
Empathy Modulates the Activity of the Sensorimotor Mirror Neuron System during Pain Observation

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Subjects
2.2. Data Acquisition and Processing
2.3. Study Design
2.4. Simple T Contrasts
2.5. Region of Interest Analysis
2.6. Resting-State Analysis: Functional Connectivity
3. Results
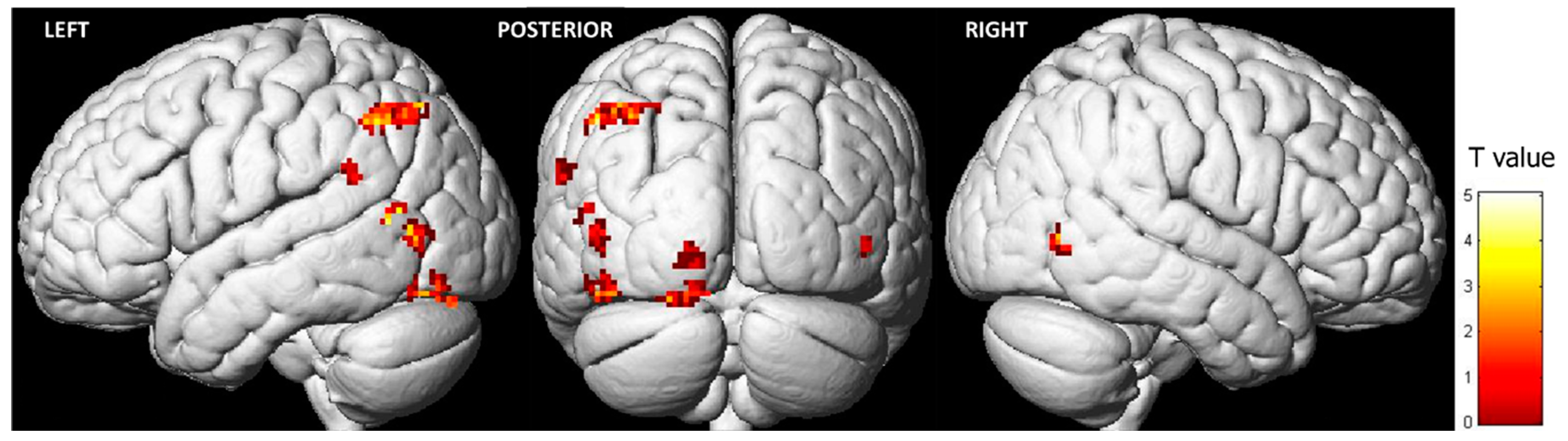
3.1. Brain Activity during the Observation of Pain Expressions
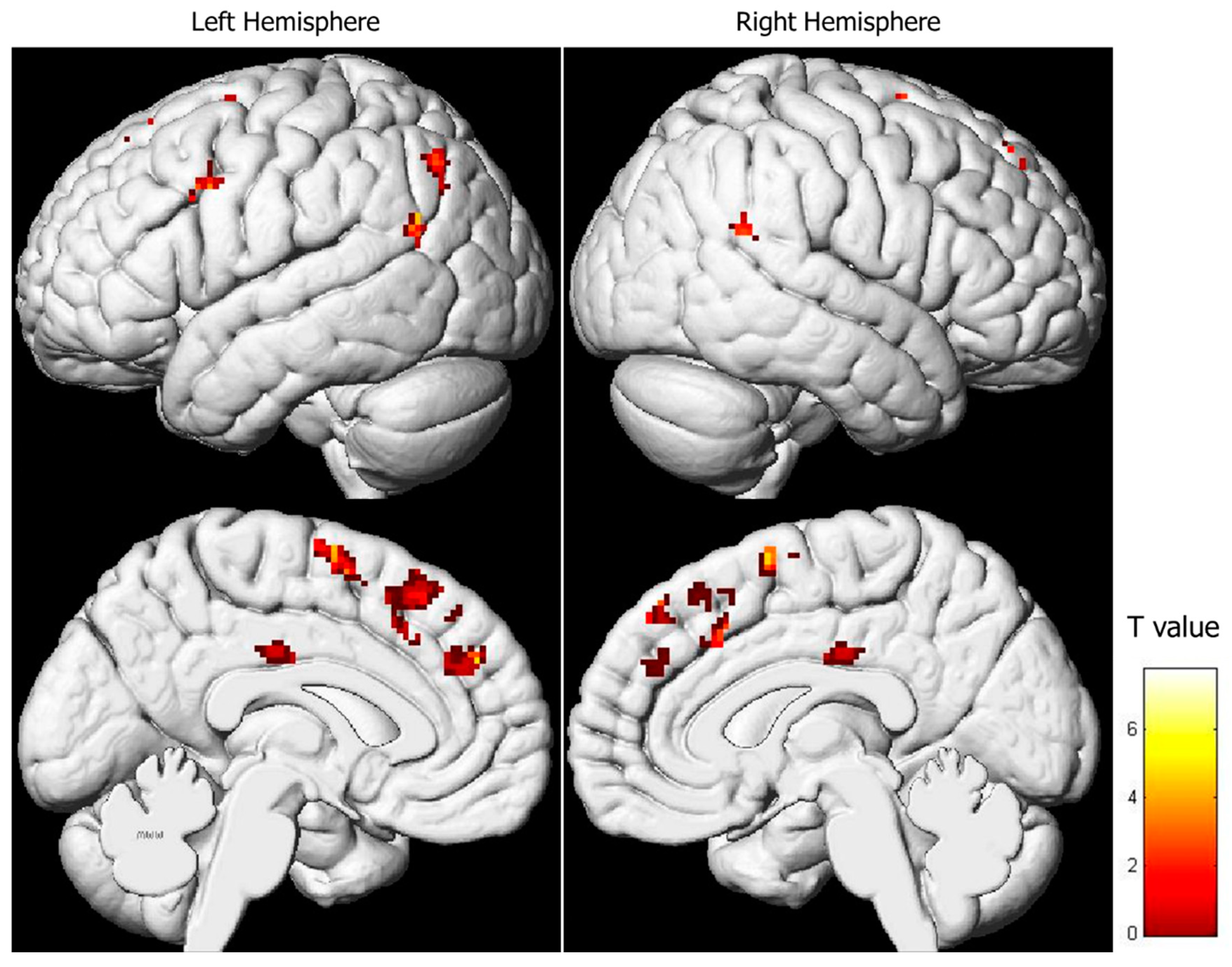
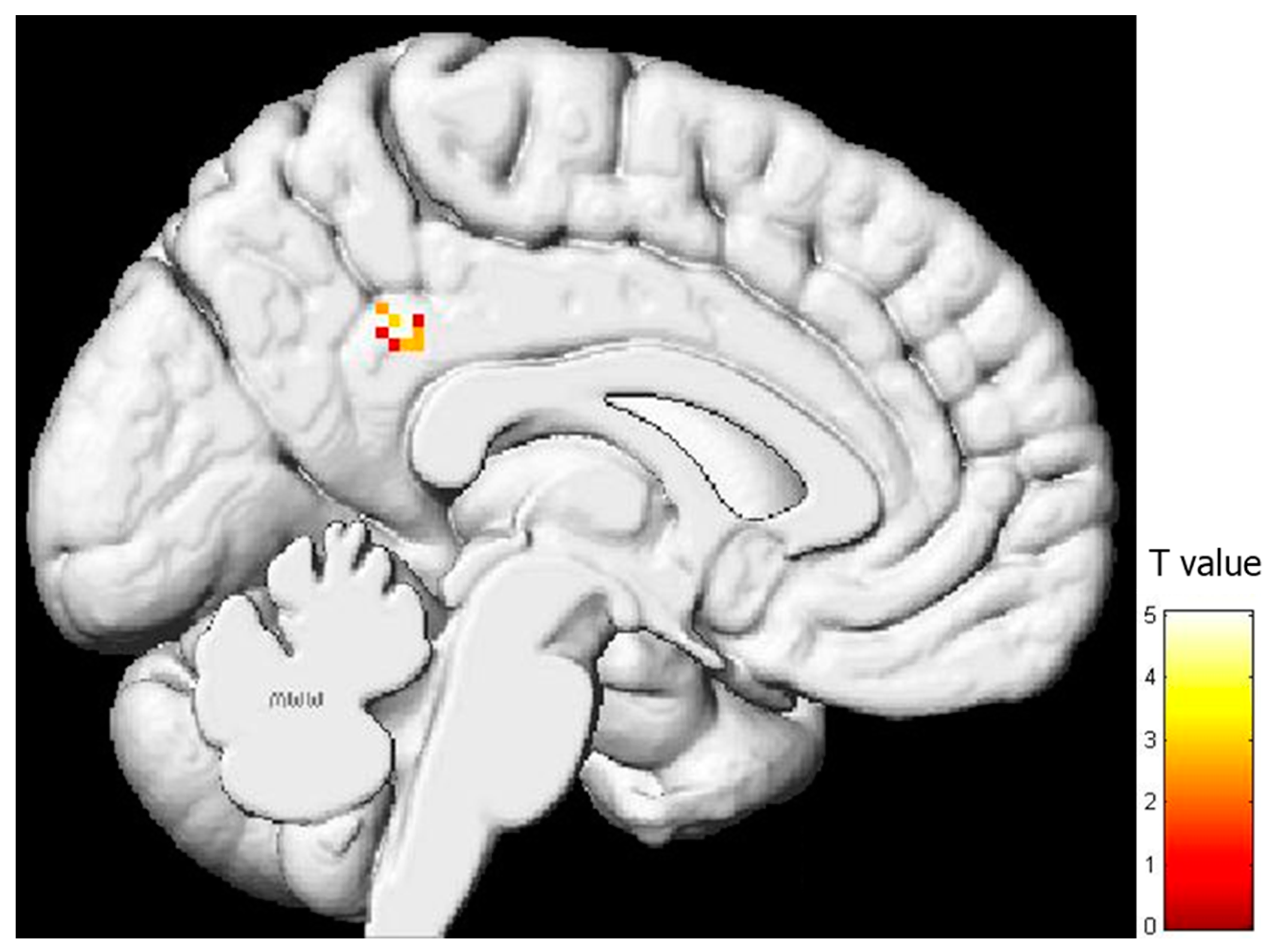
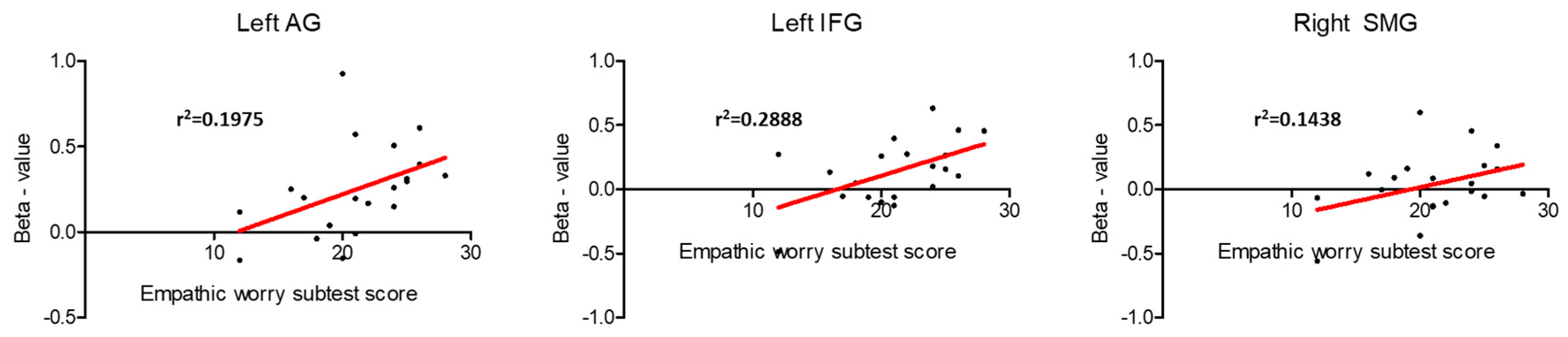
3.2. Regression Analysis between Brain Activity during Pain Expression Observation and IRI Subscale Scores
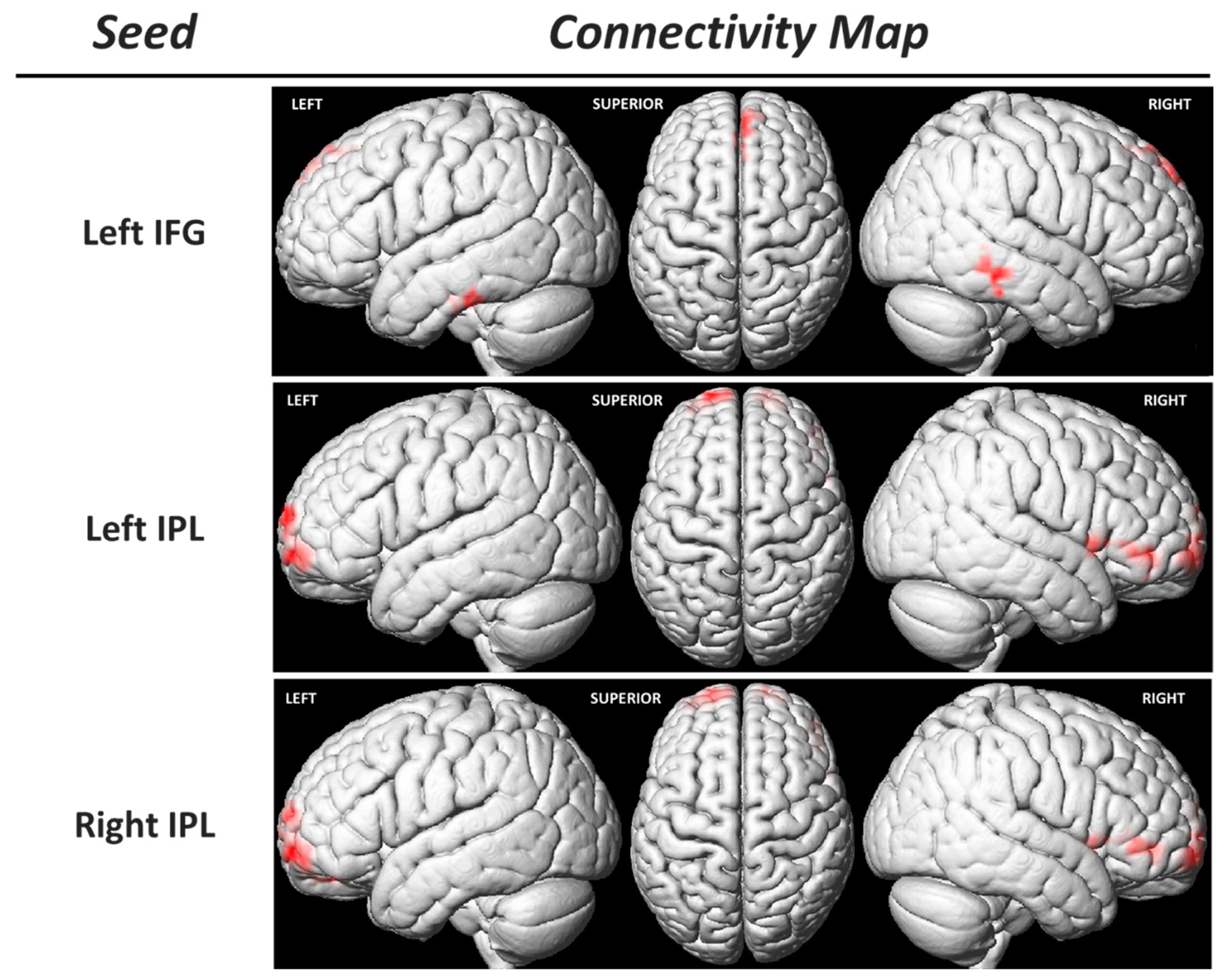
3.3. Functional Connectivity of the Sensorimotor MNS Core Regions
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xiong, R.-C.; Fu, X.; Wu, L.-Z.; Zhang, C.-H.; Wu, H.-X.; Shi, Y.; Wu, W. Brain pathways of pain empathy activated by pained facial expressions: A meta-analysis of fMRI using the activation likelihood estimation method. Neural Regen. Res. 2019, 14, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.H. Measuring individual differences in empathy: Evidence for a multidimensional approach. J. Pers. Soc. Psychol. 1983, 44, 113–126. [Google Scholar] [CrossRef]
- Decety, J.; Bartal, I.B.-A.; Uzefovsky, F.; Knafo-Noam, A. Empathy as a driver of prosocial behaviour: Highly conserved neurobehavioural mechanisms across species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150077. [Google Scholar] [CrossRef] [PubMed]
- Decety, J.; Michalska, K.J.; Akitsuki, Y. Who caused the pain? An fMRI investigation of empathy and intentionality in children. Neuropsychologia 2008, 46, 2607–2614. [Google Scholar] [CrossRef] [PubMed]
- Loggia, M.L.; Mogil, J.S.; Bushnell, M.C. Empathy hurts: Compassion for another increases both sensory and affective components of pain perception. Pain 2008, 136, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Fallon, N.; Roberts, C.; Stancak, A. Shared and distinct functional networks for empathy and pain processing: A systematic review and meta-analysis of fMRI studies. Soc. Cogn. Affect. Neurosci. 2020, 15, 709–723. [Google Scholar] [CrossRef]
- Singer, T.; Lamm, C. The social neuroscience of empathy. Ann. N. Y. Acad. Sci. 2009, 1156, 81–96. [Google Scholar] [CrossRef]
- Jackson, P.L.; Meltzoff, A.N.; Decety, J. How do we perceive the pain of others? A window into the neural processes involved in empathy. Neuroimage 2005, 24, 771–779. [Google Scholar] [CrossRef]
- Jackson, P.L.; Brunet, E.; Meltzoff, A.N.; Decety, J. Empathy examined through the neural mechanisms involved in imagining how I feel versus how you feel pain. Neuropsychologia 2006, 44, 752–761. [Google Scholar] [CrossRef]
- Morrison, I.; Lloyd, D.; di Pellegrino, G.; Roberts, N. Vicarious responses to pain in anterior cingulate cortex: Is empathy a multisensory issue? Cogn. Affect. Behav. Neurosci. 2004, 4, 270–278. [Google Scholar] [CrossRef]
- Lamm, C.; Decety, J.; Singer, T. Meta-analytic evidence for common and distinct neural networks associated with directly experienced pain and empathy for pain. Neuroimage 2011, 54, 2492–2502. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Camarda, R.; Fogassi, L.; Gentilucci, M.; Luppino, G.; Matelli, M. Functional organization of inferior area 6 in the macaque monkey. II. Area F5 and the control of distal movements. Exp. Brain Res. 1988, 71, 491–507. [Google Scholar] [CrossRef]
- di Pellegrino, G.; Fadiga, L.; Fogassi, L.; Gallese, V.; Rizzolatti, G. Understanding motor events: A neurophysiological study. Exp. Brain Res. 1992, 91, 176–180. [Google Scholar] [CrossRef]
- Gallese, V.; Fadiga, L.; Fogassi, L.; Rizzolatti, G. Action recognition in the premotor cortex. Brain 1996, 119 Pt 2, 593–609. [Google Scholar] [CrossRef] [PubMed]
- Heyes, C.; Catmur, C. What Happened to Mirror Neurons? Perspect. Psychol. Sci. 2022, 17, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Bonini, L.; Rotunno, C.; Arcuri, E.; Gallese, V. Mirror neurons 30 years later: Implications and applications. Trends Cogn. Sci. 2022, 26, 767–781. [Google Scholar] [CrossRef]
- de Gelder, B.; de Borst, A.W.; Watson, R. The perception of emotion in body expressions. Wiley Interdiscip. Rev. Cogn. Sci. 2014, 6, 149–158. [Google Scholar] [CrossRef] [PubMed]
- de Waal, F.B.M.; Preston, S.D. Mammalian empathy: Behavioural manifestations and neural basis. Nat. Rev. Neurosci. 2017, 18, 498–509. [Google Scholar] [CrossRef]
- Golland, Y.; Mevorach, D.; Levit-Binnun, N. Affiliative zygomatic synchrony in co-present strangers. Sci. Rep. 2019, 9, 3120. [Google Scholar] [CrossRef]
- Livneh, U.; Resnik, J.; Shohat, Y.; Paz, R. Self-monitoring of social facial expressions in the primate amygdala and cingulate cortex. Proc. Natl. Acad. Sci. USA 2012, 109, 18956–18961. [Google Scholar] [CrossRef]
- Caruana, F.; Jezzini, A.; Sbriscia-Fioretti, B.; Rizzolatti, G.; Gallese, V. Emotional and social behaviors elicited by electrical stimulation of the insula in the macaque monkey. Curr. Biol. 2011, 21, 195–199. [Google Scholar] [CrossRef]
- Wicker, B.; Keysers, C.; Plailly, J.; Royet, J.P.; Gallese, V.; Rizzolatti, G. Both of us disgusted in My insula: The common neural basis of seeing and feeling disgust. Neuron 2003, 40, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Calderazzo, S.M.; Busch, S.E.; Moore, T.L.; Rosene, D.L.; Medalla, M. Distribution and overlap of entorhinal, premotor, and amygdalar connections in the monkey anterior cingulate cortex. J. Comp. Neurol. 2021, 529, 885–904. [Google Scholar] [CrossRef] [PubMed]
- Allan, P.G.; Briggs, R.G.; Conner, A.K.; O’Neal, C.M.; Bonney, P.A.; Maxwell, B.D.; Baker, C.M.; Burks, J.D.; Sali, G.; Glenn, C.A.; et al. Parcellation-based tractographic modeling of the ventral attention network. J. Neurol. Sci. 2020, 408, 116548. [Google Scholar] [CrossRef] [PubMed]
- Botvinick, M.; Jha, A.P.; Bylsma, L.M.; Fabian, S.A.; Solomon, P.E.; Prkachin, K.M. Viewing facial expressions of pain engages cortical areas involved in the direct experience of pain. Neuroimage 2005, 25, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Leube, D.T.; Erb, M.; Grodd, W.; Bartels, M.; Kircher, T.T. Differential activation in parahippocampal and prefrontal cortex during word and face encoding tasks. Neuroreport 2001, 12, 2773–2777. [Google Scholar] [CrossRef] [PubMed]
- Lotze, M.; Heymans, U.; Birbaumer, N.; Veit, R.; Erb, M.; Flor, H.; Halsband, U. Differential cerebral activation during observation of expressive gestures and motor acts. Neuropsychologia 2006, 44, 1787–1795. [Google Scholar] [CrossRef]
- Cheng, Y.; Yang, C.-Y.; Lin, C.-P.; Lee, P.-L.; Decety, J. The perception of pain in others suppresses somatosensory oscillations: A magnetoencephalography study. Neuroimage 2008, 40, 1833–1840. [Google Scholar] [CrossRef]
- Ogino, Y.; Nemoto, H.; Inui, K.; Saito, S.; Kakigi, R.; Goto, F. Inner experience of pain: Imagination of pain while viewing images showing painful events forms subjective pain representation in human brain. Cereb. Cortex 2007, 17, 1139–1146. [Google Scholar] [CrossRef]
- Gu, X.; Han, S. Attention and reality constraints on the neural processes of empathy for pain. Neuroimage 2007, 36, 256–267. [Google Scholar] [CrossRef]
- Mazza, M.; Tempesta, D.; Pino, M.C.; Nigri, A.; Catalucci, A.; Guadagni, V.; Gallucci, M.; Iaria, G.; Ferrara, M. Neural activity related to cognitive and emotional empathy in post-traumatic stress disorder. Behav. Brain Res. 2015, 282, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Maldjian, J.A.; Laurienti, P.J.; Kraft, R.A.; Burdette, J.H. An automated method for neuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets. Neuroimage 2003, 19, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Caspers, S.; Geyer, S.; Schleicher, A.; Mohlberg, H.; Amunts, K.; Zilles, K. The human inferior parietal cortex: Cytoarchitectonic parcellation and interindividual variability. Neuroimage 2006, 33, 430–448. [Google Scholar] [CrossRef]
- Caspers, S.; Eickhoff, S.B.; Geyer, S.; Scheperjans, F.; Mohlberg, H.; Zilles, K.; Amunts, K. The human inferior parietal lobule in stereotaxic space. Brain Struct. Funct. 2008, 212, 481–495. [Google Scholar] [CrossRef]
- Fox, M.D.; Raichle, M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007, 8, 700–711. [Google Scholar] [CrossRef]
- Song, X.-W.; Dong, Z.-Y.; Long, X.-Y.; Li, S.-F.; Zuo, X.-N.; Zhu, C.-Z.; He, Y.; Yan, C.-G.; Zang, Y.-F. REST: A toolkit for resting-state functional magnetic resonance imaging data processing. PLoS ONE 2011, 6, e25031. [Google Scholar] [CrossRef]
- Gómez-Leal, R.; Costa, A.; Megías-Robles, A.; Fernández-Berrocal, P.; Faria, L. Relationship between emotional intelligence and empathy towards humans and animals. PeerJ 2021, 9, e11274. [Google Scholar] [CrossRef]
- Montgomery, K.J.; Haxby, J.V. Mirror Neuron System Differentially Activated by Facial Expressions and Social Hand Gestures: A Functional Magnetic Resonance Imaging Study. J. Cogn. Neurosci. 2008, 20, 1866–1877. [Google Scholar] [CrossRef]
- Wriessnegger, S.C.; Leeb, R.; Kaiser, V.; Neuper, C.; Müller-Putz, G.R. Watching object related movements modulates mirror-like activity in parietal brain regions. Clin. Neurophysiol. 2013, 124, 1596–1604. [Google Scholar] [CrossRef]
- Campbell, M.E.J.; Mehrkanoon, S.; Cunnington, R. Intentionally not imitating: Insula cortex engaged for top-down control of action mirroring. Neuropsychologia 2018, 111, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.A.; Jones, A.K. A role for midcingulate cortex in the interruptive effects of pain anticipation on attention. Clin. Neurophysiol. 2008, 119, 2370–2379. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; White, N.S.; Kwong, K.K.; Vangel, M.G.; Rosman, I.S.; Gracely, R.H.; Gollub, R.L. Using fMRI to dissociate sensory encoding from cognitive evaluation of heat pain intensity. Hum. Brain Mapp. 2006, 27, 715–721. [Google Scholar] [CrossRef]
- Tominaga, W.; Matsubayashi, J.; Deguchi, Y.; Minami, C.; Kinai, T.; Nakamura, M.; Nagamine, T.; Matsuhashi, M.; Mima, T.; Fukuyama, H.; et al. A mirror reflection of a hand modulates stimulus-induced 20-Hz activity. NeuroImage 2009, 46, 500–504. [Google Scholar] [CrossRef]
- Minio-Paluello, I.; Avenanti, A.; Aglioti, S.M. Left hemisphere dominance in reading the sensory qualities of others’ pain? Soc. Neurosci. 2006, 1, 320–333. [Google Scholar] [CrossRef]
- Merskey, H.; Watson, G.D. The lateralisation of pain. Pain 1979, 7, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Sarkheil, P.; Goebel, R.; Schneider, F.; Mathiak, K. Emotion unfolded by motion: A role for parietal lobe in decoding dynamic facial expressions. Soc. Cogn. Affect. Neurosci. 2013, 8, 950–957. [Google Scholar] [CrossRef]
- Frost, N.A.; Haggart, A.; Sohal, V.S. Dynamic patterns of correlated activity in the prefrontal cortex encode information about social behavior. PLoS Biol. 2021, 19, e3001235. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, V.; Vuilleumier, P.; Latorre, V.; Petito, A.; Gallese, V.; Popolizio, T.; Arciero, G.; Bondolfi, G. Effects of Emotional Contexts on Cerebello-Thalamo-Cortical Activity during Action Observation. PLoS ONE 2013, 8, e75912, Correction in PLoS ONE 2013, 8, 10.1371/annotation/73ee01f4-5b78-42c1-a4be-d48aa8ffd51d. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Gazzola, V.; Avenanti, A. Temporal dynamics of motor cortex excitability during perception of natural emotional scenes. Soc. Cogn. Affect. Neurosci. 2014, 9, 1451–1457. [Google Scholar] [CrossRef]
- Mukamel, R.; Ekstrom, A.D.; Kaplan, J.; Iacoboni, M.; Fried, I. Single-neuron responses in humans during execution and observation of actions. Curr. Biol. 2010, 20, 750–756. [Google Scholar] [CrossRef]
- Papadourakis, V.; Raos, V. Evidence for the representation of movement kinematics in the discharge of F5 mirror neurons during the observation of transitive and intransitive actions. J. Neurophysiol. 2017, 118, 3215–3229. [Google Scholar] [CrossRef] [PubMed]
- Molnar-Szakacs, I.; Kaplan, J.; Greenfield, P.M.; Iacoboni, M. Observing complex action sequences: The role of the fronto-parietal mirror neuron system. NeuroImage 2006, 33, 923–935. [Google Scholar] [CrossRef] [PubMed]
- De Lima, F.F.; de Osório, F.L. Empathy: Assessment Instruments and Psychometric Quality—A Systematic Literature Review With a Meta-Analysis of the Past Ten Years. Front. Psychol. 2021, 12, 781346. [Google Scholar] [CrossRef] [PubMed]
- Chrysikou, E.G.; Thompson, W.J. Assessing Cognitive and Affective Empathy Through the Interpersonal Reactivity Index. Assessment 2016, 23, 769–777. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | BA | x | y | z | T | Z | Voxels/Cluster |
|---|---|---|---|---|---|---|---|
| OCCIPITAL LOBE | |||||||
| Left Middle Occipital Gyrus | 19 | −40 | −68 | 4 | 4.69 | 3.71 | 54 |
| Left Middle Occipital Gyrus | 19 | −48 | −72 | 4 | 4.02 | 3.33 | |
| Left Lingual Gyrus | 18 | −16 | −82 | −18 | 5.25 | 3.99 | 58 |
| Left Inferior Occipital Gyrus | 17 | −10 | −94 | −14 | 5.17 | 3.96 | |
| Left Middle Occipital Gyrus | 18 | −46 | −78 | −16 | 5.17 | 3.95 | 50 |
| Left Lingual Gyrus | 17 | −10 | −96 | −8 | 4.70 | 3.71 | 28 |
| Left Lingual Gyrus | 17 | −18 | −94 | −4 | 4.41 | 3.55 | |
| TEMPORAL LOBE | |||||||
| Left Superior Temporal Gyrus | 22 | −38 | −58 | 12 | 5.50 | 4.12 | 59 |
| Left Middle Temporal Gyrus | 19 | −52 | −64 | 12 | 4.07 | 3.35 | |
| Right Middle Temporal Gyrus | 37 | 48 | −66 | 0 | 4.07 | 3.36 | 28 |
| PARIETAL LOBE | |||||||
| Left Supramarginal Gyrus | 40 | −62 | −46 | 28 | 4.03 | 3.33 | 23 |
| Left Inferior Parietal Lobule | 40 | −42 | −60 | 44 | 5.11 | 3.92 | 138 |
| Left Inferior Parietal Lobule | 40 | −34 | −68 | 48 | 4.23 | 3.45 |
| Region | BA | x | y | z | T | Z | Voxels/Cluster | r2 |
|---|---|---|---|---|---|---|---|---|
| FRONTAL LOBE | ||||||||
| Right Superior Frontal Gyrus | 6, 8 | 8 | 6 | 66 | 6.16 | 3.76 | 39 | 0.228 |
| 8 | 42 | 50 | 4.92 | 3.35 | 25 | 0.206 | ||
| 2 | 40 | 44 | 4.59 | 3.21 | 0.198 | |||
| Left Superior and Medial Frontal Gyrus | 6, 8, 9 | −6 | 28 | 60 | 5.57 | 3.58 | 87 | 0.219 |
| −6 | 30 | 52 | 5.21 | 3.45 | 0.212 | |||
| −2 | 20 | 52 | 4.62 | 3.23 | 0.199 | |||
| −4 | −2 | 68 | 5.14 | 3.43 | 70 | 0.211 | ||
| −12 | 6 | 62 | 4.74 | 3.27 | 0.202 | |||
| −2 | 4 | 58 | 4.34 | 3.11 | 0.191 | |||
| −4 | 44 | 30 | 5.16 | 3.43 | 54 | 0.211 | ||
| −12 | 50 | 30 | 4.47 | 3.16 | 0.195 | |||
| −42 | 8 | 38 | 5.95 | 3.70 | 34 | 0.225 | ||
| Left Cingulate Gyrus | 23 | −2 | −20 | 34 | 5.18 | 3.44 | 27 | 0.211 |
| Right Cingulate Gyrus | 32 | 12 | 22 | 36 | 4.65 | 3.24 | 22 | 0.200 |
| 4 | 22 | 42 | 4.47 | 3.17 | 0.195 | |||
| PARIETAL LOBE | ||||||||
| Left Superior Parietal Lobule | 7 | −34 | −68 | 46 | 7.74 | 4.18 | 43 | 0.245 |
| Right Supramarginal Gyrus | 40 | 60 | −48 | 22 | 7.23 | 4.06 | 24 | 0.241 |
| TEMPORAL LOBE | ||||||||
| Left Middle Temporal Gyrus | 39 | −48 | −60 | 24 | 5.72 | 3.63 | 36 | 0.222 |
| Region | BA | x | y | z | T | Z | Voxels/Cluster | r2 |
|---|---|---|---|---|---|---|---|---|
| PARIETAL LOBE | ||||||||
| Left posterior cingulate gyrus | 31 | −14 | −44 | 32 | 5.05 | 3.39 | 25 | 0.506 |
| Region | BA | x | y | z | T | Z | Voxels/Cluster |
|---|---|---|---|---|---|---|---|
| Seed: Left IFG | |||||||
| Left Fusiform Gyrus | 20 | −51 | −21 | −30 | 4.01 | 3.32 | 49 |
| Right Supplementary Motor Area | 6 | 0 | 39 | 51 | 4.39 | 3.54 | 48 |
| 3 | 51 | 45 | 4.03 | 3.33 | |||
| 3 | 57 | 39 | 3.43 | 2.95 | |||
| Right Middle Temporal Gyrus | 21 | 69 | −30 | −12 | 4.05 | 3.34 | 57 |
| 66 | −33 | −24 | 3.95 | 3.28 | |||
| 57 | −36 | −15 | 3.87 | 3.23 | |||
| Seed: Left IPL | |||||||
| Left Superior Frontal Gyrus | 10 | −18 | 66 | −6 | 7.07 | 4.76 | 161 |
| −18 | 69 | 15 | 5.44 | 4.08 | |||
| Right Superior Frontal Gyrus | 10 | 18 | 63 | −6 | 6.73 | 4.64 | 89 |
| 15 | 69 | 3 | 4.72 | 3.72 | |||
| 24 | 66 | −12 | 4.61 | 3.66 | |||
| Right Superior Temporal Gyrus | 22 | 57 | 12 | −3 | 5.18 | 3.96 | 119 |
| Right Inferior Frontal Gyrus | 45 | 42 | 42 | −6 | 4.85 | 3.79 | |
| 48 | 36 | −6 | 4.73 | 3.73 | |||
| Seed: Right IPL | |||||||
| Right Superior Frontal Gyrus | 10 | 15 | 66 | −6 | 5.85 | 4.27 | 65 |
| 24 | 63 | −9 | 4.80 | 3.77 | |||
| 15 | 66 | 3 | 4.51 | 3.61 | |||
| Left Superior Frontal Gyrus | 10 | −18 | 63 | −6 | 5.46 | 4.10 | 122 |
| −33 | 60 | −12 | 4.28 | 3.48 | |||
| −15 | 66 | 15 | 4.48 | 3.59 | 43 | ||
| −9 | 66 | 21 | 4.46 | 3.58 | |||
| −24 | 66 | 9 | 4.14 | 3.39 | |||
| Right Middle Frontal Gyrus | 10 | 30 | 36 | −3 | 3.89 | 3.25 | 88 |
| Right Inferior Frontal Gyrus | 45 | 48 | 39 | −3 | 3.85 | 3.22 | |
| 48 | 48 | −6 | 3.68 | 3.11 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plata-Bello, J.; Privato, N.; Modroño, C.; Pérez-Martín, Y.; Borges, Á.; González-Mora, J.L. Empathy Modulates the Activity of the Sensorimotor Mirror Neuron System during Pain Observation. Behav. Sci. 2023, 13, 947. https://doi.org/10.3390/bs13110947
Plata-Bello J, Privato N, Modroño C, Pérez-Martín Y, Borges Á, González-Mora JL. Empathy Modulates the Activity of the Sensorimotor Mirror Neuron System during Pain Observation. Behavioral Sciences. 2023; 13(11):947. https://doi.org/10.3390/bs13110947
Chicago/Turabian StylePlata-Bello, Julio, Nicole Privato, Cristián Modroño, Yaiza Pérez-Martín, África Borges, and José Luis González-Mora. 2023. "Empathy Modulates the Activity of the Sensorimotor Mirror Neuron System during Pain Observation" Behavioral Sciences 13, no. 11: 947. https://doi.org/10.3390/bs13110947
APA StylePlata-Bello, J., Privato, N., Modroño, C., Pérez-Martín, Y., Borges, Á., & González-Mora, J. L. (2023). Empathy Modulates the Activity of the Sensorimotor Mirror Neuron System during Pain Observation. Behavioral Sciences, 13(11), 947. https://doi.org/10.3390/bs13110947