
Anaerobic Fermentation of Silage from the Above-Ground Biomass of Jerusalem Artichoke (Helianthus tuberosus L.) and Maize (Zea mayse L.) as a New and Promising Input Raw Material for Biogas Production

 , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
2. Materials and Methods
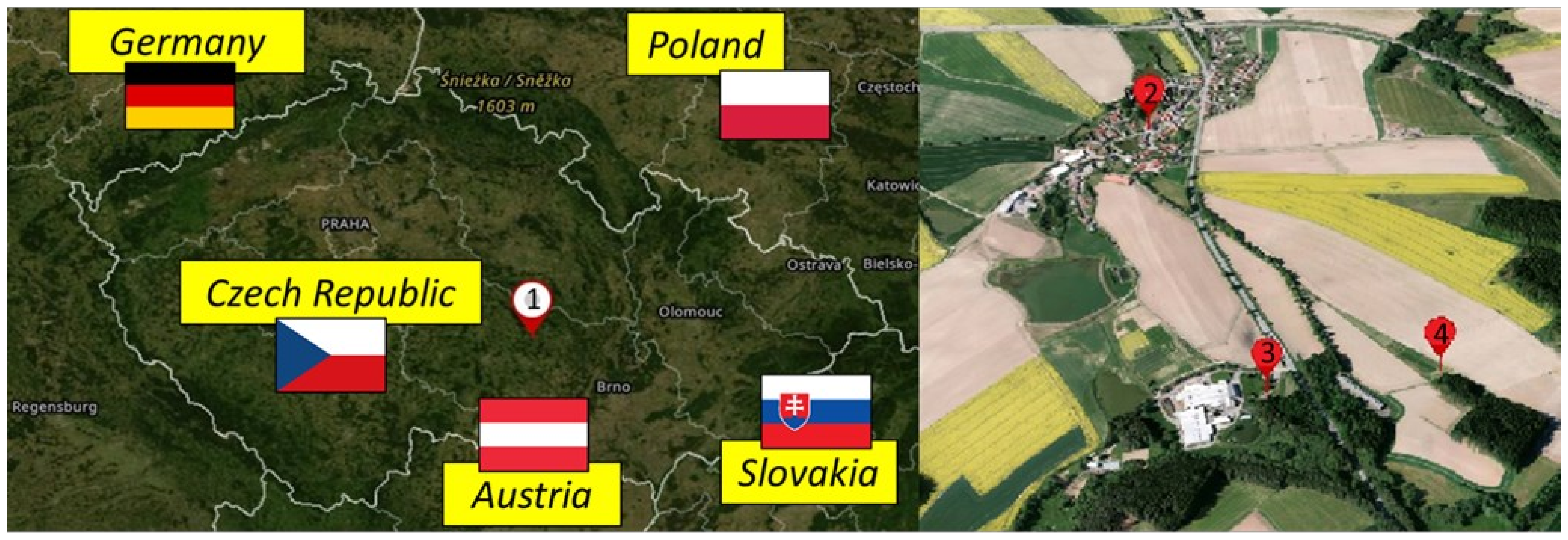
2.1. Localization of Biomass Source
2.2. Production of Model Silage
2.3. Silage Characteristics
2.4. Fermentation Tests
2.5. DNA Isolation and 16S rRNA Sequencing
Sequence Analysis
2.6. Statistical Analysis–Data Treatment
3. Results and Discussion
3.1. Silage Characteristics
3.2. Methane Yield
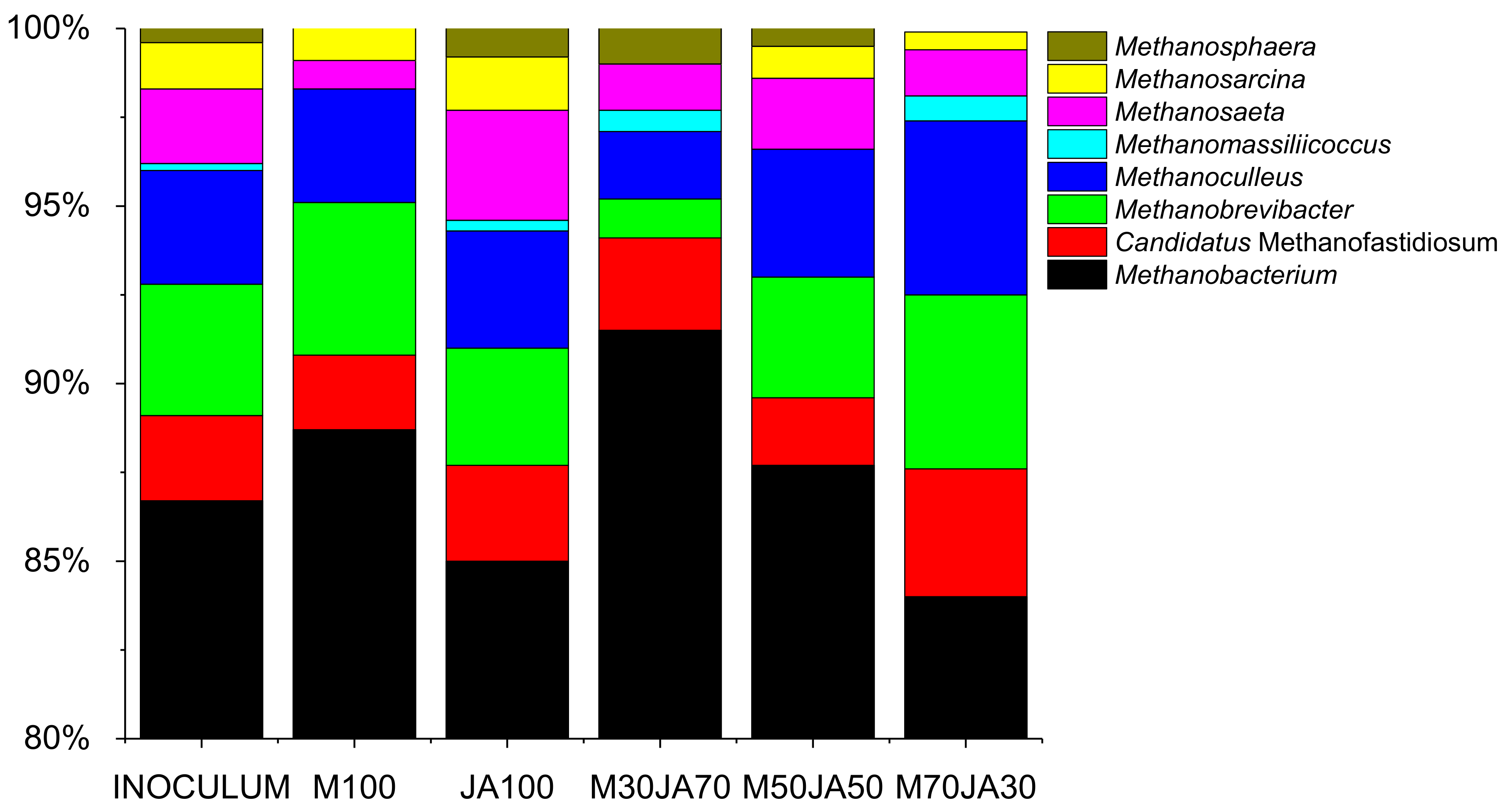
3.3. Composition of Microorganisms
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Lajdova, Z.; Lajda, J.; Kapusta, J.; Bielik, P. Consequences of maize cultivation intended for biogas production. Agric. Econ. Czech 2016, 62, 543–549. [Google Scholar]
- Herrmann, A. Biogas Production from Maize: Current State, Challenges and Prospects. 2. Agronomic and Environmental Aspects. Bioenergy Res. 2013, 6, 372–387. [Google Scholar] [CrossRef]
- Menšík, L.; Kincl, D.; Nerušil, P.; Srbek, J.; Hlisnikovský, L.; Smutný, V. Water erosion reduction using different soil tillage approaches for maize (Zea mays L.) in the Czech Republic. Land 2020, 9, 358. [Google Scholar] [CrossRef]
- Uzoh, I.M.; Igwe, C.A.; Okebalama, C.B.; Babalola, O.O. Legume-maize rotation effect on maize productivity and soil fertility parameters under selected agronomic practices in a sandy loam soil. Sci. Rep. 2019, 9, 8539. [Google Scholar] [CrossRef] [PubMed]
- Mezera, J.; Lukas, V.; Horniaček, I.; Smutný, V.; Elbl, J. Comparison of proximal and remote sensing for the diagnosis of crop status in site-specific crop management. Sensors 2021, 22, 19. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Moreno, F.; Lukas, V.; Neudert, L.; Dryšlová, T. Spatial interpretation of plant parameters in winter wheat. Precis. Agric. 2014, 15, 447–465. [Google Scholar] [CrossRef]
- García-Gen, S.; Rodríguez, J.; Lema, J.M. Optimisation of substrate blends in anaerobic co-digestion using adaptive linear programming. Bioresour. Technol. 2014, 173, 159–167. [Google Scholar] [CrossRef]
- Antoniou, N.; Monlau, F.; Sambusiti, C.; Ficara, E.; Barakat, A.; Zabaniotou, A. Contribution to Circular Economy options of mixed agricultural wastes management: Coupling anaerobic digestion with gasification for enhanced energy and material recovery. J. Clean. Prod. 2019, 209, 505–514. [Google Scholar] [CrossRef]
- Stinner, P.W. The use of legumes as a biogas substrate—Potentials for saving energy and reducing greenhouse gas emissions through symbiotic nitrogen fixation. Energy Sustain. Soc. 2015, 5, 4. [Google Scholar] [CrossRef]
- Kintl, A.; Vítěz, T.; Elbl, J.; Vítězová, M.; Dokulilová, T.; Nedělník, J.; Skládanka, J.; Brtnický, M. Mixed Culture of Corn and White Lupine as an Alternative to Silage Made from Corn Monoculture Intended for Biogas Production. Bioenergy Res. 2019, 12, 694–702. [Google Scholar] [CrossRef]
- Nabel, M.; Schrey, S.D.; Temperton, V.M.; Harrison, L.; Jablonowski, N.D. Legume Intercropping With the Bioenergy Crop Sida hermaphrodita on Marginal Soil. Front. Plant Sci. 2018, 9, 9. [Google Scholar] [CrossRef]
- Solinas, S.; Fazio, S.; Seddaiu, G.; Roggero, P.P.; Deligios, P.A.; Doro, L.; Ledda, L. Environmental consequences of the conversion from traditional to energy cropping systems in a mediterranean area. Eur. J. Agron. 2015, 70, 124–135. [Google Scholar] [CrossRef]
- Herrmann, C.; Idler, C.; Heiermann, M. Biogas crops grown in energy crop rotations: Linking chemical composition and methane production characteristics. Bioresour. Technol. 2016, 206, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Popp, D.; Schrader, S.; Kleinsteuber, S.; Harms, H.; Strauber, H. Biogas production from coumarin-rich plants--inhibition by coumarin and recovery by adaptation of the bacterial community. FEMS Microbiol. Ecol. 2015, 91, fiv103. [Google Scholar] [CrossRef] [PubMed]
- Kadankova, P.; Kintl, A.; Koukalova, V.; Kucerova, J.; Brtnicky, M. Coumarin content in silages made of mixed cropping biomass comprising maize and white sweet clover. In Proceedings of the 19th International Multidisciplinary Scientific GeoConference SGEM2019, Sofia, Bulgaria, 30 June–6 July 2019. [Google Scholar]
- Kintl, A.; Elbl, J.; Vítěz, T.; Brtnický, M.; Skládanka, J.; Hammerschmiedt, T.; Vítězová, M. Possibilities of Using White Sweetclover Grown in Mixture with Maize for Biomethane Production. Agronomy 2020, 10, 1407. [Google Scholar] [CrossRef]
- Wojcieszak, M.; Pyzik, A.; Poszytek, K.; Krawczyk, P.S.; Sobczak, A.; Lipinski, L.; Roubinek, O.; Palige, J.; Sklodowska, A.; Drewniak, L. Adaptation of Methanogenic Inocula to Anaerobic Digestion of Maize Silage. Front. Microbiol. 2017, 8, 1881. [Google Scholar] [CrossRef]
- Cortesi, A. Assessing the Synergistic Effects of Co-digestion of Maize Silage and Red Chicory Waste. Chem. Biochem. Eng. Q. 2018, 32, 383–390. [Google Scholar] [CrossRef]
- Larsen, S.U.; Hjort-Gregersen, K.; Vazifehkhoran, A.H.; Triolo, J.M. Co-ensiling of straw with sugar beet leaves increases the methane yield from straw. Bioresour. Technol. 2017, 245 Pt A, 106–115. [Google Scholar] [CrossRef]
- Vitez, T.; Dokulilova, T.; Vitezova, M.; Elbl, J.; Kintl, A.; Kynicky, J.; Hladky, J.; Brtnicky, M. The Digestion of Waste from Vegetables and Maize Processing. Waste Biomass Valorization 2020, 11, 2467–2473. [Google Scholar] [CrossRef]
- Sukačová, K.; Búzová, D.; Trávníček, P.; Červený, J.; Vítězová, M.; Vítěz, T. Optimization of microalgal growth and cultivation parameters for increasing bioenergy potential: Case study using the oleaginous microalga Chlorella pyrenoidosa Chick (IPPAS C2). Algal Res. 2019, 40, 101519. [Google Scholar] [CrossRef]
- Cherubini, F. The biorefinery concept: Using biomass instead of oil for producing energy and chemicals. Energy Convers. Manag. 2010, 51, 1412–1421. [Google Scholar] [CrossRef]
- Johansson, E.; Prade, T.; Angelidaki, I.; Svensson, S.E.; Newson, W.R.; Gunnarsson, I.B.; Hovmalm, H.P. Economically viable components from Jerusalem artichoke (Helianthus tuberosus L.) in a biorefinery concept. Int. J. Mol. Sci. 2015, 16, 8997–9016. [Google Scholar] [CrossRef]
- Shao, T.; Long, X.; Liu, Y.; Gao, X.; Liu, M.; Rengel, Z. Effect of industrial crop Jerusalem artichoke on the micro-ecological rhizosphere environment in saline soil. Appl. Soil Ecol. 2022, 166, 104080. [Google Scholar] [CrossRef]
- Kays, S.J.; Nottingham, S.F. Biology and Chemistry of Jerusalem Artichoke: Helianthus tuberosus L., 1st ed.; CRC Press: Boca Raton, FL, USA, 2007; p. 496. [Google Scholar]
- Ţîţei, V.; Teleuţă, A.; Muntean, A. The perspective of cultivation and utilization of the species Silphium perfoliatum L. and Helianthus tuberosus L. in Moldova. Bulletin of the University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca. Agriculture 2013, 70, 160–166. [Google Scholar]
- Heuzé, V.; Tran, G.; Chapoutot, P.; Bastianelli, D.; Lebas, F. Jerusalem artichoke (Helianthus tuberosus). Feedipedia, aprogramme by INRA. 2015. Available online: https://www.feedipedia.org/node/544 (accessed on 8 November 2022).
- Bogucka, B.; Jankowski, K.J. The effect of harvest strategy on the energy potential of Jerusalem artichoke. Ind. Crop. Prod. 2022, 177, 114473. [Google Scholar] [CrossRef]
- Baldini, M.; Danuso, F.; Monti, A.; Amaducci, M.T.; Stevanato, P.; Mastro, G. Chichory and Jerusalem artichoke productivity in different areas of Italy, in relation to water availability and time of harvest. Ital. J. Agron. Riv. Agron. 2006, 1, 291–307. [Google Scholar] [CrossRef]
- Szewczyk, A.; Zagaja, M.; Bryda, J.; Kosikowska, U.; Stępień-Pyśniak, D.; Winiarczyk, D.; Andres-Mach, M. Topinambur—New possibilities for use in a supplementation diet. AAEM 2019, 26, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Hay, R.K.M.; Offer, R.W. Helianthus tuberosus as an alternative forage crop for cool maritime regions: A preliminary study of the yield and nutritional quality of shoot tissues from perennial stands. J. Sci. Food Agric. 1992, 60, 213–221. [Google Scholar] [CrossRef]
- Gunnarson, S.; Malmberg, A.; Mathisen, B.; Theander, O.; Thyselius, L.; Wünsche, U. Jerusalem artichoke (Helianthus tuberosus L.) for biogas production. Biomass 1985, 7, 85–97. [Google Scholar] [CrossRef]
- Emmerling, C.; Barton, J. Anaerobic co-digestion of topinambour (Helianthus tuberosus L.) and properties of the remaining biogas manure. Arch. Acker Pflanzenbau Bodenkd. 2007, 53, 683–690. [Google Scholar] [CrossRef]
- ČSN EN 15934; Sludge, Treated Biowaste, Soil and Waste—Calculation of Dry Matter Fraction After Determination of Dry Residue or Water Content. European Committee for Standardization: Brussel, Belgium, 2012.
- ČSN EN 15935; Sludge, Treated Biowaste, Soil and Waste—Determination of Loss on Ignition. European Committee for Standardization: Brussel, Belgium, 2012.
- ISO 20483:2013; Cereals and Pulses—Determination of the Nitrogen Content and Calculation of the Crude Protein Content—Kjeldahl Method—Kjeldahl Method. European Committee for Standardization: Brussel, Belgium, 2013.
- ČSN EN ISO 13906; Animal Feeding Stuffs—Determination of Acid Detergent Fibre (ADF) and Acid Detergent Lignin (ADL) Contents. European Committee for Standardization: Brussel, Belgium, 2008.
- Hoflliger, C.; Alves, M.; Andrade, D.; Angelidaki, I.; Astals, S.; Baier, U.; Bougrier, C.; Buffière, P.; Carballa, M.; De Wilde, V.; et al. Towards a standardization of biomethane potential tests. Water Sci. Technol. 2016, 74, 2515–2522. [Google Scholar] [CrossRef]
- Pichler, M.; Coskun, Ö.K.; Ortega-Arbulú, A.S.; Conci, N.; Wörheide, G.; Vargas, S.; Orsi, W.D. A 16S rRNA gene sequencing and analysis protocol for the Illumina MiniSeq platform. MicrobiologyOpen 2018, 7, e00611. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2010, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A communal catalogue reveals Earth’s multiscale microbial diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Callahan, B.J.; Sankaran, K.; Fukuyama, J.A.; McMurdie, P.J.; Holmes, S.P. Bioconductor Workflow for Microbiome Data Analysis: From raw reads to community analyses. F1000Research 2016, 5, 1492. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Ţiţei, V. The biochemical composition and the feed value of green mass and silage from Cynara cardunculus and Helianthus tuberosus in the Republic of Moldova. In Scientific Papers. Series D. Animal Science; University of Agronomic Sciences & Veterinary Medicine Bucharest: Bucharest, Romania, 2020; pp. 122–127. [Google Scholar]
- Wang, Y.; Zhao, Y.; Xue, F.; Nan, X.; Wang, H.; Hua, D.; Liu, J.; Yang, L.; Jiang, L.; Xiong, B. Nutritional value, bioactivity, and application potential of Jerusalem artichoke (Helianthus tuberosus L.) as a neotype feed resource. Anim. Nutr. 2020, 6, 429–437. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Dairy Cattle; National Research Council: Washington, DC, USA, 2001.
- Papi, N.; Kafilzadeh, F.; Fazaeli, H. Effects of incremental substitution of maize silage with Jerusalem artichoke silage on performance of fat-tailed lambs. Small Rumin. Res. 2017, 147, 56–62. [Google Scholar] [CrossRef]
- Liu, H.Y.; Wang, X.F.; Wang, Y.J.; Huang, X.; Li, Q.; Liu, P. Study on mixed silage of corn straw and Jerusalem artichoke stalk as feed sources. China Dairy Cattle 2017, 12, 20–23. [Google Scholar]
- Razmkhah, M.; Rezaei, J.; Fazaeli, H. Use of Jerusalem artichoke tops silage to replace corn silage in sheep diet. Anim. Feed Sci. Technol. 2017, 228, 168–177. [Google Scholar] [CrossRef]
- Loughrin, J.; Antle, S.; Bryant, M.; Berry, Z.; Lovanh, N. Evaluation of Microaeration and Sound to Increase Biogas Production from Poultry Litter. Environments 2020, 7, 62. [Google Scholar] [CrossRef]
- Triolo, J.M.; Sommer, S.G.; Møller, H.B.; Weisbjerg, M.R.; Jiang, X.Y. A new algorithm to characterize biodegradability of biomass during anaerobic digestion: Influence of lignin concentration on methane production potential. Bioresour. Technol. 2011, 102, 9395–9402. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, R.; Liu, X.; Chen, C.; Xiao, X.; Feng, L.; He, Y.; Liu, G. Evaluating Methane Production from Anaerobic Mono- and Co-digestion of Kitchen Waste, Corn Stover, and Chicken Manure. Energy Fuels 2013, 27, 2085–2091. [Google Scholar] [CrossRef]
- Ozbayram, E.G.; Ince, O.; Ince, B.; Harms, H.; Kleinsteuber, S. Comparison of Rumen and Manure Microbiomes and Implications for the Inoculation of Anaerobic Digesters. Microorganisms 2018, 6, 15. [Google Scholar] [CrossRef]
- Fuksa, P.; Hakl, J.; Míchal, P.; Hrevušová, Z.; Šantrůček, J.; Tlustoš, P. Effect of silage maize plant density and plant parts on biogas production and composition. Biomass Bioenergy 2020, 142, 105770. [Google Scholar] [CrossRef]
- Zubr, J. Methanogenic fermentation of fresh and ensiled plant materials. Biomass 1986, 11, 159–171. [Google Scholar] [CrossRef]
- Lehtomäki, A.; Viinikainen, T.A.; Rintala, J.A. Screening boreal energy crops and crop residues for methane biofuel production. Biomass Bioenergy 2008, 32, 541–550. [Google Scholar] [CrossRef]
- Amon, T.; Amon, B.; Kryvoruchko, V.; Machmüller, A.; Hopfner-Sixt, K.; Bodiroza, V.; Hrbek, R.; Friedel, J.; Pötsch, E.; Wagentristl, H.; et al. Methane production through anaerobic digestion of various energy crops grown in sustainable crop rotations. Bioresour. Technol. 2007, 98, 3204–3212. [Google Scholar] [CrossRef]
- Ţîţei, V.; Coșman, S. Biochemical characteristics of the Asteraceae species silage and possible use as a feedstock for livestock and biogas production in Republic of Moldova. RJAS 2016, 48, 105–112. [Google Scholar]
- Hutňan, M.; Špalková, V.; Bodík, I.; Kolesárová, N.; Lazor, M. Biogas Production from Maize Grains and Maize Silage. Pol. J. Environ. Stud. 2010, 19, 323–329. [Google Scholar]
- Dyksma, S.; Jansen, L.; Gallert, C. Syntrophic acetate oxidation replaces acetoclastic methanogenesis during thermophilic digestion of biowaste. Microbiome 2020, 8, 105. [Google Scholar] [CrossRef]
- Amin, F.R.; Khalid, H.; El-Mashad, H.; Chen, C.; Liu, G.; Zhang, R. Functions of bacteria and archaea participating in the bioconversion of organic waste for methane production. Sci. Total Environ. 2020, 763, 143007. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Toyonaga, M.; Ohashi, A.; Tourlousse, D.M.; Matsuura, N.; Meng, X.-Y.; Tamaki, H.; Hanada, S.; Cruz, R.; Yamaguchi, T.; et al. Lentimicrobium saccharophilum gen. nov., sp. nov., a strictly anaerobic bacterium representing a new family in the phylum Bacteroidetes, and proposal of Lentimicrobiaceae fam. nov. Int. J. Syst. Evol. 2016, 66, 2635–2642. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Tamayo, R.; Popova, M.; Tillier, M.; Morgavi, D.P.; Morel, J.P.; Fonty, G.; Morel-Desrosiers, N. Hydrogenotrophic methanogens of the mammalian gut: Functionally similar, thermodynamically different—A modelling approach. PLoS ONE 2019, 14, e0226243. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Soil Reaction (pH) | Plant Available Nutrient Content (mg/kg) | |||
|---|---|---|---|---|---|
| P | K | Ca | Mg | ||
| Arable Soil | 5.9 ± 0.23 | 95 ± 4.7 | 246 ± 39.4 | 1271 ± 64.7 | 135 ± 17.0 |
| Silage | Abbreviation | Percentage of Maize in Silage | Percentage of Jerusalem Artichoke in Silage | Average Density of Model Silage in DM (kg/m3) |
|---|---|---|---|---|
| Maize | M100 | 100 | 0 | 173.9 |
| Jerusalem Artichoke | JA100 | 0 | 100 | 156.0 |
| Maize + Jerusalem Artichoke | M30JA70 | 30 | 70 | 166.6 |
| Maize + Jerusalem Artichoke | M50JA50 | 50 | 50 | 150.2 |
| Maize + Jerusalem Artichoke | M70JA30 | 70 | 30 | 168.0 |
| Parameter | Value |
|---|---|
| Inoculum: | |
| Collection point | BPS Čejč, Czech Republic; input raw material—maize silage and slurry; mesophilic temperature conditions (38 °C); |
| Dry matter [%] | 3.54 ± 0.07 |
| Volatile solids [%] | 69.73 ± 0.11 |
| pH | 7.2 |
| FOS [mg/L] | 722 |
| TAC [mg/L] | 2090 |
| NH4+ [mg/L] | 624 |
| Biomethane potential test: | |
| Fermenter volume | Total volume 5 dm3; working volume 3 dm3 |
| temperature [°C]; heating method | 40 °C ± 0.2 °C; water bath |
| mixing | manually; daily |
| Retention time [day] | 35 |
| Silage dose [g] | 50–53 |
| Inoculum to Substrate (I/S) ratio | 4.5–4.9, based on VS content |
| Method of measuring biogas production | Method of liquid expansion according to standard VDI 4630 |
| Method of measuring biogas composition | Gas analyzer Dräger X-am 8000; infrared sensors for CH4 and CO2, mixture of gases (60% CH4/40% CO2) used as calibration gas |
| Number of repetitions | Three for each silage sample |
| No. | Variant | DM | NDF | ADF | CF | ||||
|---|---|---|---|---|---|---|---|---|---|
| % | HSD | %DM ± SD | HSD | %DM ± SD | HSD | %DM ± SD | HSD | ||
| 1 | M100 | 33.60 ± 0.50 | A | 43.80 ± 0.31 | A | 21.14 ± 2.05 | C | 16.82 ± 0.37 | D |
| 2 | M70 + JA30 | 33.44 ± 0.47 | A | 41.64 ± 0.85 | B | 23.91 ± 0.93 | BC | 17.73 ± 0.15 | CD |
| 3 | M50 + JA50 | 32.52 ± 0.75 | AB | 39.33 ± 0.80 | C | 25.63 ± 0.45 | AB | 18.58 ± 0.70 | BC |
| 4 | M30 + JA70 | 31.51 ± 0.56 | BC | 36.83 ± 0.14 | D | 26.31 ± 0.05 | AB | 19.64 ± 0.10 | AB |
| 5 | JA100 | 30.58 ± 0.30 | C | 33.16 ± 0.41 | E | 27.81 ± 0.28 | A | 19.81 ± 0.52 | A |
| Silage | DM | CP | NDF | ADF | ADL |
|---|---|---|---|---|---|
| Jerusalem artichoke, (JA100) 1 | 30.6 | 5.1 | 33.2 | 27.8 | 8.1 |
| Jerusalem artichoke (squaring stage) 2 | 25.8 | 11.6 | 43.9 | 31.7 | 7.5 |
| Yellow maize, normal, (M100) 1 | 33.6 | 9.3 | 43.8 | 21.1 | 2.3 |
| Yellow maize, normal (32% to 38% DM) 3 | 35.1 | 8.8 | 45.0 | 28.1 | 2.6 |
| Barley 3 | 35.5 | 12.0 | 56.3 | 34.5 | 5.6 |
| Oats (head) 3 | 34.6 | 12.9 | 60.6 | 38.9 | 5.5 |
| Ryegrass (annual, vegetative stage) 3 | 29.7 | 16.1 | 57.8 | 34.9 | 4.5 |
| Wheat (head) 3 | 33.3 | 12.0 | 59.9 | 37.6 | 5.8 |
| No. | Variant | Starch | CP | ADL | Lipids | ||||
|---|---|---|---|---|---|---|---|---|---|
| %DM ± SD | HSD | %DM ± SD | HSD | %DM ± SD | HSD | %DM ± SD | HSD | ||
| 1 | M100 | 16.14 ± 0.00 | A | 9.30 ± 0.13 | A | 2.32 ± 0.12 | C | 4.36 ± 0.09 | A |
| 2 | M70 + JA30 | 15.26 ± 0.38 | B | 8.33 ± 0.14 | B | 3.50 ± 0.27 | BC | 3.49 ± 0.13 | B |
| 3 | M50 + JA50 | 13.54 ± 0.06 | C | 8.04 ± 0.14 | B | 4.88 ± 0.68 | B | 3.41 ± 0.20 | B |
| 4 | M30 + JA70 | 10.09 ± 0.07 | D | 7.33 ± 0.05 | C | 7.37 ± 1.33 | A | 3.37 ± 0.17 | BC |
| 5 | JA100 | 9.17 ± 0.09 | E | 5.08 ± 0.13 | D | 8.08 ± 0.31 | A | 2.89 ± 0.27 | C |
| No. | Variant | Methane Yield | Theoretical Methane Yield | Methane Content * | ||
|---|---|---|---|---|---|---|
| Nm3/kgVS | HSD | m3/kgVS | %vol | HSD | ||
| 1 | M100 | 0.352 ± 0.017 | A | 0.427 | 65 ± 0.06 | A |
| 2 | M70 + JA30 | 0.287 ± 0.014 | B | 0.412 | 65 ± 0.05 | A |
| 3 | M50 + JA50 | 0.302 ± 0.015 | B | 0.400 | 65 ± 0.06 | A |
| 4 | M30 + JA70 | 0.261 ± 0.013 | B | 0.374 | 65 ± 0.05 | A |
| 5 | JA100 | 0.274 ± 0.013 | B | 0.346 | 64 ± 0.06 | A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kintl, A.; Vítěz, T.; Huňady, I.; Brtnický, M.; Novák, D.; Lochman, J.; Vítězová, M.; Hammerschmiedt, T.; Holátko, J.; Elbl, J. Anaerobic Fermentation of Silage from the Above-Ground Biomass of Jerusalem Artichoke (Helianthus tuberosus L.) and Maize (Zea mayse L.) as a New and Promising Input Raw Material for Biogas Production. Appl. Sci. 2022, 12, 11417. https://doi.org/10.3390/app122211417
Kintl A, Vítěz T, Huňady I, Brtnický M, Novák D, Lochman J, Vítězová M, Hammerschmiedt T, Holátko J, Elbl J. Anaerobic Fermentation of Silage from the Above-Ground Biomass of Jerusalem Artichoke (Helianthus tuberosus L.) and Maize (Zea mayse L.) as a New and Promising Input Raw Material for Biogas Production. Applied Sciences. 2022; 12(22):11417. https://doi.org/10.3390/app122211417
Chicago/Turabian StyleKintl, Antonín, Tomáš Vítěz, Igor Huňady, Martin Brtnický, David Novák, Jan Lochman, Monika Vítězová, Tereza Hammerschmiedt, Jiří Holátko, and Jakub Elbl. 2022. "Anaerobic Fermentation of Silage from the Above-Ground Biomass of Jerusalem Artichoke (Helianthus tuberosus L.) and Maize (Zea mayse L.) as a New and Promising Input Raw Material for Biogas Production" Applied Sciences 12, no. 22: 11417. https://doi.org/10.3390/app122211417
APA StyleKintl, A., Vítěz, T., Huňady, I., Brtnický, M., Novák, D., Lochman, J., Vítězová, M., Hammerschmiedt, T., Holátko, J., & Elbl, J. (2022). Anaerobic Fermentation of Silage from the Above-Ground Biomass of Jerusalem Artichoke (Helianthus tuberosus L.) and Maize (Zea mayse L.) as a New and Promising Input Raw Material for Biogas Production. Applied Sciences, 12(22), 11417. https://doi.org/10.3390/app122211417