

Modeling the Effect of Selected Microorganisms’ Exposure to Molasses’s High-Osmolality Environment

 ,
,  ,
,  ,
,  ,
,  and
and 
Featured Application
Abstract
1. Introduction
- -
- Salmonella is the most essential pathogen originating from poultry, owing to salmonellosis, one of the most frequent diseases in public health [18];
- -
- -
- L. monocytogenes, a significant foodborne pathogen, is sometimes correlated with poultry products and occasionally leads to clinical disease in poultry [19].
2. Materials and Methods
2.1. Preparation of Sugar Beet Molasses Solutions
2.2. Contamination of Molasses Solutions
2.3. Incubation Conditions
2.4. Methods of Analysis of Selected Microorganisms
2.5. Response Surface Methodology
2.6. Correlation Analysis
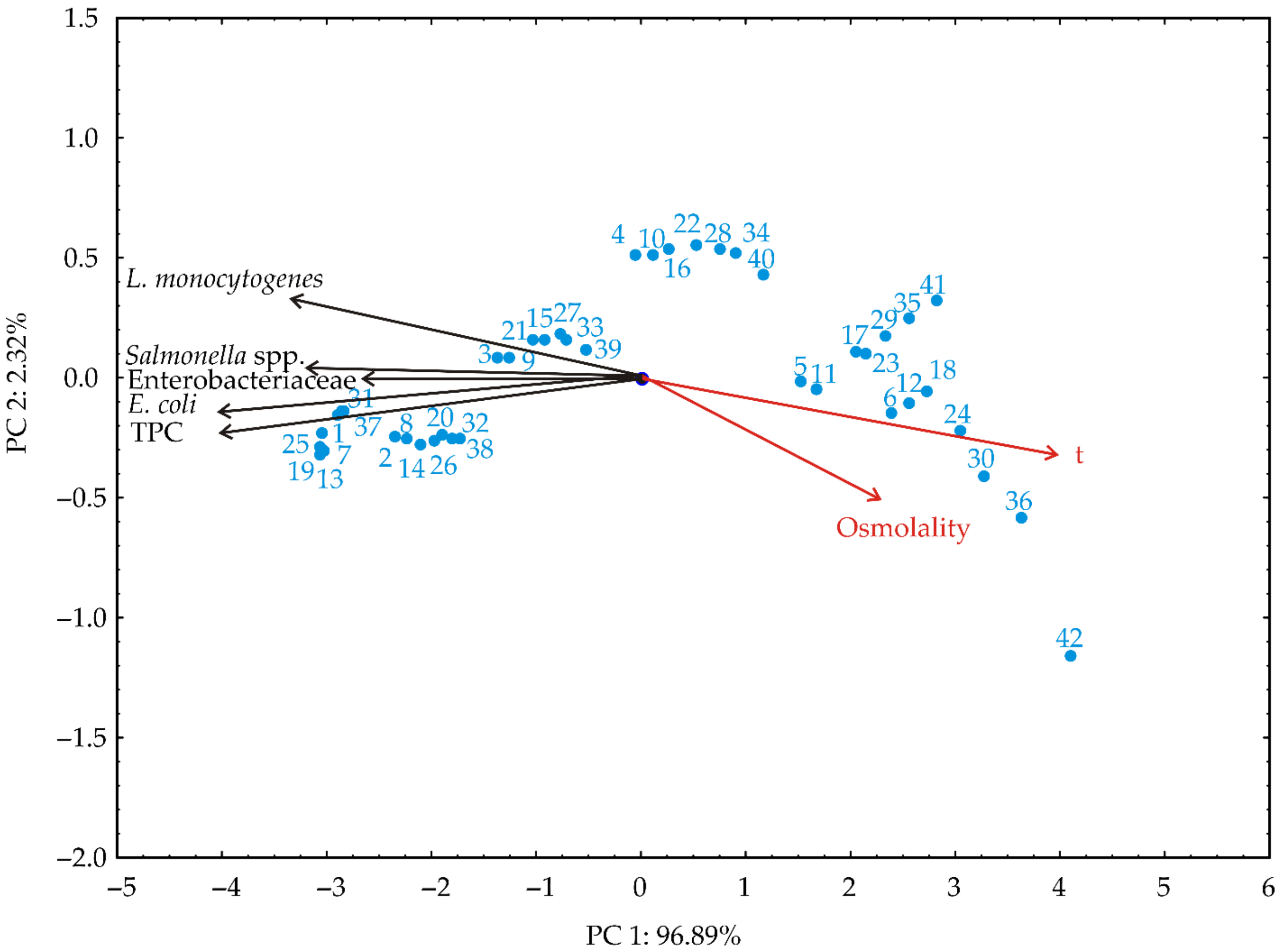
2.7. Principle Component Analysis
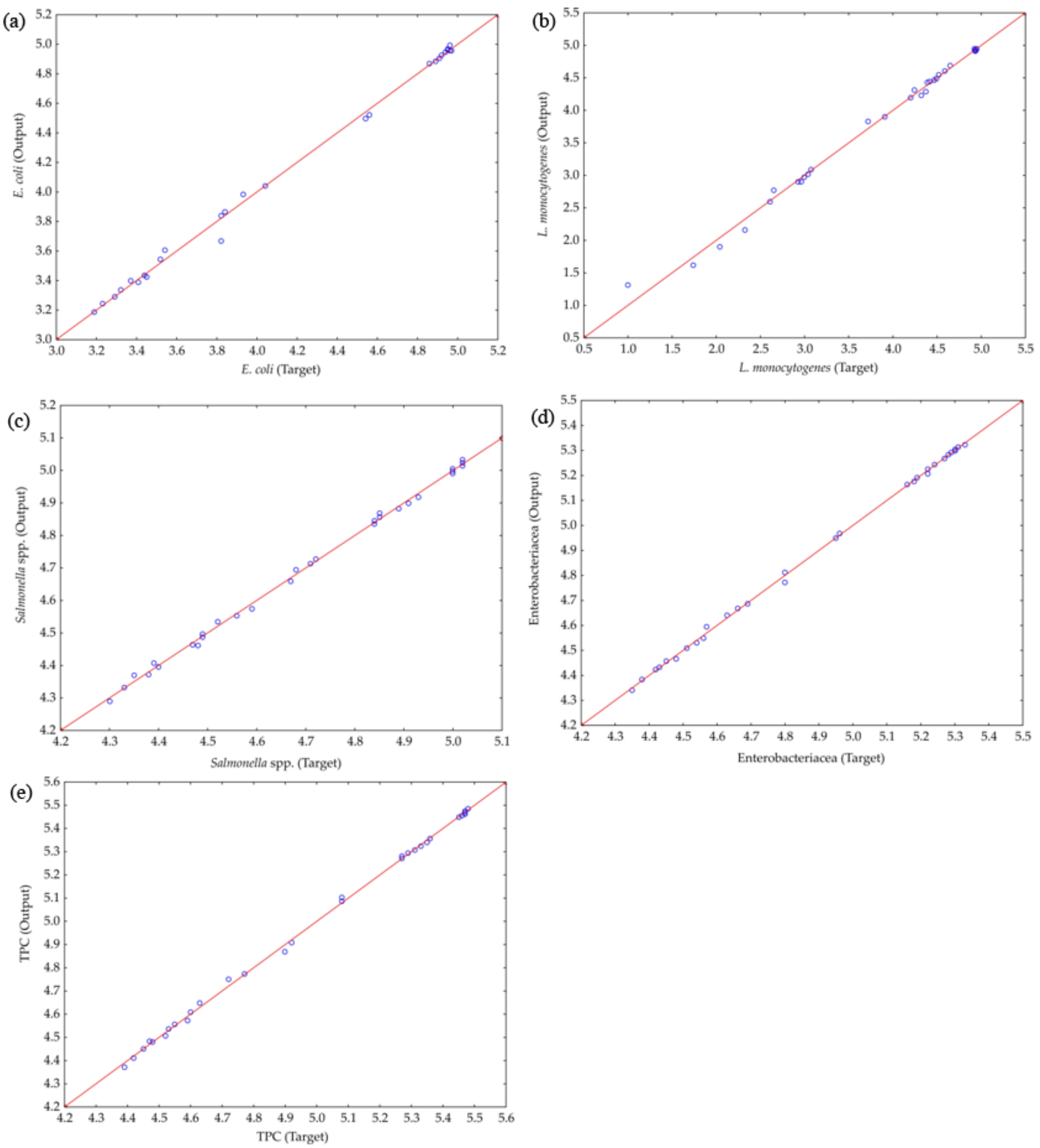
2.8. Artificial Neural Network Modeling
Error Analysis
3. Results and Discussion
3.1. RSM Modeling
3.2. Color Correlation Analysis
3.3. Principal Component Analysis
3.4. ANN Modeling
The Accuracy of the Models and the Residual Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arjeh, E.; Khodaei, S.M.; Barzegar, M.; Pirsa, S.; Karimi Sani, I.K.; Rahati, S.; Mohammadi, F. Phenolic compounds of sugar beet (Beta vulgaris L.): Separation method, chemical characterization, and biological properties. Food Sci. Nutr. 2022, 10, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, E.G.; Demidenko, A.V.; Zhila, N.O.; Shishatskaya, E.I.; Volova, T.G. Sugar Beet Molasses as a Potential C-Substrate for PHA Production by Cupriavidus necator. Bioengineering 2022, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Kaur, D.; Singh, M.; Zalpouri, R.; Singh, I. Osmotic dehydration of fruits using unconventional natural sweeteners and non-thermal-assisted technologies: A review. J. Food Process. Preserv. 2022, e16890. [Google Scholar] [CrossRef]
- Kudri, S.; Harshitha, T.; Hegde, P.; Tadkod, M.; Deasei, S.; Hemalatha, S. Effect of molasses, honey, and sugar on osmoticdehydration of muskmelon (Cucumis melo L.). Pharma Innov. 2022, 11, 162–166. Available online: https://www.thepharmajournal.com/archives/2022/vol11issue2/PartC/11-1-214-929.pdf (accessed on 12 January 2023).
- Bashir, N.; Sood, M.; Bandral, J. Food Preservation by Osmotic Dehydration—A Review. Chem. Sci. Rev. Lett. 2020, 9, 337–341. [Google Scholar] [CrossRef]
- Nićetin, M.; Pezo, L.; Lončar, B.; Filipović, V.; Šuput, D.; Zlatanović, S.; Dojčinović, B. Evaluation of water, sucrose and minerals effective diffusivities during osmotic treatment of pork in sugar beet molasses. Hem. Ind. 2015, 69, 241–251. [Google Scholar] [CrossRef]
- Filipović, I.; Ćurčić, B.; Filipović, V.; Nićetin, M.; Filipović, J.; Knežević, V. The effects of technological parameters on chicken meat osmotic dehydration process efficiency. J. Food Process. Preserv. 2016, 41, 13116. [Google Scholar] [CrossRef]
- Lončar, B.; Filipović, V.; Nićetin, M.; Knežević, V.; Pezo, L.; Filipčev, B.; Gubić, J. Influence of osmotic solutions on efficiency of osmotic dehydration treatment and sensorial properties of fish meat (Carassius gibelio). J. Hyg. Eng. Des. 2015, 13, 51–56. [Google Scholar]
- Filipović, V.; Filipović, J.; Lončar, B.; Knežević, V.; Nicetin, M.; Filipović, I. Synergetic dehydration method of osmotic treatment in molasses and successive lyophilization of peaches. J. Food Process. Preserv. 2022, 46, e16512. [Google Scholar] [CrossRef]
- Cvetković, B.; Pezo, L.; Šuput, D.; Lončar, B.; Šimurina, O.; Filipčev, B.; Jevtić-Mučibabić, R. Shelf-life study of osmodehydrated white cabbage packaged in modified atmosphere: Mathematical approach. J. Appl. Bot. Food Qual. 2021, 94, 47–52. [Google Scholar] [CrossRef]
- Nićetin, M.; Pezo, L.; Pergal, M.; Lončar, B.; Filipović, V.; Knežević, V.; Demir, H.; Filipović, J.; Manojlović, D. Celery root phenols content, antioxidant capacities and their correlations after osmotic dehydration in molasses. Foods 2022, 11, 1945. [Google Scholar] [CrossRef] [PubMed]
- Sjölin, M.; Thuvander, J.; Wallberg, O.; Lipnizki, F. Purification of Sucrose in Sugar Beet Molasses by Utilizing Ceramic Nanofiltration and Ultrafiltration Membranes. Membranes 2020, 10, 5. [Google Scholar] [CrossRef]
- Chen, M.; Zhao, Z.; Meng, H.; Yu, S. The antibiotic activity and mechanisms of sugar beet (Beta vulgaris) molasses polyphenols against selected food-borne pathogens. LWT–Food Sci. Technol. 2017, 82, 354–360. [Google Scholar] [CrossRef]
- Kulkarni, V.S.; Shaw, C. Miscellaneous Physical, Chemical, and Microbiological Test Methods. In Essential Chemistry for Formulators of Semisolid and Liquid Dosages; Kulkarni, V.S., Shaw, C., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 193–221. [Google Scholar] [CrossRef]
- Šklubalova, Z.; Zatloukal, Z. Conversion Between Osmolality and Osmolarity of Infusion Solutions. Sci. Pharm. 2009, 77, 817–826. [Google Scholar] [CrossRef]
- Alhuthali, S.; Kotidis, P.; Kontoravdi, C. Osmolality Effects on CHO Cell Growth, Cell Volume, Antibody Productivity and Glycosylation. Int. J. Mol. Sci. 2021, 22, 3290. [Google Scholar] [CrossRef]
- Commission Regulation (EC). No 2073/2005 of 15 November 2005 on Microbiological Criteria for Foodstuffs. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:02005R2073-20200308&from=EN (accessed on 12 January 2023).
- Ramatla, T.; Mileng, K.; Ndou, R.; Mphuti, N.; Syakalima, M.; Lekota, K.E.; Thekisoe, O.M.M. Molecular Detection of Integrons, Colistin and β-lactamase Resistant Genes in Salmonella enterica Serovars Enteritidis and Typhimurium Isolated from Chickens and Rats Inhabiting Poultry Farms. Microorganisms 2022, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Hargis, B.M.; Caldwell, D.J.; Byrd, J.A. Microbiological pathogens: Live poultry considerations. In Poultry Meat Processing, 2nd ed.; Sams, A., Ed.; CRC PressTaylor & Francis Group: Boca Raton, FL, USA, 2010; pp. 125–141. [Google Scholar]
- Park, J.Y.; Lim, M.-C.; Park, K.; Ok, G.; Chang, H.-J.; Lee, N.; Park, T.J.; Choi, S.-W. Detection of E. coli O157:H7 in Food Using Automated Immunomagnetic Separation Combined with Real-Time PCR. Processes 2020, 8, 908. [Google Scholar] [CrossRef]
- Guillén, S.; Nadal, L.; Álvarez, I.; Mañas, P.; Cebrián, G. Impact of the Resistance Responses to Stress Conditions Encountered in Food and Food Processing Environments on the Virulence and Growth Fitness of Non-Typhoidal Salmonellae. Foods 2021, 10, 617. [Google Scholar] [CrossRef]
- Dessaux, C.; Guerreiro, D.N.; Pucciarelli, M.G.; Conor, P.; O’Byrne, C.P.; García-del Portillo, G. Impact of osmotic stress on the phosphorylation and subcellular location of Listeria monocytogenes stressosome proteins. Sci Rep. 2020, 10, 20837. [Google Scholar] [CrossRef]
- Funes, E.; Allouche, Y.; Beltrán, G.; Jiménez, A. A review: Artificial neural networks as tool for control food industry process. J. Sens. Technol. 2015, 5, 28. [Google Scholar] [CrossRef]
- Nayak, J.; Vakula, K.; Dinesh, P.; Naik, B.; Pelusi, D. Intelligent food processing: Journey from artificial neural network to deep learning. Comput. Sci. Rev. 2020, 38, 100297. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, Q.; Wang, J. Big data analysis using neural networks. Adv. Eng. Sci. 2017, 49, 9–18. [Google Scholar]
- Guiné, R.P.F. The Use of Artificial Neural Networks (ANN) in Food Process Engineering. IJFE 2019, 5, 15–21. [Google Scholar] [CrossRef]
- Gorbachev, V.; Nikitina, M.; Velina, D.; Mutallibzoda, S.; Nosov, V.; Korneva, G.; Terekhova, A.; Artemova, E.; Khashir, B.; Sokolov, I.; et al. Artificial Neural Networks for Predicting Food Antiradical Potential. Appl. Sci. 2022, 12, 6290. [Google Scholar] [CrossRef]
- Ahmed, M.; AlQadhi, S.; Mallick, J.; Kahla, N.B.; Le, H.A.; Singh, C.K.; Hang, H.T. Artificial Neural Networks for Sustainable Development of the Construction Industry. Sustainability 2022, 14, 14738. [Google Scholar] [CrossRef]
- ISO 16649-2:2001; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Beta-Glucuronidase-Positive Escherichia. coli—Part 2: Colony-Count Technique at 44 Degrees C Using 5-bromo-4-chloro-3-indolyl beta-D-glucuronide. International Organization for Standardization: Geneva, Switzerland, 2001.
- ISO 6579-1:2017; Microbiology of the Food Chain-Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella-Part 1: Horizontal Method for the Detection of Salmonella spp. International Organization for Standardization: Geneva, Switzerland, 2017.
- ISO 11290-2:2017; Microbiology of the food chain—Horizontal method for the detection and enumeration of Listeria monocytogenes and of Listeria spp.—Part 2: Enumeration Method. International Organization for Standardization: Geneva, Switzerland, 2017.
- ISO 21528-2:2017; Microbiology of the Food Chain—Horizontal Method for the DETECTION and enumeration of Enterobacteriaceae—Part 2: Colony-Count Technique. International Organization for Standardization: Geneva, Switzerland, 2017.
- ISO 4833-1:2014; Microbiology of the Food Chain—Horizontal Method for the Enumeration of Microorganisms Colony Count at 30 Degrees C by the Pour Plate Technique. International Organization for Standardization: Geneva, Switzerland, 2014.
- ISO 11133:2014; Microbiology of Food, Animal Feed and Water—Preparation, Production, Storage and Performance Testing of Culture Media. International Organization for Standardization: Geneva, Switzerland, 2014.
- Filipović, I.; Markov, S.; Filipović, V.; Filipović, J.; Vidaković, A.; Novković, N.; Rafajlovska, V. Modeling of factors influencing the effect of osmotic solution on reduction of selected microorganisms. J. Appl. Microbiol. 2018, 125, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Rajković, D.; Marjanović Jeromela, A.; Pezo, L.; Lončar, B.; Zanetti, F.; Monti, A.; Kondić Špika, A. Yield and Quality Prediction of Winter Rapeseed—Artificial Neural Network and Random Forest Models. Agronomy 2021, 12, 58. [Google Scholar] [CrossRef]
- Vojnov, B.; Jaćimović, G.; Šeremešić, S.; Pezo, L.; Lončar, B.; Krstić, Đ.; Vujić, S.; Ćupina, B. The Effects of Winter Cover Crops on Maize Yield and Crop Performance in Semiarid Conditions—Artificial Neural Network Approach. Agronomy 2022, 12, 2670. [Google Scholar] [CrossRef]
- Voća, M.; Pezo, L.; Jukić, Ž.; Lončar, B.; Šuput, D.; Krička, T. Estimation of the storage properties of rapeseeds using an artificial neural network. Ind. Crops Prod. 2022, 187, 115358. [Google Scholar] [CrossRef]
- Pezo, L.; Lončar, B.; Šovljanski, O.; Tomić, A.; Travičić, V.; Pezo, M.; Aćimović, M. Agricultural Parameters and Essential Oil Content Composition Prediction of Aniseed. Based on Growing Year. Locality and Fertilization Type—An Artificial Neural Network Approach. Life 2022, 12, 1722. [Google Scholar] [CrossRef]
- Brandić, I.; Pezo, L.; Bilandžija, N.; Peter, A.; Šurić, J.; Voća, N. Artificial neural network as a tool for estimation of the higher heating value of miscanthus based on ultimate analysis. Mathematics 2022, 10, 3732. [Google Scholar] [CrossRef]
- Ruškić, N.; Mirović, V.; Marić, M.; Pezo, L.; Lončar, B.; Nićetin, M.; Ćurčić, Lj. Model for Determining Noise Level Depending on Traffic Volume at Intersections. Sustainability 2022, 14, 12443. [Google Scholar] [CrossRef]
- Zi, X.; Liu, Y.; Chen, T.; Li, M.; Zhou, H.; Tang, J. Effects of Sucrose, Glucose and Molasses on Fermentation Quality and Bacterial Community of Stylo Silage. Fermentation 2022, 8, 191. [Google Scholar] [CrossRef]
- Shafiqa-Atikah, M.K.; Nor-Khaizura, M.A.R.; Mahyudin, N.A.; Abas, F.; Nur-Syifa, J.; Ummul-Izzatul, Y. Evaluation of phenolic constituent, antioxidant and antibacterial activities of sugarcane molasses towards foodborne pathogens. Food Res. 2020, 4, 40–47. [Google Scholar]
- Wood, J.M. Bacterial responses to osmotic challenges. J. Gen. Physiol. 2015, 145, 381–388. [Google Scholar] [CrossRef]
- Chicco, D.; Warrens, M.J.; Jurman, G. The coefficient of determination R-squared is more informative than SMAPE, MAE, MAPE, MSE and RMSE in regression analysis evaluation. Peer J. Comput. Sci. 2021, 7, e623. [Google Scholar] [CrossRef] [PubMed]
- Filipović, V.; Lončar, B.; Filipović, J.; Nićetin, M.; Knežević, V.; Šeregelj, V.; Košutić, M.; Bodroža Solarov, M. Addition of Combinedly Dehydrated Peach to the Cookies—Technological Quality Testing and Optimization. Foods 2022, 11, 1258. [Google Scholar] [CrossRef]
- Pezo, L.L.; Ćurčić, B.L.; Filipović, V.S.; Nićetin, M.R.; Koprivica, G.B.; Mišljenović, N.M.; Lević, L.B. Artificial neural network model of pork meat cubes osmotic dehydratation. Hem. Ind. 2013, 67, 465–475. [Google Scholar] [CrossRef]
- Ćurčić, B.L.; Pezo, L.L.; Filipović, V.S.; Nićetin, M.R.; Knežević, V. Osmotic treatment of fish in two different solutions-artificial neural network model. J. Food Process. Preserv. 2015, 39, 671–680. [Google Scholar] [CrossRef]
- Lončar, B.; Pezo, L.; Filipović, V.; Nićetin, M.; Filipović, J.; Pezo, M.; Šuput, D.; Aćimović, M. Physico-Chemical, Textural and Sensory Evaluation of Spelt Muffins Supplemented with Apple Powder Enriched with Sugar Beet Molasses. Foods 2022, 11, 1750. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run No. | Time (h) | Osmolality (mmol/kg) | E. coli log10(cfu/g) | L. monocytogenes log10(cfu/g) | Salmonella spp. log10(cfu/g) | Entero- bacteriaceae log10(cfu/g) | TPC log10(cfu/g) |
|---|---|---|---|---|---|---|---|
| 1 | 0 | 5500 | 4.98 ± 0.01 20 | 4.95 ± 0.01 20 | 5.06 ± 0.03 20 | 5.35 ± 0.01 23 | 5.50 ± 0.01 22 |
| 2 | 0.5 | 5500 | 4.96 ± 0.00 20 | 4.65 ± 0.01 19 | 4.93 ± 0.01 18 | 5.27 ± 0.02 19–21 | 5.36 ± 0.00 21 |
| 3 | 1 | 5500 | 4.69 ± 0.01 17 | 4.52 ± 0.02 17,18 | 4.80 ± 0.01 15,16 | 5.06 ± 0.08 15,16 | 5.19 ± 0.02 17 |
| 4 | 2 | 5500 | 4.11 ± 0.05 13 | 4.31 ± 0.01 13,14 | 4.71 ± 0.01 11–13 | 4.82 ± 0.00 13 | 4.93 ± 0.00 13 |
| 5 | 3 | 5500 | 3.88 ± 0.01 9–11 | 3.16 ± 0.02 8 | 4.54 ± 0.01 6,7 | 4.64 ± 0.01 9 | 4.64 ± 0.01 9 |
| 6 | 5 | 5500 | 3.45 ± 0.01 7 | 2.65 ± 0.07 5 | 4.49 ± 0.02 5,6 | 4.54 ± 0.01 5–7 | 4.55 ± 0.01 6,7 |
| 7 | 0 | 5750 | 4.97 ± 0.01 20 | 4.95 ± 0.01 20 | 5.02 ± 0.03 19,20 | 5.33 ± 0.01 22,23 | 5.48 ± 0.01 22 |
| 8 | 0.5 | 5750 | 4.95 ± 0.00 19,20 | 4.59 ± 0.01 18,19 | 4.91 ± 0.01 18 | 5.24 ± 0.02 18–20 | 5.35 ± 0.01 20,21 |
| 9 | 1 | 5750 | 4.64 ± 0.01 16,17 | 4.48 ± 0.02 16,17 | 4.79 ± 0.01 14,15 | 5.06 ± 0.03 16 | 5.16 ± 0.02 16,17 |
| 10 | 2 | 5750 | 4.04 ± 0.06 12 | 4.24 ± 0.02 12,13 | 4.68 ± 0.01 10,11 | 4.80 ± 0.01 12,13 | 4.92 ± 0.00 13 |
| 11 | 3 | 5750 | 3.82 ± 0.00 9 | 3.07 ± 0.10 11 | 4.52 ± 0.03 6,7 | 4.63 ± 0.01 8,9 | 4.63 ± 0.02 8,9 |
| 12 | 5 | 5750 | 3.37 ± 0.04 4–6 | 2.61 ± 0.01 5 | 4.47 ± 0.01 4,5 | 4.51 ± 0.01 5,6 | 4.53 ± 0.02 6 |
| 13 | 0 | 6000 | 4.97 ± 0.01 20 | 4.94 ± 0.00 20 | 5.02 ± 0.03 19,20 | 5.31 ± 0.01 21–23 | 5.47 ± 0.01 22 |
| 14 | 0.5 | 6000 | 4.94 ± 0.00 19,20 | 4.52 ± 0.01 17,18 | 4.89 ± 0.00 17,18 | 5.22 ± 0.02 17–19 | 5.33 ± 0.01 19–21 |
| 15 | 1 | 6000 | 4.61 ± 0.01 15,16 | 4.45 ± 0.01 15–17 | 4.76 ± 0.01 13–15 | 5.00 ± 0.00 14,15 | 5.13 ± 0.02 116 |
| 16 | 2 | 6000 | 3.93 ± 0.04 11 | 4.20 ± 0.04 11,12 | 4.67 ± 0.02 10,11 | 4.80 ± 0.00 12,13 | 4.90 ± 0.01 13 |
| 17 | 3 | 6000 | 3.54 ± 0.01 8 | 3.04 ± 0.06 7 | 4.49 ± 0.02 5,6 | 4.57 ± 0.01 7,8 | 4.60 ± 0.02 8,9 |
| 18 | 5 | 6000 | 3.35 ± 0.04 3–5 | 2.59 ± 0.02 5 | 4.44 ± 0.01 3,4 | 4.48 ± 0.01 4,5 | 4.50 ± 0.02 4–6 |
| 19 | 0 | 6250 | 4.97 ± 0.01 20 | 4.93 ± 0.00 20 | 5.02 ± 0.03 19,20 | 5.30 ± 0.00 20–23 | 5.47 ± 0.01 22 |
| 20 | 0.5 | 6250 | 4.92 ± 0.00 18–20 | 4.49 ± 0.02 16–18 | 4.85 ± 0.00 16,17 | 5.22 ± 0.02 17–19 | 5.31 ± 0.01 18–20 |
| 21 | 1 | 6250 | 4.59 ± 0.01 14–16 | 4.40 ± 0.02 14–16 | 4.74 ± 0.01 12–14 | 4.98 ± 0.00 14 | 5.11 ± 0.00 15,16 |
| 22 | 2 | 6250 | 3.90 ± 0.02 10,11 | 4.10 ± 0.02 11 | 4.64 ± 0.01 9,10 | 4.75 ± 0.01 11,12 | 4.84 ± 0.01 12 |
| 23 | 3 | 6250 | 3.52 ± 0.01 8 | 3.00 ± 0.06 6,7 | 4.48 ± 0.01 4–6 | 4.56 ± 0.02 6,7 | 4.59 ± 0.01 7,8 |
| 24 | 5 | 6250 | 3.32 ± 0.03 3,4 | 2.32 ± 0.03 4 | 4.39 ± 0.01 2,3 | 4.45 ± 0.02 3,4 | 4.47 ± 0.01 2–4 |
| 25 | 0 | 6500 | 4.96 ± 0.01 20 | 4.93 ± 0.00 20 | 5.00 ± 0.00 19 | 5.30 ± 0.00 20–23 | 5.47 ± 0.01 22 |
| 26 | 0.5 | 6500 | 4.91 ± 0.01 18–20 | 4.47 ± 0.01 15–17 | 4.85 ± 0.01 16,17 | 5.19 ± 0.02 17,18 | 5.29 ± 0.02 18,19 |
| 27 | 1 | 6500 | 4.56 ± 0.03 14,15 | 4.37 ± 0.01 13–15 | 4.72 ± 0.01 11–13 | 4.96 ± 0.01 14 | 5.08 ± 0.00 15 |
| 28 | 2 | 6500 | 3.87 ± 0.00 9–11 | 3.99 ± 0.02 10 | 4.61 ± 0.01 8,9 | 4.71 ± 0.01 10,11 | 4.80 ± 0.00 11,12 |
| 29 | 3 | 6500 | 3.47 ± 0.01 7,8 | 2.98 ± 0.03 6,7 | 4.45 ± 0.02 4,5 | 4.52 ± 0.00 5–7 | 4.55 ± 0.01 6,7 |
| 30 | 5 | 6500 | 3.29 ± 0.02 2,3 | 2.04 ± 0.06 3 | 4.38 ± 0.03 2,3 | 4.42 ± 0.01 2,3 | 4.45 ± 0.02 2,3 |
| 31 | 0 | 6750 | 4.95 ± 0.00 19,20 | 4.92 ± 0.00 20 | 5.00 ± 0.00 19 | 5.29 ± 0.02 20–22 | 5.46 ± 0.02 22 |
| 32 | 0.5 | 6750 | 4.89 ± 0.01 18,19 | 4.42 ± 0.01 15–17 | 4.84 ± 0.01 16,17 | 5.18 ± 0.00 17 | 5.27 ± 0.02 18 |
| 33 | 1 | 6750 | 4.54 ± 0.03 14 | 4.32 ± 0.03 13,14 | 4.71 ± 0.01 11–13 | 4.95 ± 0.01 14 | 5.08 ± 0.00 15 |
| 34 | 2 | 6750 | 3.84 ± 0.03 9,10 | 3.91 ± 0.01 10 | 4.59 ± 0.01 7,8 | 4.69 ± 0.01 9,10 | 4.77 ± 0.01 11 |
| 35 | 3 | 6750 | 3.44 ± 0.01 6,7 | 2.96 ± 0.05 6,7 | 4.40 ± 0.02 2,3 | 4.48 ± 0.01 4,5 | 4.52 ± 0.03 5,6 |
| 36 | 5 | 6750 | 3.23 ± 0.04 1,2 | 1.74 ± 0.06 2 | 4.33 ± 0.04 1,2 | 4.38 ± 0.03 1,2 | 4.42 ± 0.01 1,2 |
| 37 | 0 | 7000 | 4.95 ± 0.00 19,20 | 4.92 ± 0.01 20 | 5.00 ± 0.01 19 | 5.28 ± 0.00 20–22 | 5.45 ± 0.01 22 |
| 38 | 0.5 | 7000 | 4.86 ± 0.01 18 | 4.39 ± 0.01 14–16 | 4.84 ± 0.01 16,17 | 5.16 ± 0.02 17 | 5.27 ± 0.02 18 |
| 39 | 1 | 7000 | 4.52 ± 0.03 14 | 4.20 ± 0.08 11,12 | 4.69 ± 0.01 11,12 | 4.94 ± 0.00 14 | 5.02 ± 0.03 14 |
| 40 | 2 | 7000 | 3.82 ± 0.00 9 | 3.72 ± 0.03 9 | 4.56 ± 0.01 7,8 | 4.66 ± 0.01 9,10 | 4.72 ± 0.01 10 |
| 41 | 3 | 7000 | 3.41 ± 0.01 5–7 | 2.93 ± 0.04 6 | 4.35 ± 0.01 1,2 | 4.43 ± 0.02 2–4 | 4.48 ± 0.02 3–5 |
| 42 | 5 | 7000 | 3.19 ± 0.02 1 | 1.00 ± 0.00 1 | 4.30 ± 0.03 1 | 4.35 ± 0.01 1 | 4.39 ± 0.01 1 |
| Independent Variables | Term | df + | Sum of Squares | ||||
|---|---|---|---|---|---|---|---|
| E. coli | L. monocytogenes | Salmonella spp. | Enterobacteriaceae | TPC | |||
| Exposure Time | Linear | 1 | 15.88 * | 38.03 * | 1.75 * | 3.89 * | 5.10 * |
| Quadratic | 1 | 1.07 * | 0.01 | 0.19 * | 0.42 * | 0.53 * | |
| Osmolality | Linear | 1 | 0.26 * | 1.67 * | 0.09 * | 0.11 * | 0.10 * |
| Quadratic | 1 | 0.01 | 0.06 | 0.00 | 0.00 | 0.00 | |
| Cross Product | Time × osmolality | 1 | 0.04 | 0.86 * | 0.01 * | 0.01 * | 0.01 * |
| Error | Residual variance | 36 | 0.50 | 1.56 | 0.024 | 0.03 | 0.03 |
| Total sum of squares | 41 | 17.77 | 41.60 | 2.07 | 4.46 | 5.79 | |
| R2 | 0.97 | 0.96 | 0.99 | 0.99 | 0.99 | ||
| E. coli | L. monocytogenes | Salmonella spp. | Enterobacteriaceae | TPC | |
|---|---|---|---|---|---|
| β0 | 8.103405 * | −1.90308 | 5.372118 * | 5.768105 * | 5.468519 * |
| β1 | −0.450115 * | 0.52251 * | −0.143098 * | −0.276613 * | −0.351648 * |
| β11 | 0.063844 * | −0.00506 | 0.027177 * | 0.040088 * | 0.045015 * |
| β2 | −0.000890 | 0.00219 | −0.000066 | −0.000072 | 0.000079 |
| β22 | 0.000000 | −0.00000 | 0.000000 | 0.000000 | −0.000000 |
| β12 | −0.000037 | −0.00017 * | −0.000018 * | −0.000017 * | −0.000013 * |
| Network Name | Performance * | Error ** | Training Algorithm | Error Function | Hidden Activation | Output Activation | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Train. | Test. | Valid. | Train. | Test. | Valid. | |||||
| MLP 2-7-5 | 0.999 | 0.992 | 0.999 | 0.004 | 0.007 | 0.004 | BFGS 222 | SOS | Logistic | Identity |
| Cycle | ANN | ||||
|---|---|---|---|---|---|
| E. coli | L. monocytogenes | Salmonella spp. | Enterobacteriaceae | TPC | |
| Train | 0.9998 | 0.9969 | 0.9990 | 0.9996 | 0.9995 |
| Test | 0.9889 | 0.9971 | 0.9868 | 0.9936 | 0.9983 |
| Validation | 0.9994 | 0.9992 | 0.9985 | 0.9994 | 0.9998 |
| Parameter | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| Osmolality | −7.598 | −10.376 | 4.913 | −6.278 | −8.156 | −5.626 | −9.446 |
| Time | 1.027 | 0.663 | −1.211 | −0.228 | −0.219 | 0.019 | −0.431 |
| Bias | 4.354 | 6.193 | −2.841 | −1.827 | 2.056 | −1.123 | 3.374 |
| Outputs | 1 | 2 | 3 | 4 | 5 | 6 | 7 | Bias |
|---|---|---|---|---|---|---|---|---|
| E. coli | −1.975 | 2.184 | 0.238 | −3.249 | 3.170 | −0.966 | −1.380 | −0.038 |
| L. monocytogenes | 3.812 | −0.721 | 3.187 | −2.314 | −0.781 | 3.672 | 0.430 | −2.516 |
| Salmonella spp. | −1.050 | 2.077 | 1.293 | 0.412 | 0.956 | 1.309 | −0.427 | −0.883 |
| Enterobacteriaceae | −2.312 | 2.685 | 0.499 | −1.411 | 3.048 | −0.823 | −1.563 | −0.213 |
| TPC | −0.844 | 1.456 | 0.758 | −0.376 | 0.957 | 0.904 | −0.157 | −0.513 |
| Parameter | χ2 | RMSE | MBE | MPE | SSE | AARD | r2 |
|---|---|---|---|---|---|---|---|
| E. coli | 0.001 | 0.037 | −2.5 × 10−5 | 0.574 | 0.041 | 0.924 | 0.997 |
| L. monocytogenes | 0.007 | 0.085 | 0.000 | 2.553 | 0.217 | 1.859 | 0.994 |
| Salmonella spp. | 0.000 | 0.010 | −3.7 × 10−5 | 0.197 | 0.003 | 0.568 | 0.998 |
| Enterobacteriaceae | 9.14 × 10−5 | 0.009 | −0.000 | 0.145 | 0.003 | 0.351 | 0.999 |
| TPC | 0.000 | 0.012 | −1.5 × 10−5 | 0.196 | 0.005 | 0.541 | 0.999 |
| Parameter | Skew | Kurt | Mean | StDev | Var |
|---|---|---|---|---|---|
| E. coli | 2.294 | 9.468 | −2.5 × 10−5 | 0.037 | 0.001 |
| L. monocytogenes | −1.509 | 5.684 | 0.000 | 0.086 | 0.007 |
| Salmonella spp. | −0.278 | −0.906 | −3.7 × 10−5 | 0.010 | 0.000 |
| Enterobacteriaceae | 0.356 | 2.228 | 0.000 | 0.009 | 9.14 × 10−5 |
| TPC | 0.036 | 0.861 | −1.5 × 10−5 | 0.012 | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filipović, V.; Lončar, B.; Knežević, V.; Nićetin, M.; Filipović, J.; Petković, M. Modeling the Effect of Selected Microorganisms’ Exposure to Molasses’s High-Osmolality Environment. Appl. Sci. 2023, 13, 1207. https://doi.org/10.3390/app13021207
Filipović V, Lončar B, Knežević V, Nićetin M, Filipović J, Petković M. Modeling the Effect of Selected Microorganisms’ Exposure to Molasses’s High-Osmolality Environment. Applied Sciences. 2023; 13(2):1207. https://doi.org/10.3390/app13021207
Chicago/Turabian StyleFilipović, Vladimir, Biljana Lončar, Violeta Knežević, Milica Nićetin, Jelena Filipović, and Marko Petković. 2023. "Modeling the Effect of Selected Microorganisms’ Exposure to Molasses’s High-Osmolality Environment" Applied Sciences 13, no. 2: 1207. https://doi.org/10.3390/app13021207
APA StyleFilipović, V., Lončar, B., Knežević, V., Nićetin, M., Filipović, J., & Petković, M. (2023). Modeling the Effect of Selected Microorganisms’ Exposure to Molasses’s High-Osmolality Environment. Applied Sciences, 13(2), 1207. https://doi.org/10.3390/app13021207