
Investigating the Anticancer Activity and Characterization of Bioactive Constituents of Moricandia sinaica (Boiss.) Boiss through In Vitro and In Silico Approaches in Triple-Negative Breast Cancer Cell Line

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant
2.2. Screening Crude Extracts for Anticancer Activity
2.3. GC/MS Analysis
2.4. Estimation of Total Phenol and Flavonoid Contents
2.5. Quantification of Apoptosis by Annexin V-FITC/(PI) Staining
2.6. Cell Cycle Analysis
2.7. Molecular Docking
2.8. Statistics
3. Results
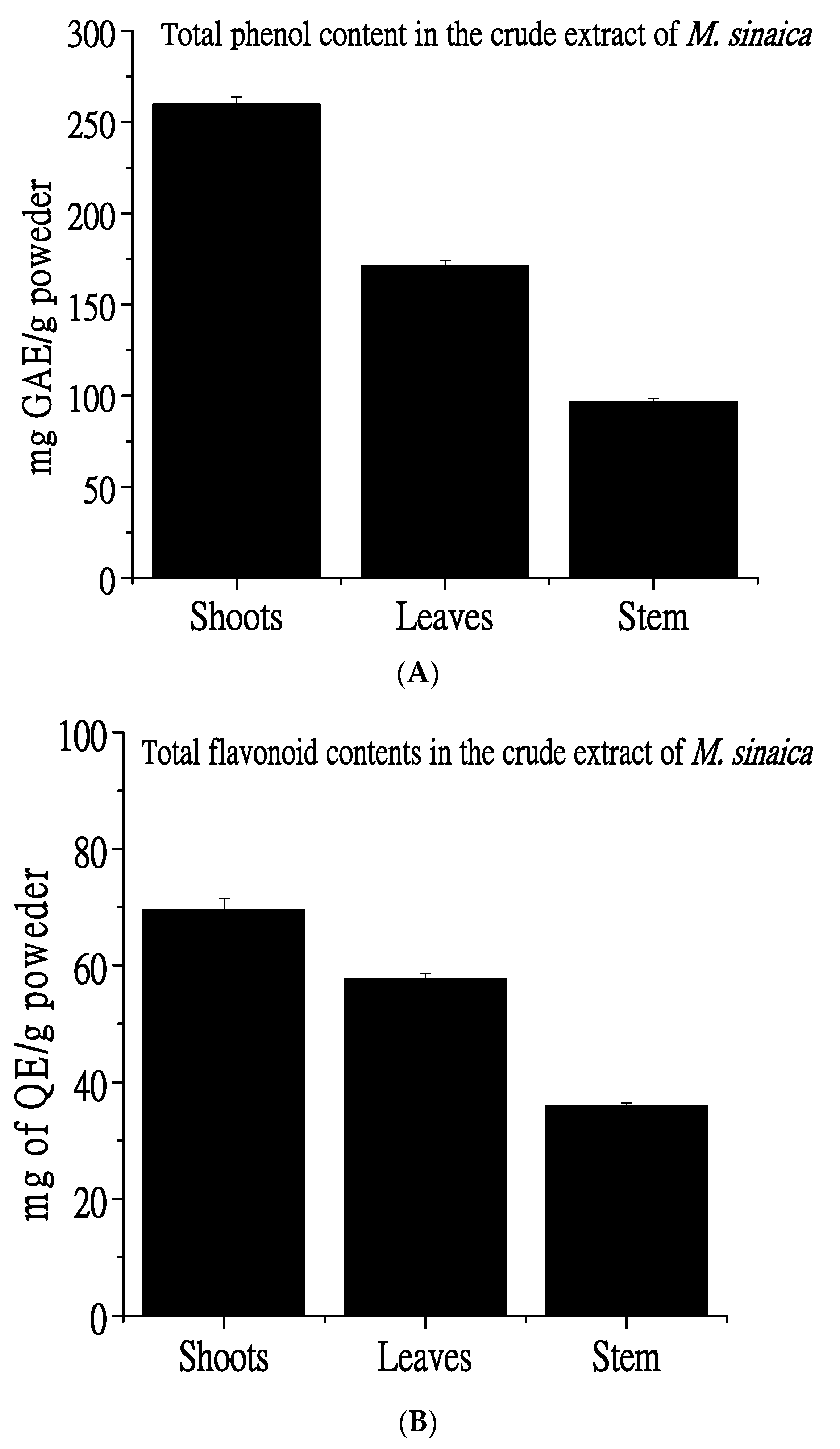
3.1. Total Phenolic and Flavonoid Analysis
3.2. Methanol Extract of the Shoots of M. sinaica Selectively Perturbed Cell Survival of Breast Cancer MDA-MB-231 Cells
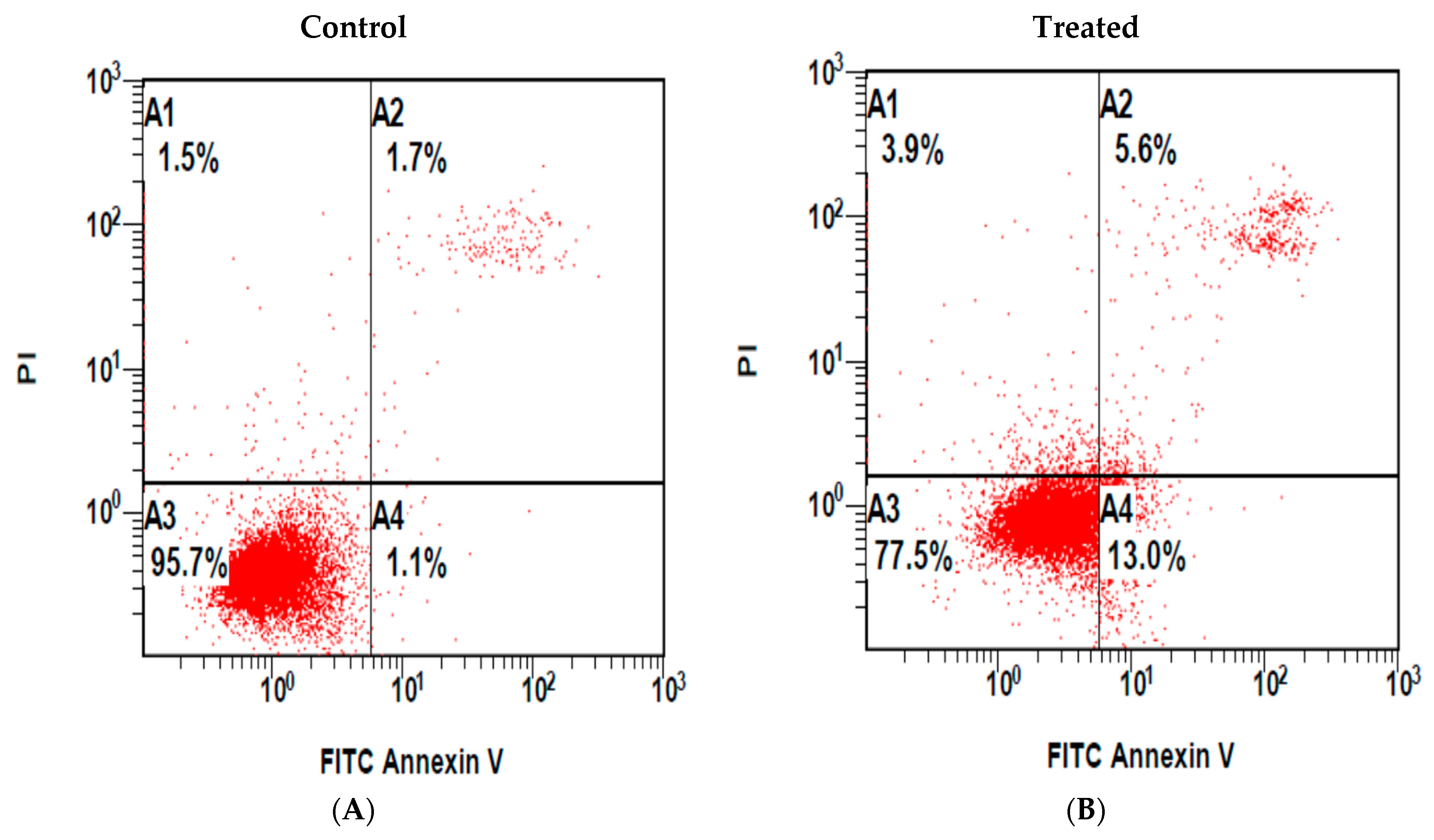
3.3. MeOH Extract of the Shoots of M. sinaica-Induced Apoptosis in MDA-MB-231 Cells
3.4. Methanol Extract of the Shoots of M. sinaica-Induced G1 Cell Cycle Arrest in MDA-MB-231 Cells
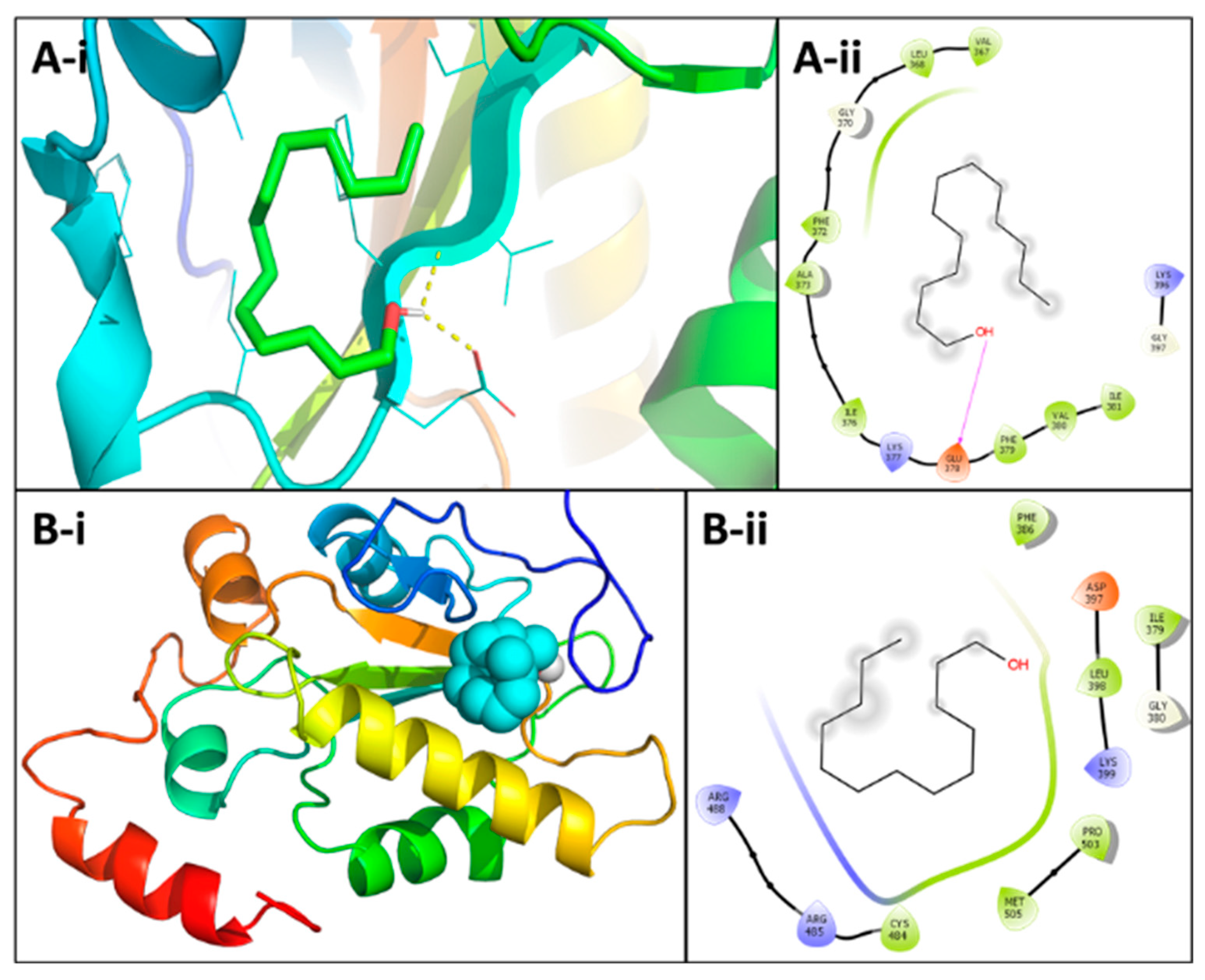
3.5. Identification of Phytochemicals and In Silico Molecular Docking Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, V.; Gouveia, M.J.; Santos, J.; Koksch, B.; Amorim, I.; Gartner, F.; Vale, N. Breast cancer: Insights in disease and influence of drug methotrexate. RSC Med. Chem. 2020, 11, 646–664. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Mosly, D.; Turnbull, A.; Sims, A.; Ward, C.; Langdon, S. Predictive markers of endocrine response in breast cancer. World J. Exp. Med. 2018, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bauer, K.R.; Brown, M.; Cress, R.D.; Parise, C.A.; Caggiano, V. Descriptive analysis of estrogen receptor (ER)-negative, progesterone receptor (PR)-negative, and HER2-negative invasive breast cancer, the so-called triple-negative phenotype: A population-based study from the California cancer Registry. Cancer 2007, 109, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Nedeljkovic, M.; Damjanovic, A. Mechanisms of Chemotherapy Resistance in Triple-Negative Breast Cancer-How We Can Rise to the Challenge. Cells 2019, 8, 957. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Li, A.; Inagaki, Y.; Gao, J.; Li, J.; Kokudo, N.; Li, X.K.; Tang, W. Chinese herbal medicines as adjuvant treatment during chemo- or radio-therapy for cancer. Biosci. Trends 2010, 4, 297–307. [Google Scholar]
- Liao, G.S.; Apaya, M.K.; Shyur, L.F. Herbal medicine and acupuncture for breast cancer palliative care and adjuvant therapy. Evid. Based Complement. Altern. Med. 2013, 2013, 437948. [Google Scholar] [CrossRef]
- Zheng, J.; Zhou, Y.; Li, Y.; Xu, D.P.; Li, S.; Li, H.B. Spices for Prevention and Treatment of Cancers. Nutrients 2016, 8, 495. [Google Scholar] [CrossRef]
- Xue, J.P.; Wang, G.; Zhao, Z.B.; Wang, Q.; Shi, Y. Synergistic cytotoxic effect of genistein and doxorubicin on drug-resistant human breast cancer MCF-7/Adr cells. Oncol. Rep. 2014, 32, 1647–1653. [Google Scholar] [CrossRef]
- Gonzalez-Vallinas, M.; Molina, S.; Vicente, G.; Sanchez-Martinez, R.; Vargas, T.; Garcia-Risco, M.R.; Fornari, T.; Reglero, G.; Ramirez de Molina, A. Modulation of estrogen and epidermal growth factor receptors by rosemary extract in breast cancer cells. Electrophoresis 2014, 35, 1719–1727. [Google Scholar] [CrossRef]
- Pratap, A.; Gupta, S.K. Biology and Ecology of Wild Crucifers; Gupta, S.K., Ed.; CRC Press: Boca Raton, FL, USA; Tayler and Francis Group: Boca Raton, FL, USA, 2009. [Google Scholar]
- Skandrani, I.; Leloup, L.; Kovacic, H.; Dijoux-Franca, M.G.; Ghedira, K.; Chekir Ghedira, L. Antioxidant, Antimutagenic, Tanning and Calpain Induction Activities of Methanolic Extract of Tunisian Plant (Moricandia Arvensis). Iran. J. Pharm. Res. 2017, 16, 119–134. [Google Scholar] [PubMed]
- Skandrani, I.; Sghaier, M.B.; Neffati, A.; Boubaker, J.; Bouhlel, I.; Kilani, S.; Mahmoud, A.; Ghedira, K.; Chekir-Ghedira, L. Antigenotoxic and free radical scavenging activities of extracts from Moricandia arvensis. Drug Chem. Toxicol. 2007, 30, 361–382. [Google Scholar] [CrossRef]
- Skandrani, I.; Limem, I.; Neffati, A.; Boubaker, J.; Ben Sghaier, M.; Bhouri, W.; Bouhlel, I.; Kilani, S.; Ghedira, K.; Chekir-Ghedira, L. Assessment of phenolic content, free-radical-scavenging capacity genotoxic and anti-genotoxic effect of aqueous extract prepared from Moricandia arvensis leaves. Food Chem. Toxicol. 2010, 48, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Skandrani, I.; Bouhlel, I.; Limem, I.; Boubaker, J.; Bhouri, W.; Neffati, A.; Ben Sghaier, M.; Kilani, S.; Ghedira, K.; Ghedira-Chekir, L. Moricandia arvensis extracts protect against DNA damage, mutagenesis in bacteria system and scavenge the superoxide anion. Toxicol. Vitro 2009, 23, 166–175. [Google Scholar] [CrossRef] [PubMed]
- El-Mekkawy, S.; Shahat, A.A.; Alqahtani, A.S.; Alsaid, M.S.; Abdelfattah, M.A.O.; Ullah, R.; Emam, M.; Yasri, A.; Sobeh, M. A Polyphenols-Rich Extract from Moricandia sinaica Boiss. Exhibits Analgesic, Anti-Inflammatory and Antipyretic Activities In Vivo. Molecules 2020, 25, 5049. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Morrone, F.; Argentieri, M.P.; Gambacorta, L.; Conforti, F.; Avato, P. Phytochemical and Biological Profile of Moricandia arvensis (L.) DC.: An Inhibitor of Pancreatic Lipase. Molecules 2018, 23, 2829. [Google Scholar] [CrossRef]
- D’Andrea, G. Quercetin: A flavonol with multifaceted therapeutic applications? Fitoterapia 2015, 106, 256–271. [Google Scholar] [CrossRef]
- Kashyap, D.; Sharma, A.; Tuli, H.S.; Sak, K.; Punia, S.; Mukherjee, T.K. Kaempferol—A dietary anticancer molecule with multiple mechanisms of action: Recent trends and advancements. J. Funct. Foods 2017, 30, 203–219. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Belkhiri, A.; Lockwood, G.B. An indole derivative and glucosinolates from Moricandia arvensis. Phytochemistry 1990, 29, 1315–1316. [Google Scholar] [CrossRef]
- Braham, H.; Mighri, Z.; Jannet, H.B.; Matthew, S.; Abreu, P.M. Antioxidant phenolic glycosides from Moricandia arvensis. J. Nat. Prod. 2005, 68, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Skandrani, I.; Boubaker, J.; Bhouri, W.; Limem, I.; Kilani, S.; Ben Sghaier, M.; Neffati, A.; Bouhlel, I.; Ghedira, K.; Chekir-Ghedira, L. Leaf extracts from Moricandia arvensis promote antiproliferation of human cancer cells, induce apoptosis, and enhance antioxidant activity. Drug Chem. Toxicol. 2010, 33, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Tatsuzawa, F.; Ito, K.; Muraoka, H.; Namauo, T.; Kato, K.; Takahata, Y.; Ogawa, S. Triacylated peonidin 3-sophoroside-5-glucosides from the purple flowers of Moricandia ramburii Webb. Phytochemistry 2012, 76, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.C.; Granieri, L.; Shrestha, M.; Wang, D.Y.; Vorobieva, I.; Rubie, E.A.; Jones, R.; Ju, Y.; Pellecchia, G.; Jiang, Z.; et al. Identification of CDC25 as a Common Therapeutic Target for Triple-Negative Breast Cancer. Cell Rep. 2018, 23, 112–126. [Google Scholar] [CrossRef]
- Kabakci, Z.; Kappeli, S.; Cantu, C.; Jensen, L.D.; Konig, C.; Toggweiler, J.; Gentili, C.; Ribaudo, G.; Zagotto, G.; Basler, K.; et al. Pharmacophore-guided discovery of CDC25 inhibitors causing cell cycle arrest and tumor regression. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Lee, A.V.; Oesterreich, S.; Davidson, N.E. MCF-7 cells--changing the course of breast cancer research and care for 45 years. J. Natl. Cancer Inst. 2015, 107. [Google Scholar] [CrossRef]
- Holliday, D.L.; Speirs, V. Choosing the right cell line for breast cancer research. Breast Cancer Res. 2011, 13, 215. [Google Scholar] [CrossRef]
- Chavez, K.J.; Garimella, S.V.; Lipkowitz, S. Triple negative breast cancer cell lines: One tool in the search for better treatment of triple negative breast cancer. Breast Dis. 2010, 32, 35–48. [Google Scholar] [CrossRef]
- Farooq, M.; Nasr, F.A.; Almoutiri, N.D.; Al-yahya, N.; Wadaan, M.A.; Abutaha, N. The phytochemical screening and antiangiogenic activity of audthan al himar (Moricandia sinaica Boiss.) extracts in zebrafish embryos and human umbilical vein endothelial cells. J. King Saud Univ. Sci. 2020, 32, 2370–2376. [Google Scholar] [CrossRef]
- Aref, H.L.; Karima, B.H.S.; Fekih, A.; Chemli, R.; Mars, M.; Aouni, M.; Chaumon, J.P.; Said, K. Variability in antimicrobial activity of latex from two varieties of Ficus carica. Afr. J. Microbiol. Res. 2011, 5, 1361–1367. [Google Scholar]
- Trott, O.; Olson, A.J. Software News and Update AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Akçay, A. The Calculation of LD50 Using Probit Analysis. FASEB J. 2013, 27. [Google Scholar] [CrossRef]
- Beillerot, A.; Battaglia, E.; Bennasroune, A.; Bagrel, D. Protection of CDC25 phosphatases against oxidative stress in breast cancer cells: Evaluation of the implication of the thioredoxin system. Free Radic. Res. 2012, 46, 674–689. [Google Scholar] [CrossRef] [PubMed]
- Cangi, M.G.; Cukor, B.; Soung, P.; Signoretti, S.; Moreira, G.; Ranashinge, M.; Cady, B.; Pagano, M.; Loda, M. Role of the Cdc25A phosphatase in human breast cancer. J. Clin. Investig. 2000, 106, 753–761. [Google Scholar] [CrossRef]
- Zacksenhaus, E.; Liu, J.C.; Granieri, L.; Vorobieva, I.; Wang, D.Y.; Ghanbari-Azarnier, R.; Li, H.Q.; Ali, A.; Chung, P.E.D.; Ju, Y.J.; et al. CDC25 as a common therapeutic target for triple-negative breast cancer—The challenges ahead. Mol. Cell Oncol. 2018, 5. [Google Scholar] [CrossRef]
- Dean, R.A.; Fam, H.K.; An, J.H.; Choi, K.H.; Shimizu, Y.; Jones, S.J.M.; Boerkoel, C.F.; Interthal, H.; Pfeifer, T.A. Identification of a Putative Tdp1 Inhibitor (CD00509) by in Vitro and Cell-Based Assays. J. Biomol. Screen. 2014, 19, 1372–1382. [Google Scholar] [CrossRef]
- Chen, C.L.; Chu, J.S.; Su, W.C.; Huang, S.C.; Lee, W.Y. Hypoxia and metabolic phenotypes during breast carcinogenesis: Expression of HIF-1 alpha, GLUT1, and CAIX. Virchows Arch. 2010, 457, 53–61. [Google Scholar] [CrossRef]
- Murugesan Amudha, S.R. Assessing the Bioactive Constituents of Cadaba Fruticosa (L.) Druce Through Gc-Ms. Int. J. Pharm. Pharm. Sci. 2014, 6, 3. [Google Scholar]
- Ahmad, S.; Ullah, F.; Sadiq, A.; Ayaz, M.; Imran, M.; Ali, I.; Zeb, A.; Ullah, F.; Shah, M.R. Chemical composition, antioxidant and anticholinesterase potentials of essential oil of Rumex hastatus D. Don collected from the North West of Pakistan. BMC Complement. Altern. Med. 2016, 16. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Evain-Bana, E.; Schiavo, L.; Bour, C.; Lanfranchi, D.A.; Berardozzi, S.; Ghirga, F.; Bagrel, D.; Botta, B.; Hanquet, G.; Mori, M. Synthesis, biological evaluation and molecular modeling studies on novel quinonoid inhibitors of CDC25 phosphatases. J. Enzym. Inhib. Med. Chem. 2017, 32, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Moaty, H.I. Bioactive Compounds of Moricandia Nitens and Its Anticancer Effect. Indo Am. J. Pharm Sci. 2016, 3, 1283–1290. [Google Scholar] [CrossRef]
- Aidi Wannes, W.; Saidani Tounsi, M.; Marzouk, B. A review of Tunisian medicinal plants with anticancer activity. J. Complement. Integr. Med. 2017, 15. [Google Scholar] [CrossRef]
- Perfectti, F.; Gomez, J.M.; Gonzalez-Megias, A.; Abdelaziz, M.; Lorite, J. Molecular phylogeny and evolutionary history of Moricandia DC (Brassicaceae). PeerJ 2017, 5, e3964. [Google Scholar] [CrossRef] [PubMed]
- Bhagya, N.; Chandrashekar, K.R. Identification and quantification of cytotoxic phenolic acids and flavonoids in Ixora brachiata by UHPLC-DAD and UHPLC-ESI-MS/MS. Int. J. Mass Spectrom. 2020, 450. [Google Scholar] [CrossRef]
- Paudel, M.R.; Chand, M.B.; Pant, B.; Pant, B. Assessment of Antioxidant and Cytotoxic Activities of Extracts of Dendrobium crepidatum. Biomolecules 2019, 9, 478. [Google Scholar] [CrossRef]
- Taraphdar, A.; Roy, M.; Bhattacharya, R. Natural products as inducers of apoptosis: Implication for cancer therapy and prevention. Curr. Sci. 2001, 80, 1387–1396. [Google Scholar]
- Jackson, R.C.; Di Veroli, G.Y.; Koh, S.B.; Goldlust, I.; Richards, F.M.; Jodrell, D.I. Modelling of the cancer cell cycle as a tool for rational drug development: A systems pharmacology approach to cyclotherapy. PLoS Comput. Biol. 2017, 13. [Google Scholar] [CrossRef]
- Saleh, K.A.; Albinhassan, T.H.; Elbehairi, S.E.I.; Alshehry, M.A.; Alfaifi, M.Y.; Al-Ghazzawi, A.M.; Al-Kahtani, M.A.; Alasmari, A.D.A. Cell Cycle Arrest in Different Cancer Cell Lines (Liver, Breast, and Colon) Induces Apoptosis under the Influence of the Chemical Content of Aeluropus lagopoides Leaf Extracts. Molecules 2019, 24, 507. [Google Scholar] [CrossRef]
- Uzzan, B.; Nicolas, P.; Cucherat, M.; Perret, G.Y. Microvessel density as a prognostic factor in women with breast cancer: A systematic review of the literature and meta-analysis. Cancer Res. 2004, 64, 2941–2955. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Part | Extract | IC50 in Human Cancer and Normal Cell Lines (µg/mL) | |||
|---|---|---|---|---|---|
| HepG2 | MDA-MB-231 | MCF7 | HUVEC | ||
| Stem | Methanol | NA | 400.56 | NA | NA |
| Ethyl acetate | NA | 866.66 | 758.27 | NA | |
| Chloroform | NA | 365.72 | 568.37 | NA | |
| Leaves | Methanol | 746.36 | 475.163 | NA | 245.34 |
| Ethyl acetate | 621.76 | 686.68 | NA | NA | |
| Chloroform | 324.731 | 568.79 | NA | 468.36 | |
| Shoots | Methanol | NA | 14.32 | 365.4 | NA |
| Ethyl acetate | NA | 48.93 | 786.1 | NA | |
| Chloroform | 863.97 | 43.98 | 359.73 | 754.98 | |
| Positive control | Doxorubicin | 0.88 | 4.47 | 2.91 | 1.7 |
| Compound | Structure | Retention Time (s) | % Area Covered |
|---|---|---|---|
| 2-Tridecen-1-ol |  | 20.85 | 34.87 |
| 7-Butyl-bicyclo [4.1.0]heptane |  | 21.34 | 7.41 |
| 1,2-Benzenedicarboxylic acid |  | 22.32 | 18.62 |
| 1,19-Eicosadiene |  | 22.67 | 7.27 |
| Hexadecanoic acid |  | 23.05 | 17.4 |
| Trans phytol |  | 23.81 | 15.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farooq Khan, M.; Nasr, F.A.; Baabbad, A.A.; Alqahtani, A.S.; Wadaan, M.A.M. Investigating the Anticancer Activity and Characterization of Bioactive Constituents of Moricandia sinaica (Boiss.) Boiss through In Vitro and In Silico Approaches in Triple-Negative Breast Cancer Cell Line. Appl. Sci. 2021, 11, 1244. https://doi.org/10.3390/app11031244
Farooq Khan M, Nasr FA, Baabbad AA, Alqahtani AS, Wadaan MAM. Investigating the Anticancer Activity and Characterization of Bioactive Constituents of Moricandia sinaica (Boiss.) Boiss through In Vitro and In Silico Approaches in Triple-Negative Breast Cancer Cell Line. Applied Sciences. 2021; 11(3):1244. https://doi.org/10.3390/app11031244
Chicago/Turabian StyleFarooq Khan, Muhammad, Fahd A. Nasr, Almohannad A. Baabbad, Ali S. Alqahtani, and Mohammad A. M. Wadaan. 2021. "Investigating the Anticancer Activity and Characterization of Bioactive Constituents of Moricandia sinaica (Boiss.) Boiss through In Vitro and In Silico Approaches in Triple-Negative Breast Cancer Cell Line" Applied Sciences 11, no. 3: 1244. https://doi.org/10.3390/app11031244
APA StyleFarooq Khan, M., Nasr, F. A., Baabbad, A. A., Alqahtani, A. S., & Wadaan, M. A. M. (2021). Investigating the Anticancer Activity and Characterization of Bioactive Constituents of Moricandia sinaica (Boiss.) Boiss through In Vitro and In Silico Approaches in Triple-Negative Breast Cancer Cell Line. Applied Sciences, 11(3), 1244. https://doi.org/10.3390/app11031244