
Mindfulness Affects the Boundaries of Bodily Self-Representation: The Effect of Focused-Attention Meditation in Fading the Boundary of Peripersonal Space
 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Prospective Power Analysis
2.2. Participants
2.3. Apparatus and Stimuli
2.4. Design and Procedure
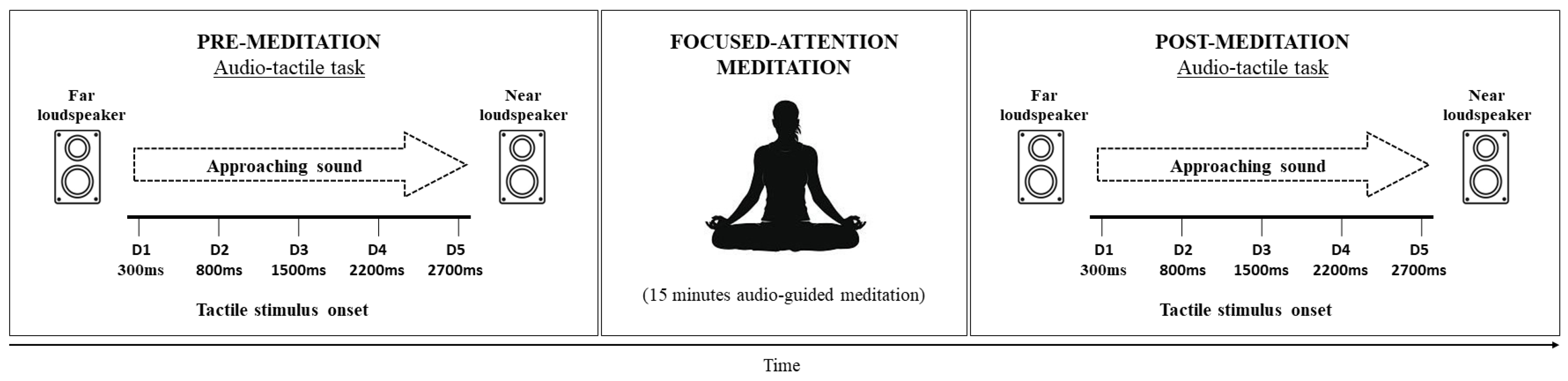
2.4.1. The Audio–Tactile Task
2.4.2. Focused-Attention Meditation Session
2.5. Data Analysis
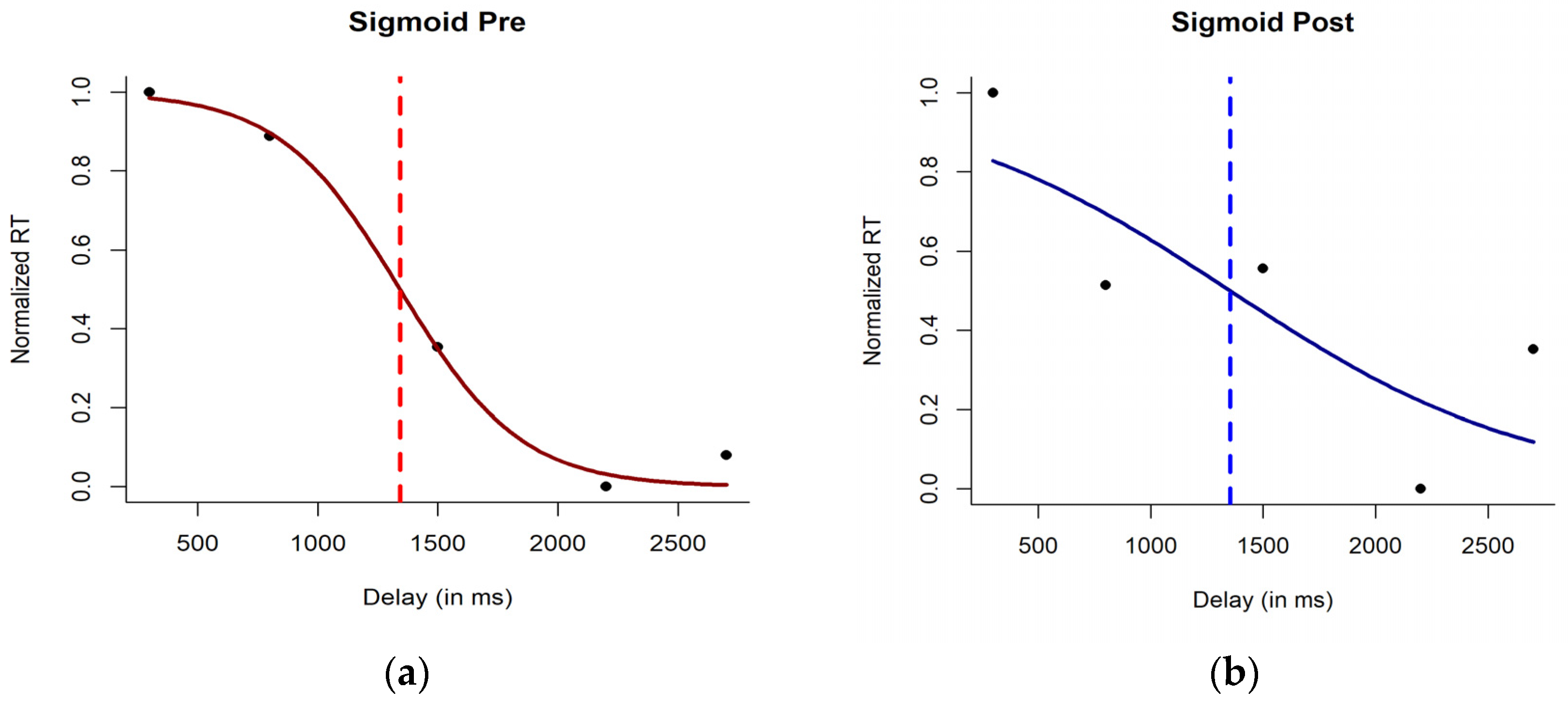
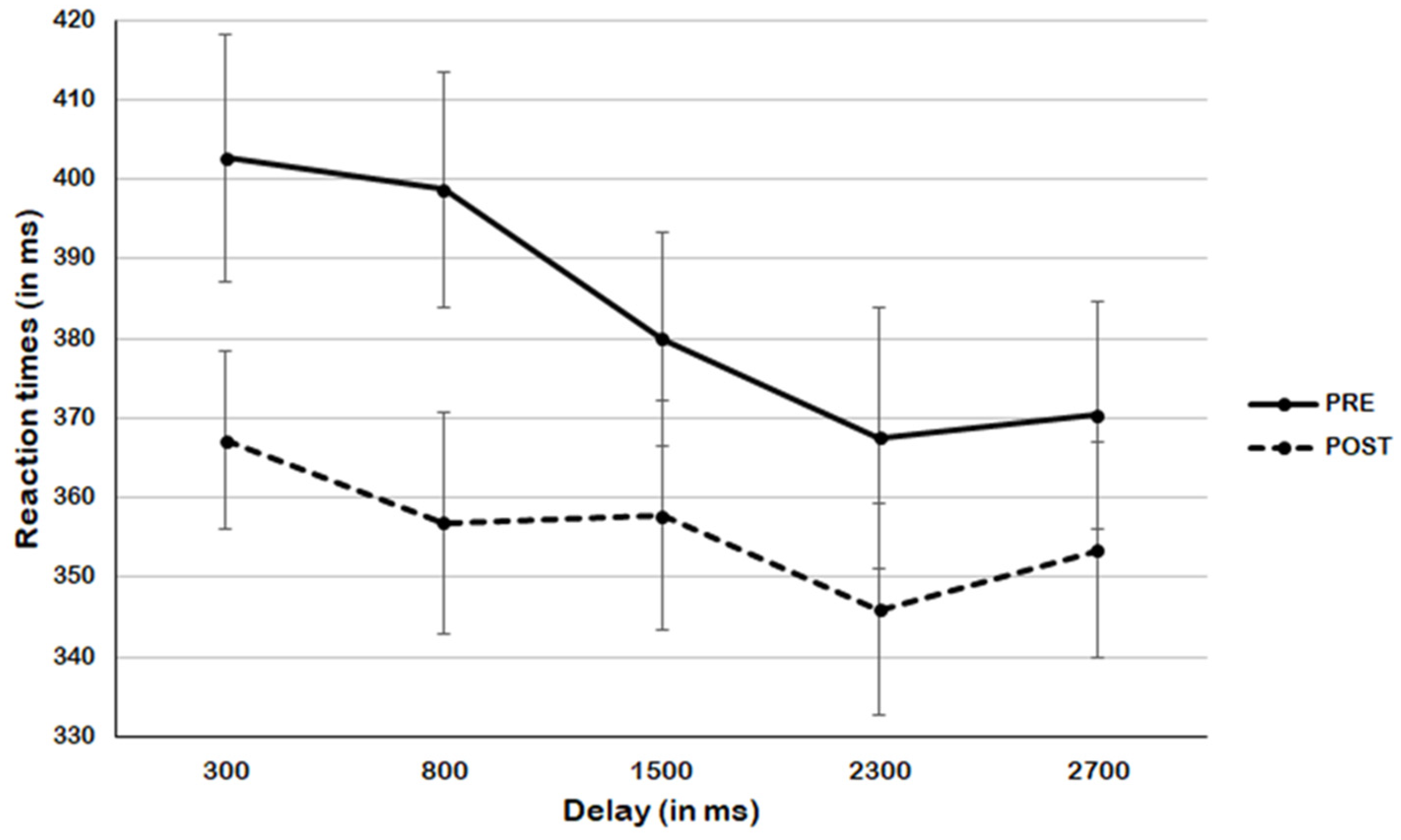
3. Results
4. Discussion
5. Limitation of the Study and Future Directions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Transcription of the Audio-Guided Focused-Attention Meditation Practice
References
- Gallagher, S. Philosophical Conceptions of the Self: Implications for Cognitive Science. Trends Cogn. Sci. 2000, 4, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Damasio, A.R. How the Brain Creates the Mind. Sci. Am. 1999, 281, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Blanke, O.; Slater, M.; Serino, A. Behavioral, Neural, and Computational Principles of Bodily Self-Consciousness. Neuron 2015, 88, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Serino, A. Peripersonal Space (PPS) as a Multisensory Interface between the Individual and the Environment, Defining the Space of the Self. Neurosci. Biobehav. Rev. 2019, 99, 138–159. [Google Scholar] [CrossRef] [PubMed]
- di Pellegrino, G.; Làdavas, E. Peripersonal Space in the Brain. Neuropsychologia 2015, 66, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Iriki, A.; Tanaka, M.; Iwamura, Y. Coding of Modified Body Schema during Tool Use by Macaque Postcentral Neurones. NeuroReport 1996, 7, 2325–2330. [Google Scholar] [CrossRef] [PubMed]
- Farnè, A.; Làdavas, E. Dynamic Size-Change of Hand Peripersonal Space Following Tool Use. NeuroReport 2000, 11, 1645. [Google Scholar] [CrossRef]
- Teneggi, C.; Canzoneri, E.; di Pellegrino, G.; Serino, A. Social Modulation of Peripersonal Space Boundaries. Curr. Biol. 2013, 23, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Holmes, N.P.; Spence, C. The Body Schema and Multisensory Representation(s) of Peripersonal Space. Cogn. Process. 2004, 5, 94–105. [Google Scholar] [CrossRef]
- Blanke, O. Multisensory Brain Mechanisms of Bodily Self-Consciousness. Nat. Rev. Neurosci. 2012, 13, 556–571. [Google Scholar] [CrossRef]
- Blanke, O.; Metzinger, T. Full-Body Illusions and Minimal Phenomenal Selfhood. Trends Cogn. Sci. 2009, 13, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Tsakiris, M. The Multisensory Basis of the Self: From Body to Identity to Others. Q. J. Exp. Psychol. 2017, 70, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D.M. Spatial Limits on Referred Touch to an Alien Limb May Reflect Boundaries of Visuo-Tactile Peripersonal Space Surrounding the Hand. Brain Cogn. 2007, 64, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Samad, M.; Chung, A.J.; Shams, L. Perception of Body Ownership Is Driven by Bayesian Sensory Inference. PLoS ONE 2015, 10, e0117178. [Google Scholar] [CrossRef] [PubMed]
- Noel, J.-P.; Pfeiffer, C.; Blanke, O.; Serino, A. Peripersonal Space as the Space of the Bodily Self. Cognition 2015, 144, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Kabat-Zinn, J. Mindfulness-Based Interventions in Context: Past, Present, and Future. Clin. Psychol. Sci. Pract. 2003, 10, 144–156. [Google Scholar] [CrossRef]
- Birnie, K.; Speca, M.; Carlson, L.E. Exploring Self-Compassion and Empathy in the Context of Mindfulness-Based Stress Reduction (MBSR). Stress Health 2010, 26, 359–371. [Google Scholar] [CrossRef]
- Hutcherson, C.; Seppala, E.; Gross, J. Loving-Kindness Meditation Increases Social Connectedness. Emotion 2008, 8, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Berkovich-Ohana, A.; Dor-Ziderman, Y.; Glicksohn, J.; Goldstein, A. Alterations in the Sense of Time, Space, and Body in the Mindfulness-Trained Brain: A Neurophenomenologically-Guided MEG Study. Front. Psychol. 2013, 4, 912. [Google Scholar] [CrossRef]
- Dor-Ziderman, Y.; Ataria, Y.; Fulder, S.; Goldstein, A.; Berkovich-Ohana, A. Self-Specific Processing in the Meditating Brain: A MEG Neurophenomenology Study. Neurosci. Conscious. 2016, 2016, niw019. [Google Scholar] [CrossRef]
- Farb, N.A.S.; Segal, Z.V.; Mayberg, H.; Bean, J.; McKeon, D.; Fatima, Z.; Anderson, A.K. Attending to the Present: Mindfulness Meditation Reveals Distinct Neural Modes of Self-Reference. Soc. Cogn. Affect. Neurosci. 2007, 2, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Desbordes, G. Self-Related Processing in Mindfulness-Based Interventions. Curr. Opin. Psychol. 2019, 28, 312–316. [Google Scholar] [CrossRef]
- Chiarella, S.G.; Makwana, M.; Simione, L.; Hartkamp, M.; Calabrese, L.; Raffone, A.; Srinivasan, N. Mindfulness Meditation Weakens Attachment to Self: Evidence from a Self vs. Other Binding Task. Mindfulness 2020, 11, 2411–2422. [Google Scholar] [CrossRef]
- Lindahl, J.R.; Britton, W.B. “I Have This Feeling of Not Really Being Here”: Buddhist Meditation and Changes in Sense of Self. J. Conscious. Stud. 2019, 26, 157–183. [Google Scholar]
- Logie, K.; Frewen, P. Self/Other Referential Processing Following Mindfulness and Loving-Kindness Meditation. Mindfulness 2015, 6, 778–787. [Google Scholar] [CrossRef]
- Shi, Z.; He, L. Mindfulness: Attenuating Self-Referential Processing and Strengthening Other-Referential Processing. Mindfulness 2020, 11, 599–605. [Google Scholar] [CrossRef]
- Trautwein, F.-M.; Naranjo, J.R.; Schmidt, S. Meditation Effects in the Social Domain: Self-Other Connectedness as a General Mechanism? In Meditation—Neuroscientific Approaches and Philosophical Implications; Schmidt, S., Walach, H., Eds.; Studies in Neuroscience, Consciousness and Spirituality; Springer International Publishing: Cham, Switzerland, 2014; pp. 175–198. ISBN 978-3-319-01634-4. [Google Scholar]
- Trautwein, F.-M.; Naranjo, J.R.; Schmidt, S. Decentering the Self? Preliminary Evidence for Changes in Self- vs. Other Related Processing as a Long-Term Outcome of Loving-Kindness Meditation. Front. Psychol. 2016, 7, 1785. [Google Scholar] [CrossRef]
- Besse, R.; Whitaker, W.K.; Brannon, L.A. Reducing Loneliness: The Impact of Mindfulness, Social Cognitions, and Coping. Psychol. Rep. 2022, 125, 1289–1304. [Google Scholar] [CrossRef] [PubMed]
- Teoh, S.L.; Letchumanan, V.; Lee, L.-H. Can Mindfulness Help to Alleviate Loneliness? A Systematic Review and Meta-Analysis. Front. Psychol. 2021, 12, 633319. [Google Scholar] [CrossRef]
- Lindsay, E.K.; Young, S.; Brown, K.W.; Smyth, J.M.; Creswell, J.D. Mindfulness Training Reduces Loneliness and Increases Social Contact in a Randomized Controlled Trial. Proc. Natl. Acad. Sci. USA 2019, 116, 3488–3493. [Google Scholar] [CrossRef]
- Ataria, Y.; Dor-Ziderman, Y.; Berkovich-Ohana, A. How Does It Feel to Lack a Sense of Boundaries? A Case Study of a Long-Term Mindfulness Meditator. Conscious. Cogn. 2015, 37, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.H.H.; Palmer, S.B.; Aday, J.S.; Davoli, C.C.; Bloesch, E.K. Meditation Alters Representations of Peripersonal Space: Evidence from Auditory Evoked Potentials. Conscious. Cogn. 2020, 83, 102978. [Google Scholar] [CrossRef] [PubMed]
- Reed, C.; Leland, D.; Brekke, B.; Hartley, A. Attention’s Grasp: Early and Late Hand Proximity Effects on Visual Evoked Potentials. Front. Psychol. 2013, 4, 420. [Google Scholar] [CrossRef] [PubMed]
- Ardizzi, M.; Ferri, F. Interoceptive Influences on Peripersonal Space Boundary. Cognition 2018, 177, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Khalsa, S.S.; Rudrauf, D.; Hassanpour, M.S.; Davidson, R.J.; Tranel, D. The Practice of Meditation Is Not Associated with Improved Interoceptive Awareness of the Heartbeat. Psychophysiology 2020, 57, e13479. [Google Scholar] [CrossRef] [PubMed]
- Yordanova, J.; Kolev, V.; Nicolardi, V.; Simione, L.; Mauro, F.; Garberi, P.; Raffone, A.; Malinowski, P. Attentional and Cognitive Monitoring Brain Networks in Long-Term Meditators Depend on Meditation States and Expertise. Sci. Rep. 2021, 11, 4909. [Google Scholar] [CrossRef] [PubMed]
- Yordanova, J.; Kolev, V.; Mauro, F.; Nicolardi, V.; Simione, L.; Calabrese, L.; Malinowski, P.; Raffone, A. Common and Distinct Lateralised Patterns of Neural Coupling during Focused Attention, Open Monitoring and Loving Kindness Meditation. Sci. Rep. 2020, 10, 7430. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J. Mindfulness, Interoception, and the Body: A Contemporary Perspective. Front. Psychol. 2019, 10, 2012. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J.E. The Embodied Mind: An Exploratory Study on the Subjective, Developmental, and Outcome Effects of an Intervention Designed to Increase Interoceptive Awareness. Ph.D. Thesis, University of West Georgia, Carrollton, GA, USA, 2014. [Google Scholar]
- Hasenkamp, W.; Wilson-Mendenhall, C.D.; Duncan, E.; Barsalou, L.W. Mind Wandering and Attention during Focused Meditation: A Fine-Grained Temporal Analysis of Fluctuating Cognitive States. NeuroImage 2012, 59, 750–760. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Scandolara, C.; Matelli, M.; Gentilucci, M. Afferent Properties of Periarcuate Neurons in Macaque Monkeys. II. Visual Responses. Behav. Brain Res. 1981, 2, 147–163. [Google Scholar] [CrossRef]
- Bogdanova, O.V.; Bogdanov, V.B.; Dureux, A.; Farnè, A.; Hadj-Bouziane, F. The Peripersonal Space in a Social World. Cortex 2021, 142, 28–46. [Google Scholar] [CrossRef] [PubMed]
- Maravita, A.; Spence, C.; Driver, J. Multisensory Integration and the Body Schema: Close to Hand and within Reach. Curr. Biol. 2003, 13, R531–R539. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.A.; Moore, D.R. Auditory Neuroscience: The Salience of Looming Sounds. Curr. Biol. 2003, 13, R91–R93. [Google Scholar] [CrossRef] [PubMed]
- Cardini, F.; Fatemi-Ghomi, N.; Gajewska-Knapik, K.; Gooch, V.; Aspell, J.E. Enlarged Representation of Peripersonal Space in Pregnancy. Sci. Rep. 2019, 9, 8606. [Google Scholar] [CrossRef] [PubMed]
- Ferroni, F.; Ardizzi, M.; Magnani, F.; Ferri, F.; Langiulli, N.; Rastelli, F.; Lucarini, V.; Giustozzi, F.; Volpe, R.; Marchesi, C.; et al. Tool-Use Extends Peripersonal Space Boundaries in Schizophrenic Patients. Schizophr. Bull. 2022, 48, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Hong, S.-J.J.; Baxter, T.; Scott, J.; Shenoy, S.; Buck, L.; Bodenheimer, B.; Park, S. Altered Peripersonal Space and the Bodily Self in Schizophrenia: A Virtual Reality Study. Schizophr. Bull. 2021, 47, 927–937. [Google Scholar] [CrossRef]
- Noel, J.-P.; Park, H.-D.; Pasqualini, I.; Lissek, H.; Wallace, M.; Blanke, O.; Serino, A. Audio-Visual Sensory Deprivation Degrades Visuo-Tactile Peri-Personal Space. Conscious. Cogn. 2018, 61, 61–75. [Google Scholar] [CrossRef]
- Canzoneri, E.; Magosso, E.; Serino, A. Dynamic Sounds Capture the Boundaries of Peripersonal Space Representation in Humans. PLoS ONE 2012, 7, e44306. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Serino, A.; Bassolino, M.; Farnè, A.; Làdavas, E. Extended Multisensory Space in Blind Cane Users. Psychol. Sci. 2007, 18, 642–648. [Google Scholar] [CrossRef]
- Maister, L.; Cardini, F.; Zamariola, G.; Serino, A.; Tsakiris, M. Your Place or Mine: Shared Sensory Experiences Elicit a Remapping of Peripersonal Space. Neuropsychologia 2015, 70, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Todd, J.; Aspell, J.E. Mindfulness, Interoception, and the Body. Brain Sci. 2022, 12, 696. [Google Scholar] [CrossRef] [PubMed]
- Làdavas, E.; Serino, A. Action-Dependent Plasticity in Peripersonal Space Representations. Cogn. Neuropsychol. 2008, 25, 1099–1113. [Google Scholar] [CrossRef] [PubMed]
- Martel, M.; Cardinali, L.; Roy, A.C.; Farnè, A. Tool-Use: An Open Window into Body Representation and Its Plasticity. Cogn. Neuropsychol. 2016, 33, 82–101. [Google Scholar] [CrossRef] [PubMed]
- McCourt, M.E.; Garlinghouse, M. Asymmetries of Visuospatial Attention Are Modulated by Viewing Distance and Visual Field Elevation: Pseudoneglect in Peripersonal and Extrapersonal Space. Cortex 2000, 36, 715–731. [Google Scholar] [CrossRef] [PubMed]
- Làdavas, E.; di Pellegrino, G.; Farnè, A.; Zeloni, G. Neuropsychological Evidence of an Integrated Visuotactile Representation of Peripersonal Space in Humans. J. Cogn. Neurosci. 1998, 10, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Farnè, A.; Làdavas, E. Auditory Peripersonal Space in Humans. J. Cogn. Neurosci. 2002, 14, 1030–1043. [Google Scholar] [CrossRef] [PubMed]
- Sambo, C.F.; Forster, B. An ERP Investigation on Visuotactile Interactions in Peripersonal and Extrapersonal Space: Evidence for the Spatial Rule. J. Cogn. Neurosci. 2009, 21, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Sambo, C.F.; Forster, B.; Williams, S.C.; Iannetti, G.D. To Blink or Not to Blink: Fine Cognitive Tuning of the Defensive Peripersonal Space. J. Neurosci. 2012, 32, 12921–12927. [Google Scholar] [CrossRef]
- De Paepe, A.L.; Crombez, G.; Spence, C.; Legrain, V. Mapping Nociceptive Stimuli in a Peripersonal Frame of Reference: Evidence from a Temporal Order Judgment Task. Neuropsychologia 2014, 56, 219–228. [Google Scholar] [CrossRef]
- Bufacchi, R.J.; Iannetti, G.D. An Action Field Theory of Peripersonal Space. Trends Cogn. Sci. 2018, 22, 1076–1090. [Google Scholar] [CrossRef] [PubMed]
- Colby, C.L.; Duhamel, J.R.; Goldberg, M.E. Ventral Intraparietal Area of the Macaque: Anatomic Location and Visual Response Properties. J. Neurophysiol. 1993, 69, 902–914. [Google Scholar] [CrossRef] [PubMed]
- Sedda, A.; Ambrosini, E.; Dirupo, G.; Tonin, D.; Valsecchi, L.; Redaelli, T.; Spinelli, M.; Costantini, M.; Bottini, G. Affordances after Spinal Cord Injury. J. Neuropsychol. 2019, 13, 354–369. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.R.; Lourenco, S.F. On the Nature of near Space: Effects of Tool Use and the Transition to Far Space. Neuropsychologia 2006, 44, 977–981. [Google Scholar] [CrossRef] [PubMed]
- Graziano, M.S.A.; Cooke, D.F. Parieto-Frontal Interactions, Personal Space, and Defensive Behavior. Neuropsychologia 2006, 44, 2621–2635. [Google Scholar] [CrossRef] [PubMed]
- Hediger, H. Studies of the Psychology and Behavior of Captive Animals in Zoos and Circuses; Criterion Books, Inc.: Oxford, UK, 1955. [Google Scholar]
- Cooke, D.F.; Graziano, M.S.A. Sensorimotor Integration in the Precentral Gyrus: Polysensory Neurons and Defensive Movements. J. Neurophysiol. 2004, 91, 1648–1660. [Google Scholar] [CrossRef] [PubMed]
- Cooke, D.F.; Taylor, C.S.R.; Moore, T.; Graziano, M.S.A. Complex Movements Evoked by Microstimulation of the Ventral Intraparietal Area. Proc. Natl. Acad. Sci. USA 2003, 100, 6163–6168. [Google Scholar] [CrossRef]
- Graziano, M.S.A.; Taylor, C.S.R.; Moore, T. Probing Cortical Function with Electrical Stimulation. Nat. Neurosci. 2002, 5, 921. [Google Scholar] [CrossRef]
- Taffou, M.; Viaud-Delmon, I. Cynophobic Fear Adaptively Extends Peri-Personal Space. Front. Psychiatry 2014, 5, 122. [Google Scholar] [CrossRef]
- Lourenco, S.F.; Longo, M.R.; Pathman, T. Near Space and Its Relation to Claustrophobic Fear. Cognition 2011, 119, 448–453. [Google Scholar] [CrossRef]
- Luberto, C.M.; Shinday, N.; Song, R.; Philpotts, L.L.; Park, E.R.; Fricchione, G.L.; Yeh, G.Y. A Systematic Review and Meta-Analysis of the Effects of Meditation on Empathy, Compassion, and Prosocial Behaviors. Mindfulness 2018, 9, 708–724. [Google Scholar] [CrossRef] [PubMed]
- Sforza, A.; Bufalari, I.; Haggard, P.; Aglioti, S.M. My Face in Yours: Visuo-Tactile Facial Stimulation Influences Sense of Identity. Soc. Neurosci. 2010, 5, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-Y.; Hölzel, B.K.; Posner, M.I. The Neuroscience of Mindfulness Meditation. Nat. Rev. Neurosci. 2015, 16, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, P.; Calabrese, L.; Chiarella, S.G.; D’Errico, G.; De Paolis, L.T.; Grassini, S.; Mastrati, G.; Moccaldi, N.; Raffone, A.; Vallefuoco, E. Mindfulness-Based Emotional Acceptance in Combination with Neurofeedback for Improving Emotion Self-Regulation: A Pilot Study. In Proceedings of the 2022 IEEE International Conference on Metrology for Extended Reality, Artificial Intelligence and Neural Engineering (MetroXRAINE), Rome, Italy, 26–28 October 2022; pp. 465–470. [Google Scholar]
- Accoto, A.; Chiarella, S.G.; Raffone, A.; Montano, A.; de Marco, A.; Mainiero, F.; Rubbino, R.; Valzania, A.; Conversi, D. Beneficial Effects of Mindfulness-Based Stress Reduction Training on the Well-Being of a Female Sample during the First Total Lockdown Due to COVID-19 Pandemic in Italy. Int. J. Environ. Res. Public Health 2021, 18, 5512. [Google Scholar] [CrossRef] [PubMed]
- Colombo, S.L.; Chiarella, S.G.; Raffone, A.; Simione, L. Understanding the Environmental Attitude-Behaviour Gap: The Moderating Role of Dispositional Mindfulness. Sustainability 2023, 15, 7285. [Google Scholar] [CrossRef]
- Hanley, A.W.; Derringer, S.A.; Hanley, R.T. Dispositional Mindfulness May Be Associated with Deeper Connections with Nature. Ecopsychology 2017, 9, 225–231. [Google Scholar] [CrossRef]
- Hanley, A.W.; Bettmann, J.E.; Kendrick, C.E.; Deringer, A.; Norton, C.L. Dispositional Mindfulness Is Associated with Greater Nature Connectedness and Self-Reported Ecological Behavior. Ecopsychology 2020, 12, 54–63. [Google Scholar] [CrossRef]
- Sanyer, M.M.; Bettmann, J.E.; Anstadt, G.; Ganesh, K.; Hanley, A.W. Decenter to Reenter Nature: Relationships between Decentering, Self-Transcendence, and Nature Connectedness. Psychol. Conscious. Theory Res. Pract. 2023, 10, 205–212. [Google Scholar] [CrossRef]
- Macaulay, R.; Lee, K.; Johnson, K.; Williams, K. Mindful Engagement, Psychological Restoration, and Connection with Nature in Constrained Nature Experiences. Landsc. Urban Plan. 2022, 217, 104263. [Google Scholar] [CrossRef]
- Graziano, M.S.A.; Gross, C.G. The Representation of Extrapersonal Space: A Possible Role for Bimodal, Visual-Tactile Neurons. In The Cognitive Neurosciences; The MIT Press: Cambridge, MA, USA, 1995; pp. 1021–1034. ISBN 978-0-262-07157-4. [Google Scholar]
- Fogassi, L.; Gallese, V.; Fadiga, L.; Luppino, G.; Matelli, M.; Rizzolatti, G. Coding of Peripersonal Space in Inferior Premotor Cortex (Area F4). J. Neurophysiol. 1996, 76, 141–157. [Google Scholar] [CrossRef]
- Schicke, T.; Bauer, F.; Röder, B. Interactions of Different Body Parts in Peripersonal Space: How Vision of the Foot Influences Tactile Perception at the Hand. Exp. Brain Res. 2009, 192, 703–715. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiarella, S.G.; De Pastina, R.; Raffone, A.; Simione, L. Mindfulness Affects the Boundaries of Bodily Self-Representation: The Effect of Focused-Attention Meditation in Fading the Boundary of Peripersonal Space. Behav. Sci. 2024, 14, 306. https://doi.org/10.3390/bs14040306
Chiarella SG, De Pastina R, Raffone A, Simione L. Mindfulness Affects the Boundaries of Bodily Self-Representation: The Effect of Focused-Attention Meditation in Fading the Boundary of Peripersonal Space. Behavioral Sciences. 2024; 14(4):306. https://doi.org/10.3390/bs14040306
Chicago/Turabian StyleChiarella, Salvatore Gaetano, Riccardo De Pastina, Antonino Raffone, and Luca Simione. 2024. "Mindfulness Affects the Boundaries of Bodily Self-Representation: The Effect of Focused-Attention Meditation in Fading the Boundary of Peripersonal Space" Behavioral Sciences 14, no. 4: 306. https://doi.org/10.3390/bs14040306
APA StyleChiarella, S. G., De Pastina, R., Raffone, A., & Simione, L. (2024). Mindfulness Affects the Boundaries of Bodily Self-Representation: The Effect of Focused-Attention Meditation in Fading the Boundary of Peripersonal Space. Behavioral Sciences, 14(4), 306. https://doi.org/10.3390/bs14040306