.; Um, M.; Choi, W.; Kim, S.; Ha, T. Protective effects of Glycyrrhiza uralensis Fisch. on the cognitive deficits caused by β-amyloid peptide 25-35 in young mice. Biogerontology 2006, 7, 239–247. [Google Scholar] [CrossRef]
- Barakat, W.; Safwet, N.; El-Maraghy, N.N.; Zakaria, M.N.M. Candesartan and glycyrrhizin ameliorate ischemic brain damage through downregulation of the TLR signaling cascade. Eur. J. Pharmacol. 2014, 724, 43–50. [Google Scholar] [CrossRef]
- Hwang, S.H.; Park, S.B.; Yu, D.Y.; Cho, J.Y.; Lee, W.W.; Park, M.R.; Lee, J.W.; Jeon, Y.D. Neuroprotective Effect of the Mixture of Gastrodiae elata and Glycyrrhizae uralensis In Vitro. Appl. Sci. 2023, 13, 190. [Google Scholar] [CrossRef]
- Van der Borght, K.; Havekes, R.; Bos, T.; Eggen, B.J.; Van der Zee, E.A. Exercise improves memory acquisition and retrieval in the Y-maze task: Relationship with hippocampal neurogenesis. Behav. Neurosci. 2007, 121, 324–334. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Y-maze for assessment of spatial working and reference memory in mice. Methods Mol. Biol. 2019, 1916, 105–111. [Google Scholar]
- Fayuk, D.; Yakel, J.L. Regulation of nicotinic acetylcholine receptor channel function by acetylcholinesterase inhibitors in rat hippocampal CA1 interneurons. Mol. Pharmacol. 2004, 66, 658–666. [Google Scholar] [CrossRef]
- Pallas, M.; Camins, A.; Smith, M.A.; Perry, G.; Lee, H.G.; Casadesus, G. From aging to Alzheimer ’s disease: Unveiling “the switch” with the senescence-accelerated mouse model (SAMP8). J. Alzheimer’s Dis. 2008, 15, 615–624. [Google Scholar] [CrossRef]
- Akbarian, M.; Hosseini, M.; Mizavi, F.; Amirahmadi, S.; Arab, F.A.; Rajabian, A. Punica granatum peel supplementation attenuates cognitive deficits and brain injury in rat by targeting the Nrf2-HO-1 pathway. Food Sci. Nutr. 2023, 11, 168–180. [Google Scholar] [CrossRef]
- Rinne, J.O.; Kaasinen, V.; Järvenpää, T.; Någren, K.; Roivanen, A.; Yu, M.; Oikonen, V.; Kurki, T. Brain acetylcholinesterase activity in mild cognitive impairment and early Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 2003, 74, 113–115. [Google Scholar] [CrossRef]
- Ahmed, T.; Zahid, S.; Mahboo, A.; Farhat, S.M. Cholinergic system and post-translational modifications: An insight on the role in Alzheimer’s disease. Curr. Neuropharmacol. 2017, 15, 480–494. [Google Scholar] [CrossRef]
- Parsons, C.G.; Danysz, W.; Dekundy, A.; Pulte, I. Memantine and cholinesterase inhibitors: Complementary mechanisms in the treatment of Alzheimer’s disease. Neurotox Res. 2013, 24, 358–369. [Google Scholar] [CrossRef]
- Kapoor, A.; Wang, B.J.; Hsu, W.M.; Chang, M.Y.; Liang, S.M.; Liao, Y.F. Retinoic acid-elicited RARα/RXRα signaling attenuates Aβ production by directly γ-secretase-mediated cleavage of amyloid precursor protein. ACS Chem. Neurosci. 2013, 4, 1093–1100. [Google Scholar] [CrossRef]
- Chen, C.H.; Zhou, W.; Liu, S.; Deng, Y.; Cai, F.; Tone, M.; Tone, Y.; Tong, Y.; Song, W. Increased NF-κB signaling up-regulates BACE1 expression and its therapeutic potential in Alzheimer’s disease. Int. J. Neuropsychopharmacol. 2012, 15, 77–90. [Google Scholar] [CrossRef]
- Ly, P.T.T.; Wu, Y.; Zou, H.; Wang, R.; Zhou, W.; Kinoshita, A.; Zhang, M.; Yang, Y.; Cai, F.; Woodgett, J.; et al. Inhibition of GSK3β-mediated BACE1 expression reduces Alzheimer-associated phenotypes. J. Clin. Investig. 2013, 123, 224–235. [Google Scholar] [CrossRef]
- Hernández-Rodríguez, M.; Arciniega-Martínez, I.M.; García-Marín, I.D.; Correa-Basurto, J.; Rosales-Hernández, M.C. Chronic Administration of Scopolamine Increased GSK3βP9, Beta Secretase, Amyloid Beta, and Oxidative Stress in the Hippocampus of Wistar Rats. Mol. Neurobiol. 2020, 57, 3979–3988. [Google Scholar] [CrossRef]
- Komulainen, P.; Pedersen, M.; Hänninen, T.; Bruunsgaard, H.; Lakka, T.A.; Kivipelto, M.; Hassinen, M.; Rauramaa, T.H.; Pedersen, B.K.; Rauramaa, R. BDNF is a novel marker of cognitive function in ageing women: The DR’s EXTRA Study. Neurobiol. Learn Mem. 2008, 90, 596–603. [Google Scholar] [CrossRef]
- Kuboyama, T.; Tohda, C.; Komatsu, K. Neuritic regeneration and synaptic reconstruction induced by withanolide A. Br. J. Pharmacol. 2005, 144, 961–971. [Google Scholar] [CrossRef]
- Konar, A.; Shah, N.; Singh, R.; Saxena, N.; Kaul, S.C.; Wadhwa, R.; Thaku, M.K. Protective Role of Ashwagandha Leaf Extract and Its Component Withanone on Scopolamine-Induced Changes in the Brain and Brain-Derived Cells. PLoS ONE 2011, 6, e27265. [Google Scholar] [CrossRef] [PubMed]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).