
Establishment of an Aseptic Culture System and Analysis of the Effective Growth Conditions for Eleocharis acicularis Ramets for Use in Phytoremediation


Abstract
:1. Introduction
2. Materials and Methods
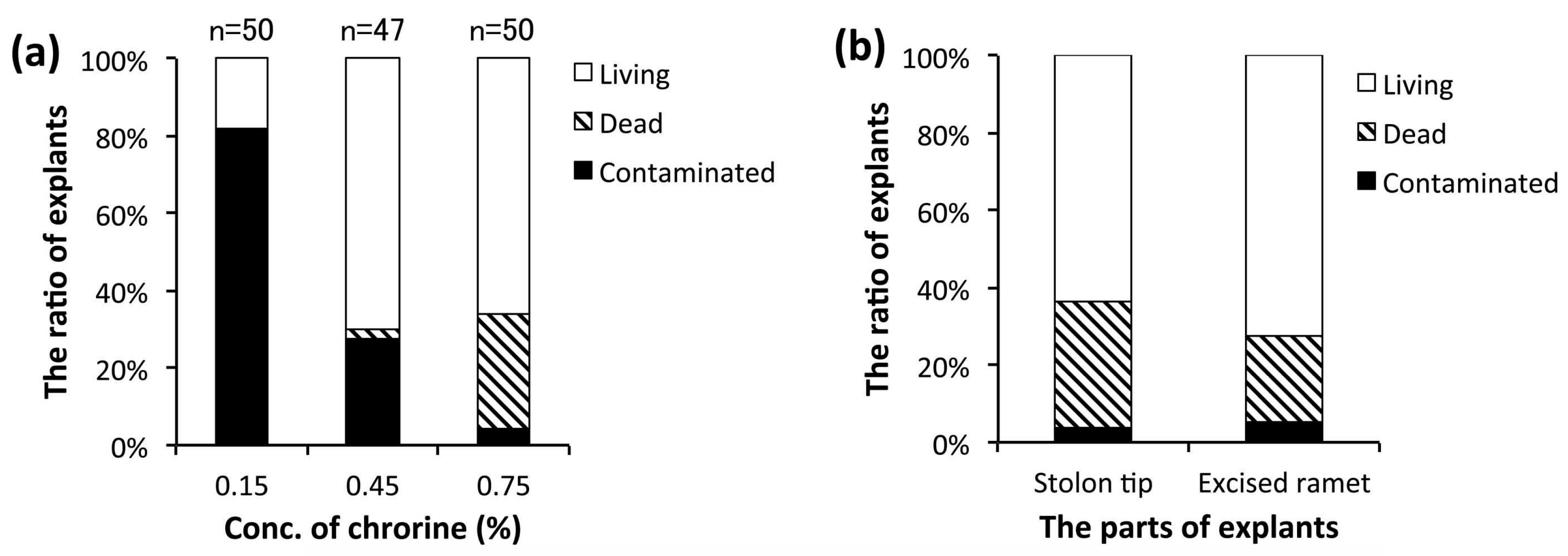
2.1. Plant Material and Aseptic Culture Conditions
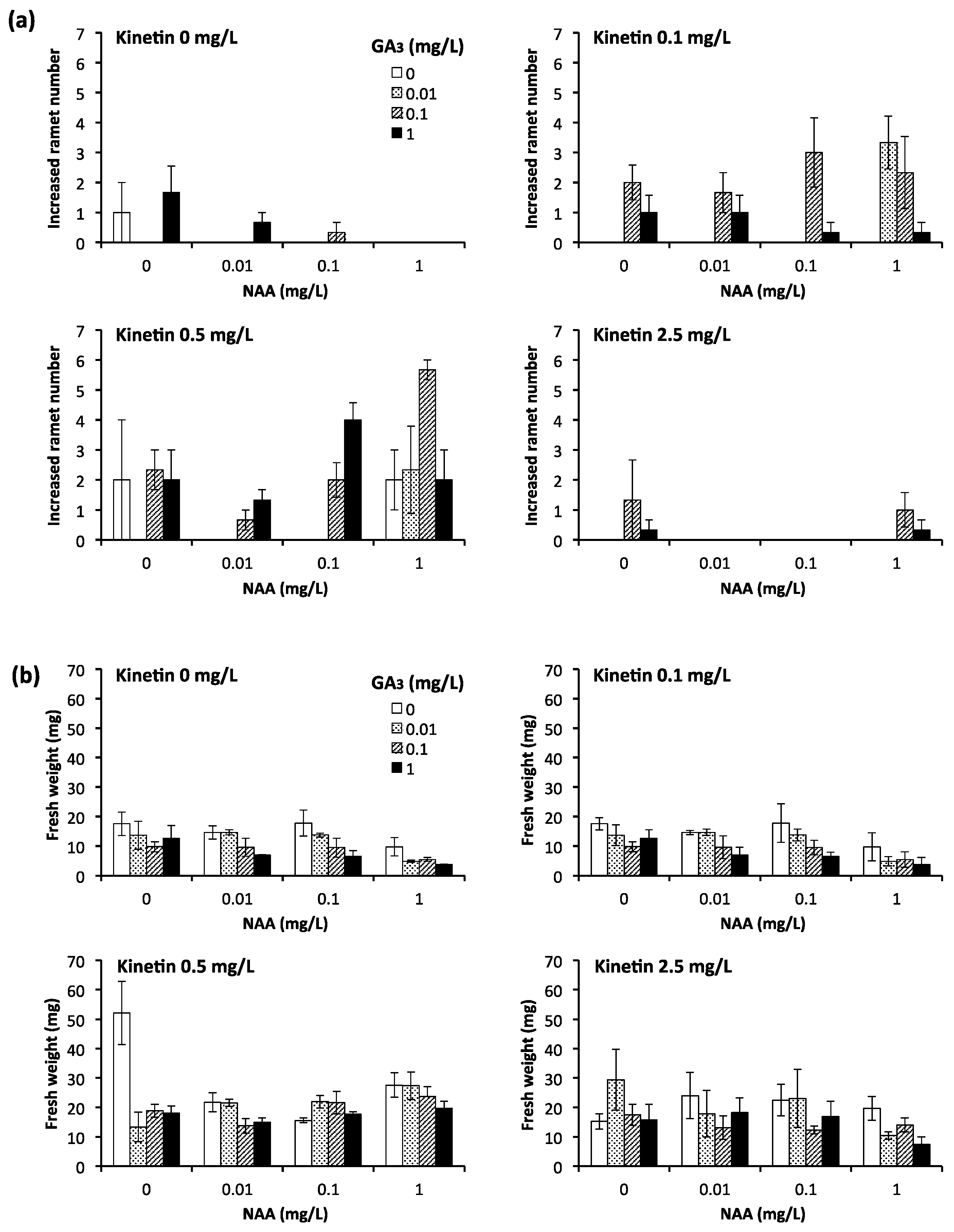
2.2. Analysis of the Effect of Plant Hormones and pH Conditions on the Growth of Young Ramets of E. acicularis
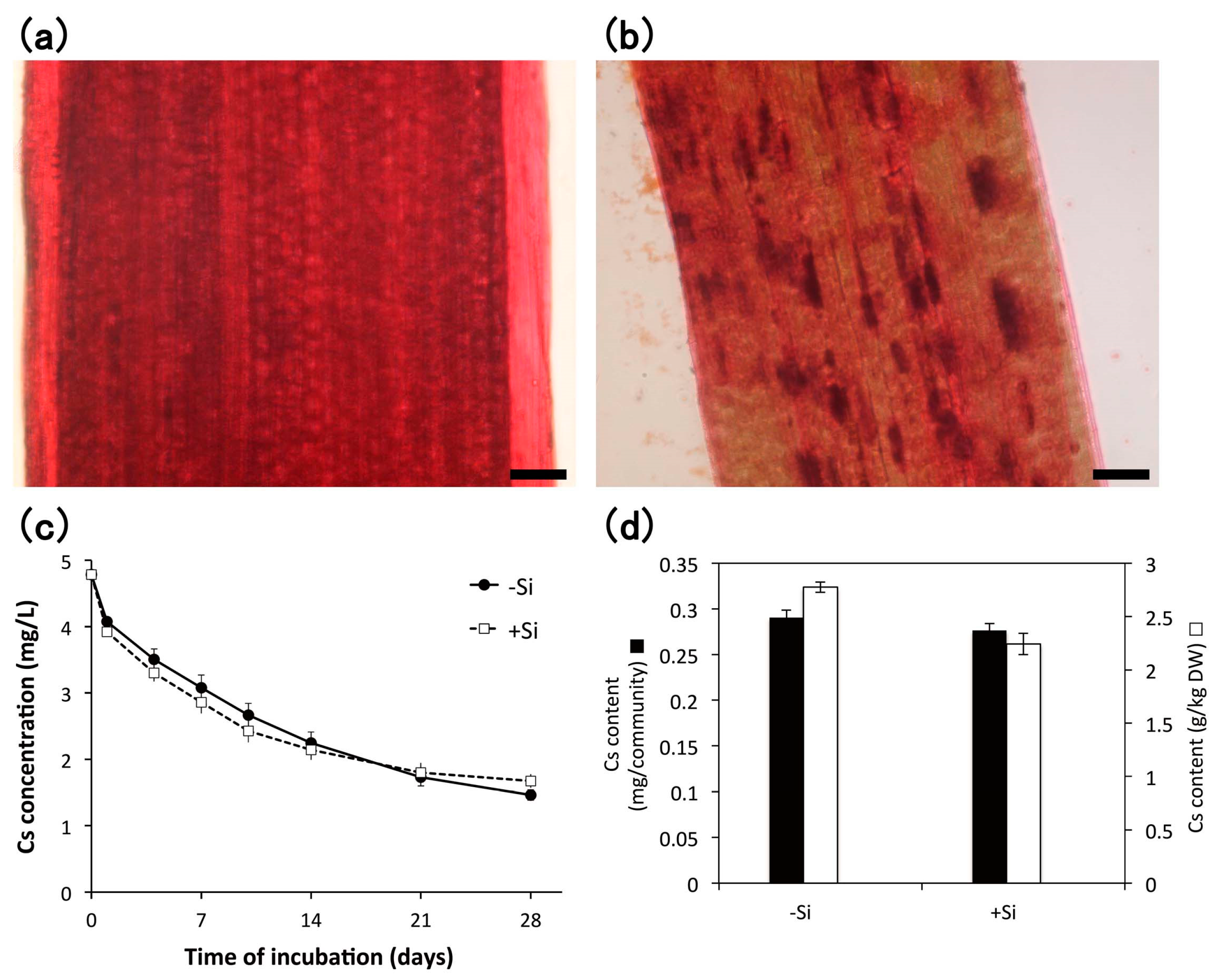
2.3. Preparation of E. acicularis Harboring Phytoliths
2.4. Staining of Phytoliths
2.5. Heavy Metal Preparation
2.6. Experimental Setup and Sampling for Analysis of Heavy Metal Accumulation in E. acicularis
2.7. Analytical Methods
2.7.1. Digestion of Plants for the Measurement of Cs, Zn, Pb, Cd, and In
2.7.2. ICP-MS Analysis
2.7.3. Atomic Absorption Spectrophotometry Analysis
2.7.4. ICP-AES Analysis
2.7.5. Analysis of Si Contents in Plants
2.8. BCF
2.9. Replicates and Statistical Analysis
3. Results
3.1. Morphological Features of E. acicularis
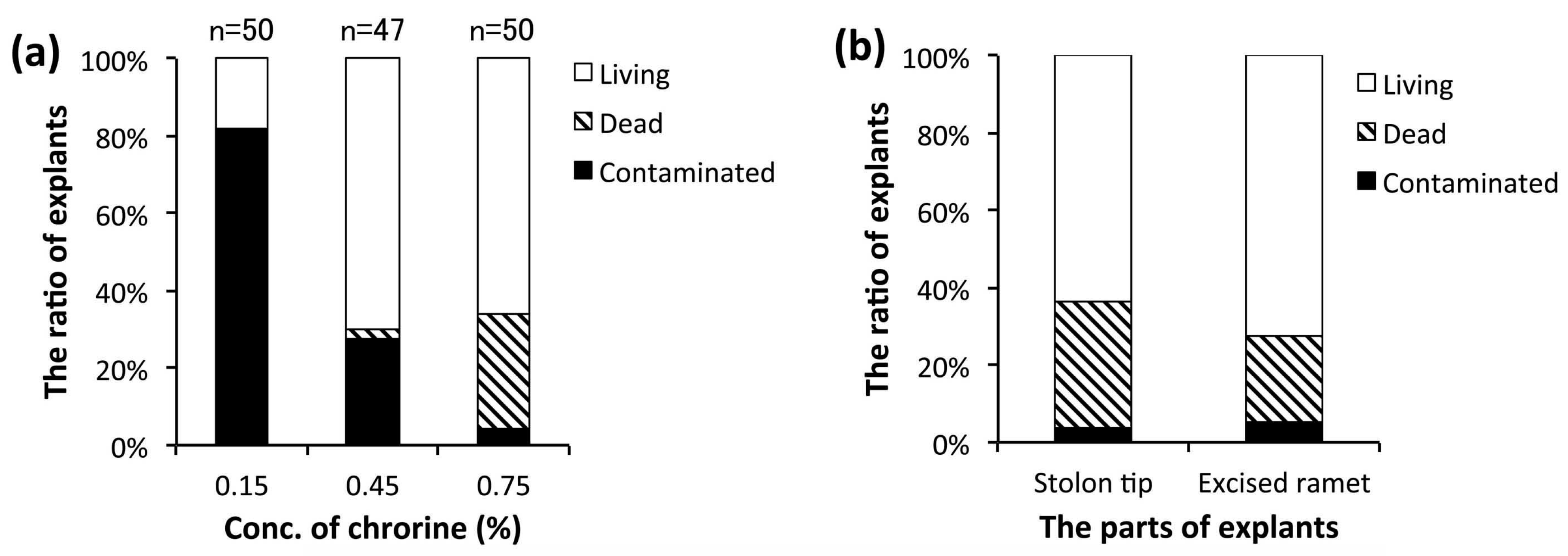
3.2. Establishment of an Aseptic Culture System
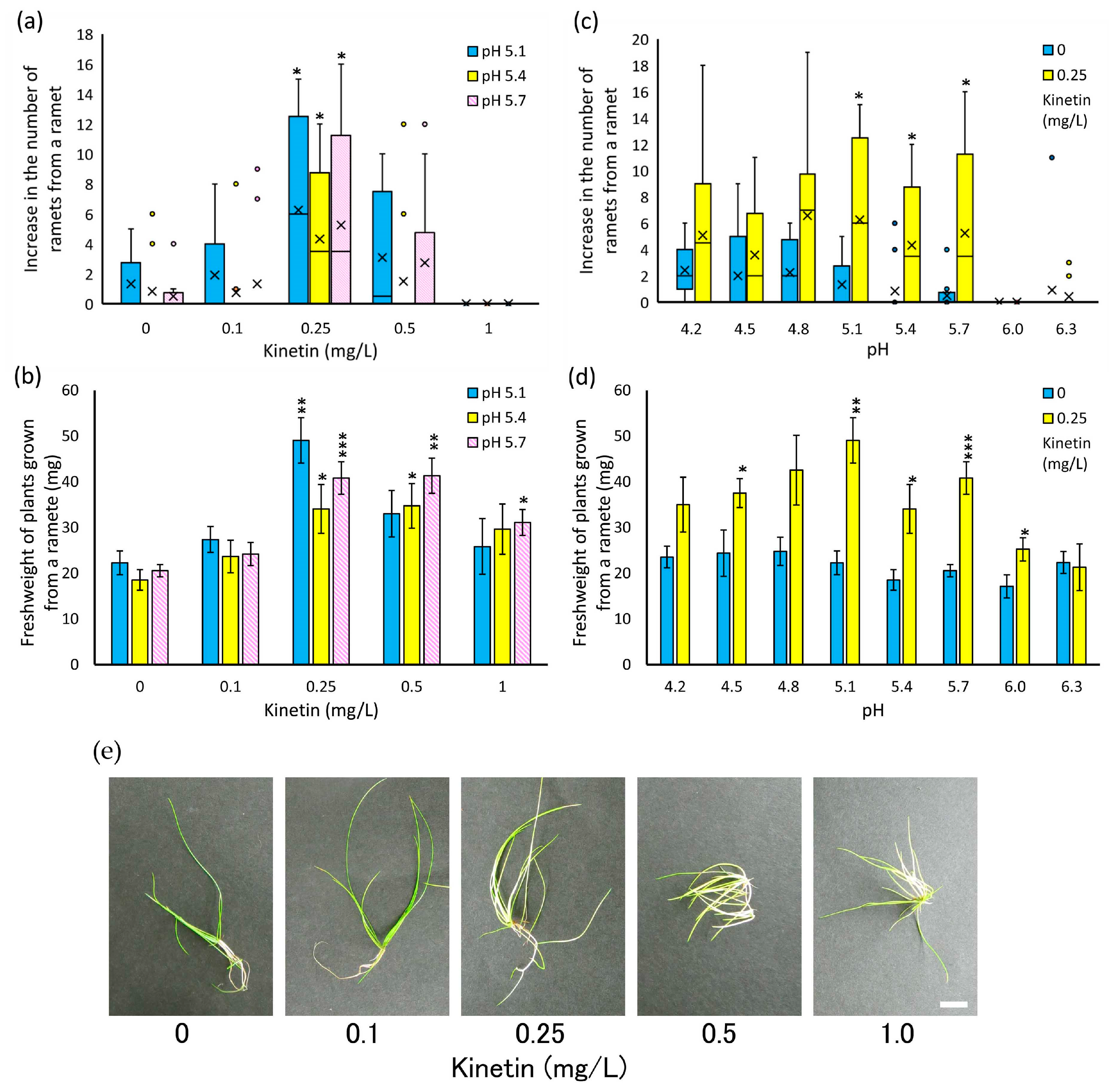
3.3. Analysis of Effective Growth Conditions for Individual Young Ramets
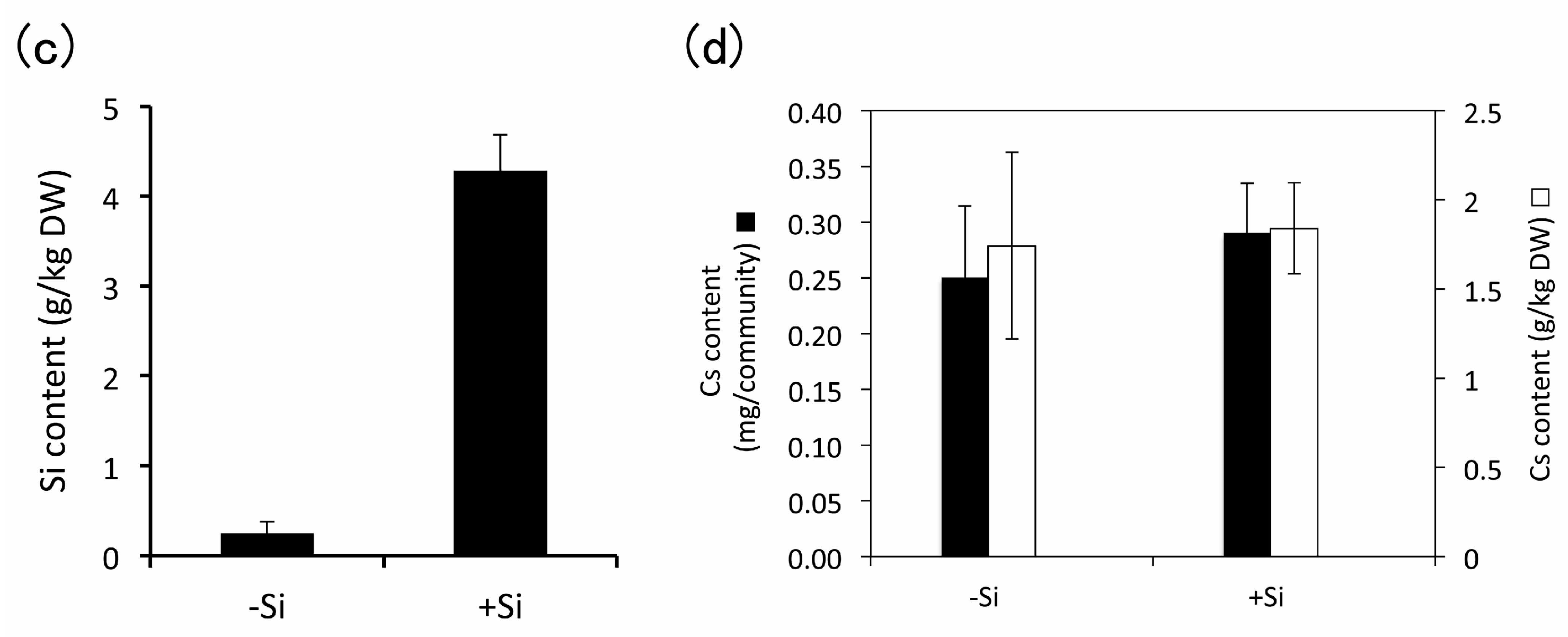
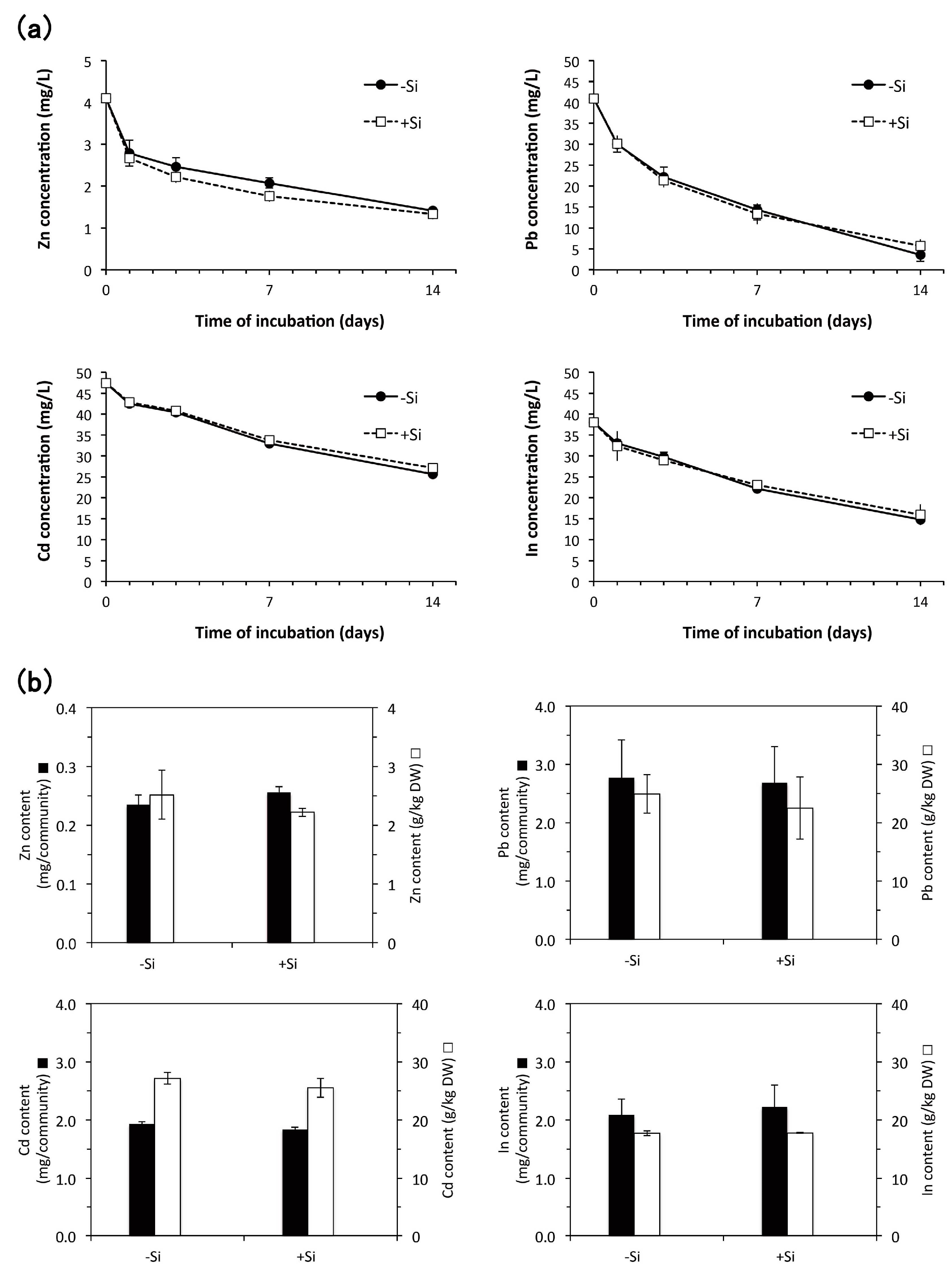
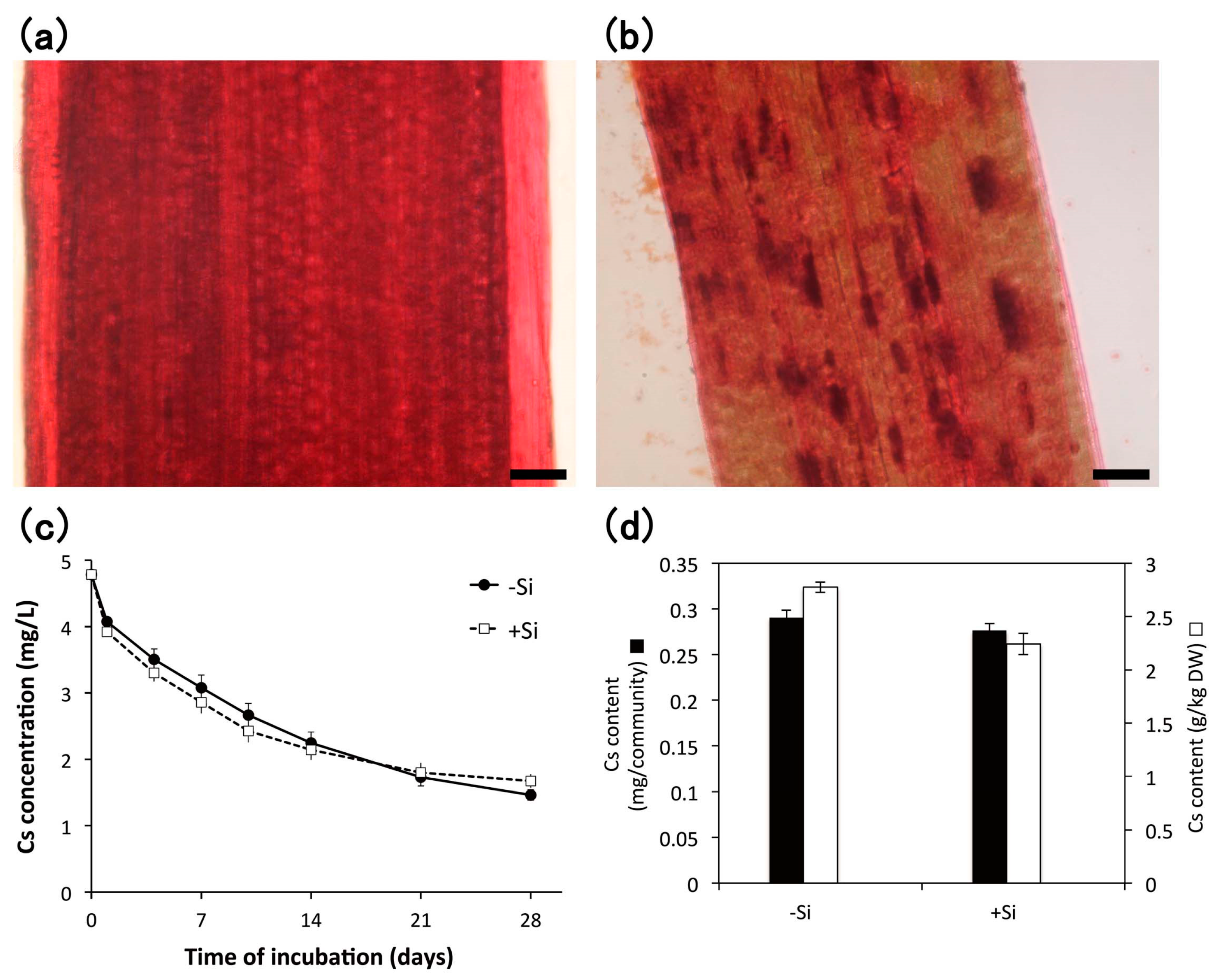
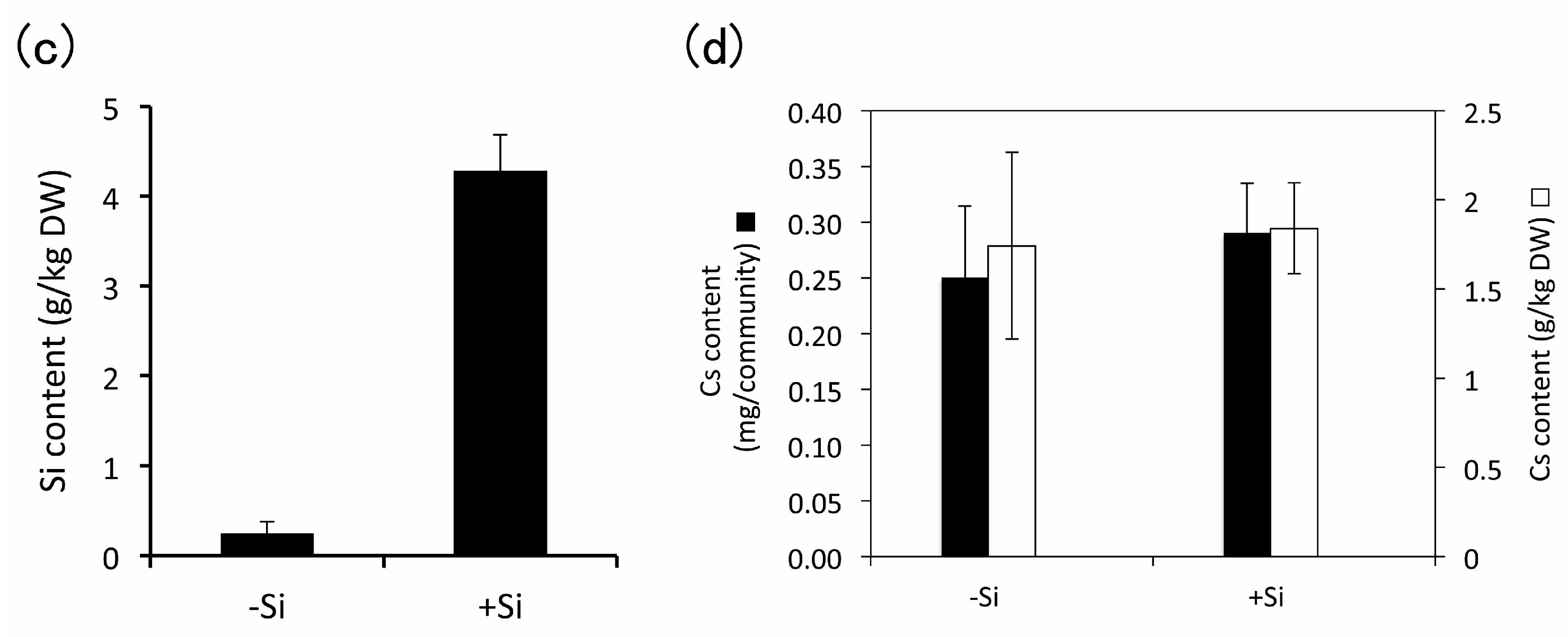
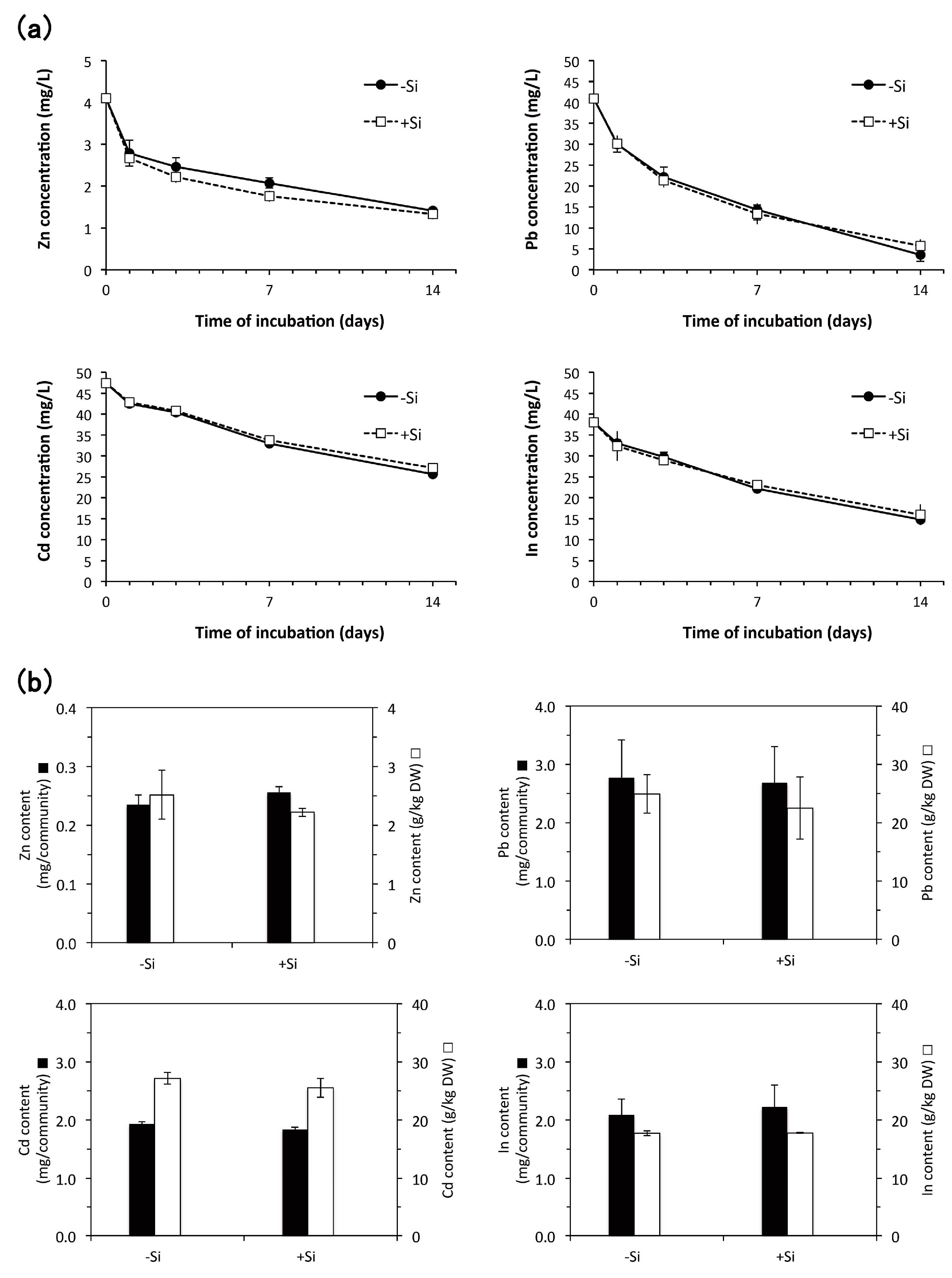
3.4. Heavy Metal Accumulation
4. Discussion
4.1. Primary Culture and Subculture Conditions
4.2. Growth Conditions for Young Ramets
4.3. Heavy Metal Accumulation
4.3.1. Cesium
4.3.2. Effect of Aseptic Culture of E. acicularis on Accumulation of Zn, Pb, Cd, and In
4.3.3. Effect of Silicon Addition during Subculture of E. acicularis on Heavy Metal Accumulation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.; Shahid, M.; Niazi, N.K.; Murartaza, B.; Bibi, I.; Dumat, C. A comparison of technologies for remediation of heavy metal contaminated soils. J. Geochem. Explor. 2016. [Google Scholar] [CrossRef]
- Gomes, M.A.; Hauser-Davis, R.A.; de Souza, A.N.; Vitória, A.P. Metal phytoremediation: General strategies, genetically modified plants and applications in metal nanoparticle contamination. Ecotoxicol. Environ. Saf. 2016, 134, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Ha, N.T.H.; Sakakibara, M.; Sano, S.; Hori, R.S.; Sera, K. The potential of Eleocharis acicularis for phytoremediation: Case study at an abandoned mine site. Clean 2009, 37, 203–208. [Google Scholar]
- Ha, N.T.H.; Sakakibara, M.; Sano, S. Phytoremediation of Sb, As, Cu, and Zn from contaminated water by the aquatic macrophyte Eleocharis acicularis. Clean 2009, 37, 720–725. [Google Scholar] [CrossRef]
- Ha, N.T.H.; Sakakibara, M.; Sano, S. Accumulation of Indium and other heavy metals by Eleocharis acicularis: An option for phytoremediation and phytomining. Bioresour. Technol. 2011, 102, 2228–2234. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, M.; Ohmori, Y.; Ha, N.T.H.; Sano, S.; Sera, K. Phytoremediation of heavy metal-contaminated water and sediment by Eleocharis acicularis. Clean 2011, 39, 735–741. [Google Scholar] [CrossRef]
- Sakakibara, M.; Sato, Y.; Sano, S.; Kubota, Y. Radioactive concentration of radioactive cesium of native Eleocharis acicularis in paddy fields of Fukushima Prefecture, northeastern Japan. J. Soc. Remediat. Radioact. Contam. Environ. 2014, 2, 13–18. [Google Scholar]
- Chekroun, K.B.; Baghour, M. The role of algae in phytoremediation of heavy metals: A review. J. Mater. Environ. Sci. 2013, 4, 873–880. [Google Scholar]
- Rezania, S.; Taib, S.M.; Din, M.F.M.; Dahalan, F.A.; Kamyab, H. Comprehensive review on phytotechnology: Heavy metals removal by diverse aquatic plants species from wastewater. J. Hazard. Mater. 2016, 318, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Dayanandan, P.; Kaufman, P.B.; Franklin, C.I. Detection of silica in plants. Am. J. Bot. 1983, 70, 1079–1084. [Google Scholar] [CrossRef]
- Sueoka, Y.; Sakakibara, M.; Sano, S.; Yamamoto, Y. A new method of environmental assessment and monitoring of Cu, Zn, As, and Pb pollution in surface soil using terricolous fruticose lichens. Environments 2016, 3, 35. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Tamai, K.; Mitani, N. Genotypic difference in silicon uptake and expression of silicon transporter genes in rice. Plant Physiol. 2007, 145, 919–924. [Google Scholar] [CrossRef] [PubMed]
- Okuda, A.; Takahashi, E. Studies on the physiological role of silicon in crop plants: 1. Discussion on the silicon deficient culture method. J. Sci. Soil Manure Jpn. 1961, 32, 475–480. [Google Scholar]
- Kara, Y.; Zeytunluoglu, A. Bioaccumulation of Toxic Metals (Cd and Cu) by Groenlandia densa (L.) Fourr. Bull. Environ. Contam. Toxicol. 2007, 79, 609–612. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Binth, L.T.; Muoi, L.T.; Oanh, H.T.K.; Thang, T.D.; Phong, D.T. Rapid propagation of agave by in vitro tissue culture. Plant Cell Tissue Organ Cult. 1990, 23, 67–70. [Google Scholar] [CrossRef]
- Kojoma, M.; Ohyama, K.; Seki, H.; Hiraoka, Y.; Asazu, S.A.; Sawa, S.; Sekizaki, H.; Yoshida, S.; Muranaka, T. In vitro proliferation and triterpenoid characteristics of licorice (Glycyrrhiza uralensis Fischer, Leguminosae) stolons. Plant Biotechnol. 2010, 27, 56–66. [Google Scholar] [CrossRef]
- Hedden, P.; Spontel, V. A century of gibberellin research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef] [PubMed]
- Saini, S.; Sharma, I.; Kaur, N.; Pati, P.K. Auxin: A master regulator in plant root development. Plant Cell Rep. 2013, 32, 741–757. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.H.; Lall, S.; Che, P. Cytokinins and shoot development. Trends Plant Sci. 2003, 8, 453–459. [Google Scholar] [CrossRef]
- Miller, C.O.; Skoog, F.; Saltza, M.H.V.; Strong, F.M. Kinetin, a cell division factor from deoxyribonucleic acid. J. Am. Chem. Soc. 1955, 77, 1392. [Google Scholar] [CrossRef]
- Takehara, A.; Nishiwaki, H.; Sato, Y.; Sakakibara, M. Roles of organic acids released by aquatic macrophyte Eleocharis acicularis. In Proceedings of the International Conference of Transdisciplinary Research on Environmental Problem in Southeastern Asia (TREPSEA), Makassar, Indonesia, 4–5 September 2014; pp. 85–88. [Google Scholar]
- Lottermoser, B.G.; Ashley, P.M. Trace element uptake by Eleocharis equisetina (spike rush) in an abandoned acid mine tailings pond, northeastern Australia: Implications for land and water reclamation in tropical regions. Environ. Pollut. 2011, 159, 3028–3035. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Lai, J.L.; Tao, Z.Y.; Han, N.; Wu, G. Characterizations of bio-accumulations, subcellular distribution and chemical forms of cesium in Brassica juncea, and Vicia faba. J. Environ. Radioact. 2016, 154, 52–59. [Google Scholar] [CrossRef] [PubMed]
- John, R.; Ahmad, P.; Gadgil, K.; Sharma, S. Heavy metal toxicity: Effect on plant growth, biochemical parameters and metal accumulation by Brassica juncea L. Int. J. Plant Prod. 2009, 3, 65–76. [Google Scholar]
- Baker, A.J.M.; Brooks, R.R. Terrestrial higher plants which hyper accumulate metallic elements—A review of their distribution, ecology, and phytochemistry. Biorecovery 1989, 1, 81–126. [Google Scholar]
- Kobayashi, D.; Uozumi, N.; Hisamatsu, S.; Yamagami, M. AtKUP/HAK/KT9, a K+ transporter from Arabidopsis thaliana, mediates Cs+ uptake in Escherichia coli. Biosci. Biotechnol. Biochem. 2010, 74, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Isaure, M.-P.; Fraysse, A.; Devès, G.; Le Lay, P.; Fayard, B.; Susini, J.; Bourguignon, J.; Ortega, R. Micro-chemical imaging of cesium distribution in Arabidopsis thaliana plant and its interaction with potassium and essential trace elements. Biochimie 2006, 88, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Seregin, I.V.; Kozhevnikova, A.D. Roles of root and shoot tissues in transport and accumulation of cadmium, lead, nickel, and strontium. Russ. J. Plant Physiol. 2008, 55, 1–22. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heavy Metal | Initial Conc. of Fundamental Solution (mg/L) | Si Conc. (mM) | Incubation Period (d) | Heavy Metal Removal (mg/L Solution) | DW of Whole Plants after Incubation (mg/L Solution) | Heavy Metal Accumulation in Whole Plants (mg/kg DW) | Accumulation Rate (%) (Accumulation in Whole Plants/Removal from Solution) | BCF |
|---|---|---|---|---|---|---|---|---|
| Cs | 4.79 ± 0.02 | 0 | 28 | 3.32 ± 0.10 | 1046 ± 25 | 2782 ± 48 | 87.6 ± 0.7 | 581 ± 10 |
| 0.1 | 3.11 ± 0.12 | 1234 ± 25 | 2248 ± 99 | 89.1 ± 1.1 | 470 ± 21 | |||
| Zn | 4.10 ± 0.04 | 0 | 14 | 2.69 ± 0.06 | 970 ± 228 | 2517 ± 414 | 87.0 ± 4.3 | 613 ± 101 |
| 1 | 2.77 ± 0.12 | 1152 ± 11 | 2219 ± 69 | 92.1 ± 0.2 | 541 ± 17 | |||
| Pb | 40.89 ± 0.29 | 0 | 14 | 37.35 ± 1.44 | 1097 ± 116 | 24885 ± 3293 | 73.5 ± 14.5 | 609 ± 81 |
| 1 | 35.21 ± 1.54 | 1196 ± 9 | 22486 ± 5320 | 75.6 ± 14.2 | 550 ± 130 | |||
| Cd | 47.40 ± 0.21 | 0 | 14 | 21.75 ± 0.24 | 713 ± 12 | 27153 ± 1017 | 88.9 ± 0.9 | 573 ± 21 |
| 1 | 20.25 ± 0.25 | 722 ± 30 | 25556 ± 1613 | 90.9 ± 3.1 | 539 ± 34 | |||
| In | 38.06 ± 0.75 | 0 | 14 | 23.25 ± 0.21 | 1180 ± 14 | 17737 ± 2519 | 90.0 ± 12.5 | 466 ± 66 |
| 1 | 22.11 ± 2.43 | 1244 ± 79 | 17805 ± 1865 | 100.1 ± 5.8 | 468 ± 49 |
| Heavy Metal | Cs | Zn | Pb | Cd | In |
|---|---|---|---|---|---|
| Content (mg/kg DW) | 6.12 ± 0.60 | 148.5 ± 0.17 | 0.36 ± 0.16 | 0.15 ± 0.09 | 0.02 ± 0.00 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, Y.; Goto, S.; Teraoka, S.; Takagaki, K.; Takehara, A.; Sano, S.; Sakakibara, M. Establishment of an Aseptic Culture System and Analysis of the Effective Growth Conditions for Eleocharis acicularis Ramets for Use in Phytoremediation. Environments 2017, 4, 40. https://doi.org/10.3390/environments4020040
Sato Y, Goto S, Teraoka S, Takagaki K, Takehara A, Sano S, Sakakibara M. Establishment of an Aseptic Culture System and Analysis of the Effective Growth Conditions for Eleocharis acicularis Ramets for Use in Phytoremediation. Environments. 2017; 4(2):40. https://doi.org/10.3390/environments4020040
Chicago/Turabian StyleSato, Yasushi, Shinpei Goto, Shoya Teraoka, Katsuya Takagaki, Akinari Takehara, Sakae Sano, and Masayuki Sakakibara. 2017. "Establishment of an Aseptic Culture System and Analysis of the Effective Growth Conditions for Eleocharis acicularis Ramets for Use in Phytoremediation" Environments 4, no. 2: 40. https://doi.org/10.3390/environments4020040
APA StyleSato, Y., Goto, S., Teraoka, S., Takagaki, K., Takehara, A., Sano, S., & Sakakibara, M. (2017). Establishment of an Aseptic Culture System and Analysis of the Effective Growth Conditions for Eleocharis acicularis Ramets for Use in Phytoremediation. Environments, 4(2), 40. https://doi.org/10.3390/environments4020040