Acetate Activates Deep Subsurface Fracture Fluid Microbial Communities in Olkiluoto, Finland



Abstract
1. Introduction
2. Materials and Methods
3. Results
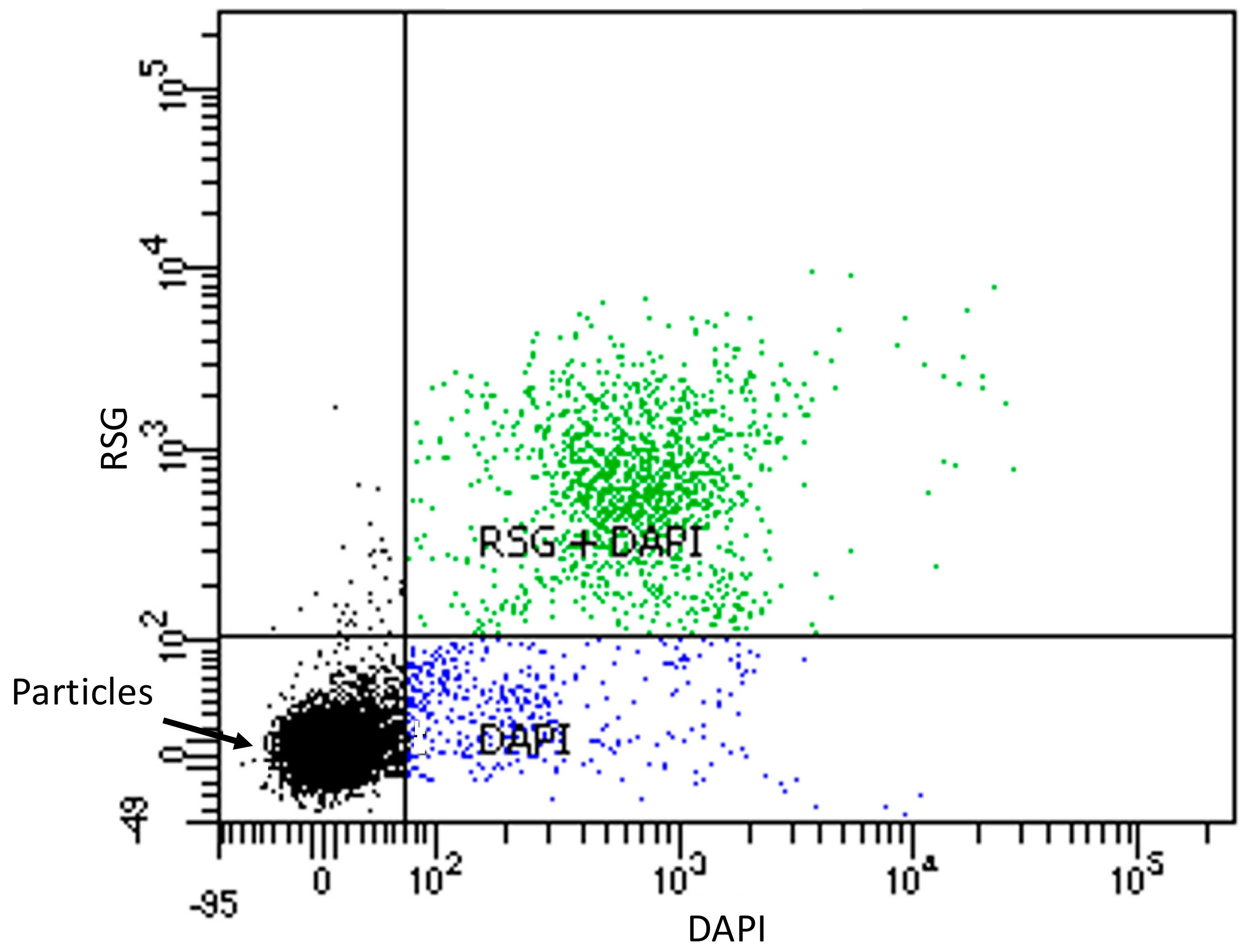
3.1. Microbial Community Analyses
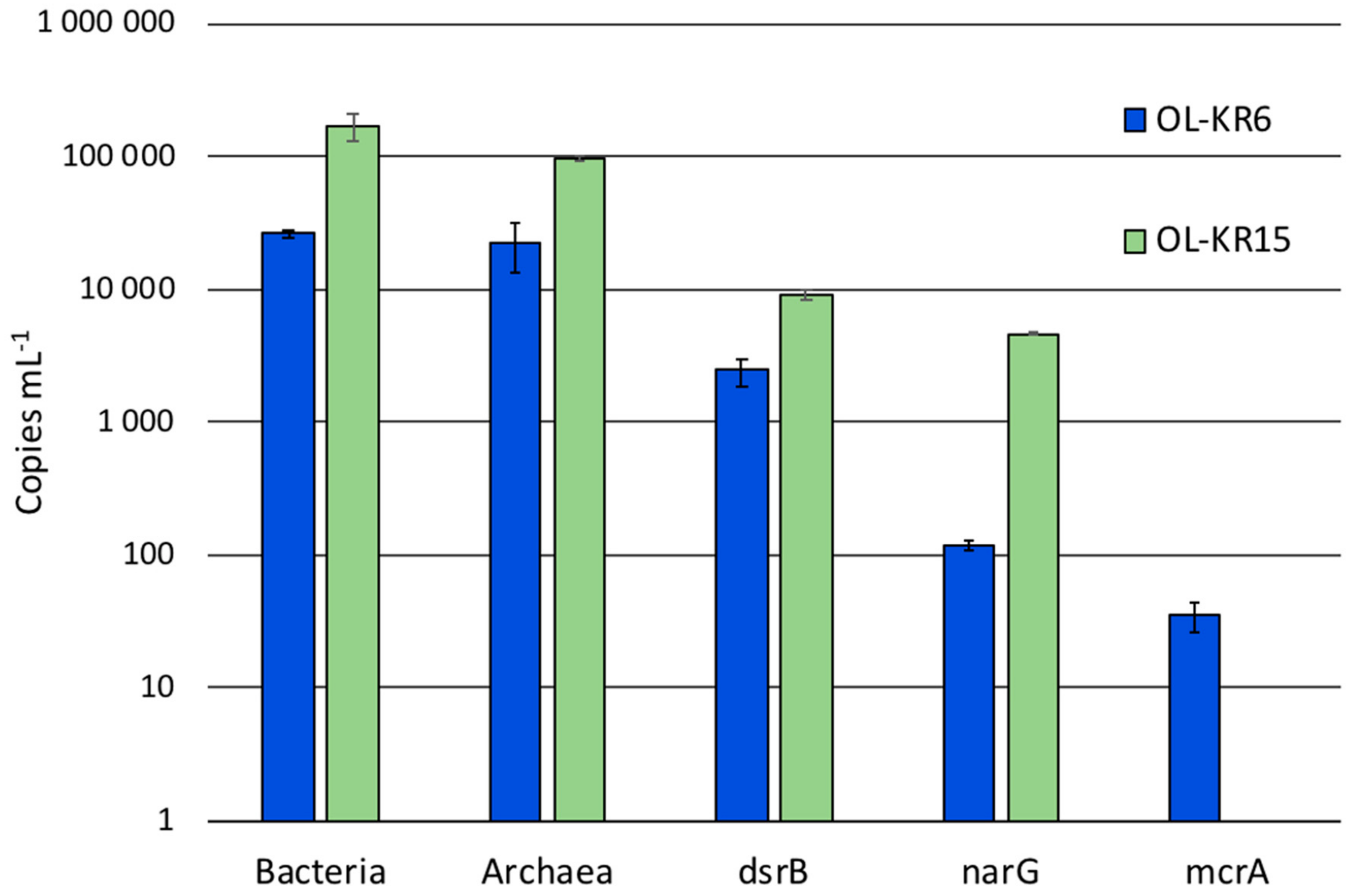
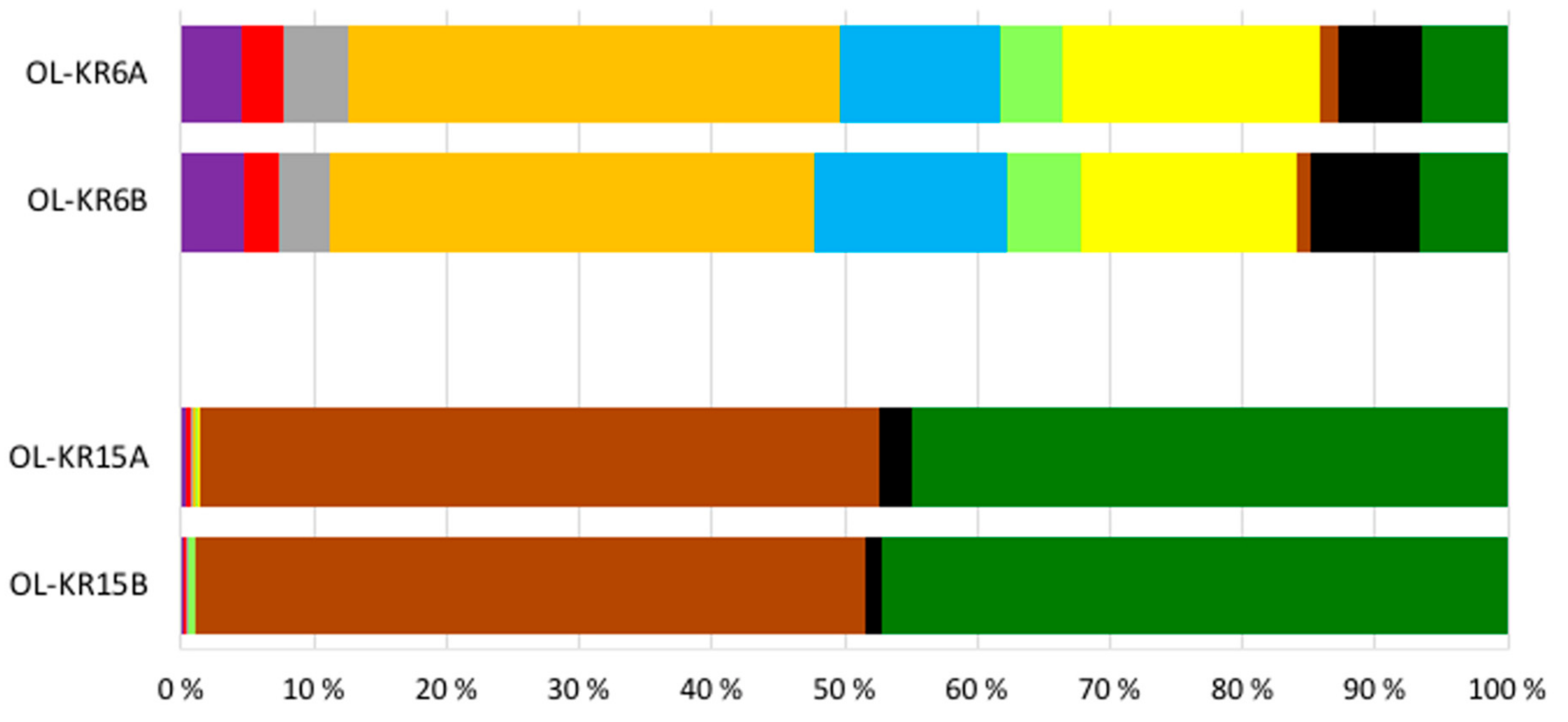
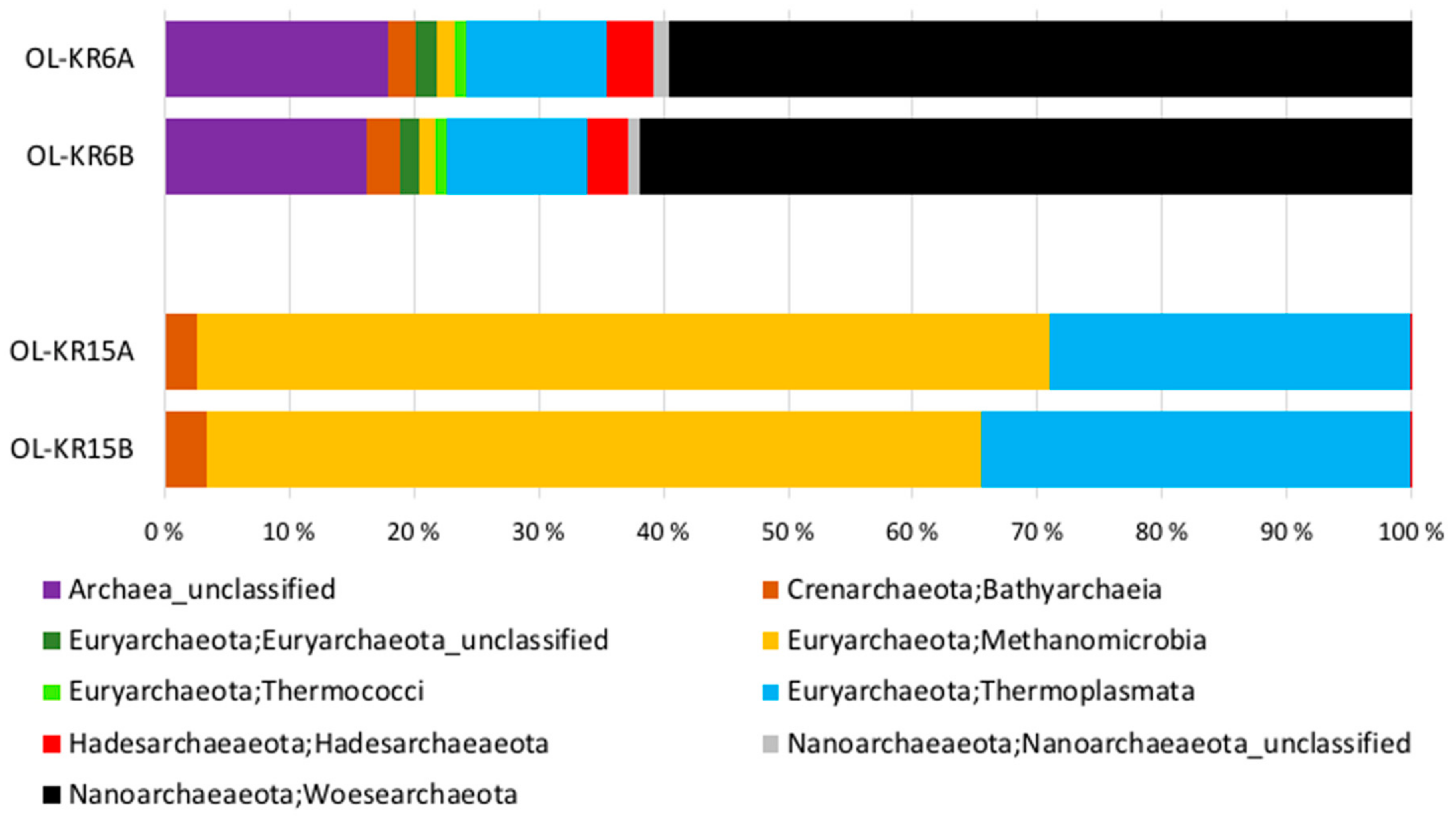
3.2. Sequence Statistics, Diversity Estimates and Microbial Communities in the Original Water
3.3. Activation of Microbial Communities in the Fracture Fluids
4. Discussions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lovley, D.R.; Chapelle, F.H. Deep subsurface microbial processes. Rev. Geophys. 1995, 33, 365–381. [Google Scholar] [CrossRef]
- Hoehler, T.M.; Jørgensen, B.B. Microbial life under extreme energy limitation. Nat. Rev. Microbiol. 2013, 11, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Bomberg, M.; Lamminmäki, T.; Itävaara, M. Microbial communities and their predicted metabolic characteristics in deep fracture groundwaters of the crystalline bedrock at Olkiluoto, Finland. Biogeosciences 2016, 13, 6031–6047. [Google Scholar] [CrossRef]
- Purkamoa, L.; Bomberg, M.; Kietäväinen, R.; Salavirta, H.; Nyyssönen, M.; Nuppunen-Puputti, M.; Ahonen, L.; Kukkonen, I.; Itävaara, M. Microbial co-occurrence patterns in deep Precambrian bedrock fracture fluids. Biogeosciences 2016, 13, 3091–3108. [Google Scholar] [CrossRef]
- Chivian, D.; Brodie, E.L.; Alm, E.J.; Culley, D.E.; Dehal, P.S.; DeSantis, T.Z.; Gihring, T.M.; Lapidus, A.; Lin, L.-H.; Lowry, S.R.; et al. Environmental genomics reveals a single-species ecosystem deep within earth. Science 2008, 322, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Posiva Ltd. Olkiluoto Site Description 2011, 1st ed.; Posiva Ltd.: Eurajoki, Finland, 2012; p. 1028. ISBN 978-951-652-179-7. Available online: www.posiva.fi/tietopankki/olkiluoto_site_description_2011.1793.xhtml#.Wwz2Uqm-l0s (accessed on 30 October 2018).
- Hammes, F.; Berney, M.; Egli, T. Cultivation-independent assessment of bacterial viability. In High Resolution Microbial Single Cell Analytics, 1st ed.; Müller, D., Bley, T., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2011; Volume 124, pp. 123–150. ISBN 978-3-642-16886-4. [Google Scholar]
- Del Giorgio, P.A.; Gasol, J.M. Physiological structure and single-cell activity in marine bacterioplankton. In Microbial Ecology of the Oceans, 2nd ed.; Kirshman, D.L., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2008; pp. 244–298. ISBN 978-0-470-04344-8. [Google Scholar]
- Lennon, J.T.; Jones, S.E. Microbial seed banks: The ecological and evolutionary implications of dormancy. Nat. Rev. Microbiol. 2011, 9, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Créach, V.; Baudoux, A.-C.; Bertru, G.; Le Rouzic, B. Direct estimate of active bacteria: CTC use and limitations. J. Microbiol. Methods 2003, 52, 19–28. [Google Scholar] [CrossRef]
- Bhupathiraju, V.; Hernandez, M.; Landfear, D.; Alvarez-Cohen, L. Application of a tetrazolium dye as an indicator of viability in anaerobic bacteria. J. Microbiol. Methods 1999, 37, 231–243. [Google Scholar] [CrossRef]
- Kalyuzhnaya, M.G.; Lindstrom, M.E.; Chistoserdova, L. Real-time detection of actively metabolizing microbes by redox sensing as applied to methylotroph populations in Lake Washington. ISME J. 2008, 2, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Kotelnikova, S.; Pedersen, K. Evidence for methanogenic Archaea and homoacetogenic Bacteria in deep granitic rock aquifers. FEMS Microbiol. Rev. 1997, 20, 339–349. [Google Scholar] [CrossRef]
- Haroon, M.F.; Hu, S.; Shi, Y.; Imelfort, M.; Keller, J.; Hugenholtz, P.; Yuan, Z.; Tyson, G.W. Anaerobic oxidation of methane coupled to nitrate reduction in a novel archaeal lineage. Nature 2013, 500, 567–570. [Google Scholar] [CrossRef] [PubMed]
- McMahon, P.B.; Chapelle, F.H. Microbial production of organic acids in aquitard sediments and its role in aquifer geochemistry. Nature 1991, 349, 233–235. [Google Scholar] [CrossRef]
- Lin, L.H.; Hall, J.; Lippmann-Pipke, J.; Ward, J.A.; Lollar, B.S.; DeFlaun, M.; Rothmel, R.; Moser, D.; Gihring, T.M.; Mislowack, B.; et al. Radiolytic H2 in continental crust: Nuclear power for deep subsurface microbial communities. Geochem. Geophys. Geosyst. 2005, 6, 1–13. [Google Scholar] [CrossRef]
- Lin, L.H.; Slater, G.F.; Sherwood Lollar, B.; Lacrampe-Couloume, G.; Onstott, T.C. The yield and isotopic composition of radiolytic H2, a potential energy source for the deep subsurface biosphere. Geochim. Cosmochim. Acta 2005, 69, 893–903. [Google Scholar] [CrossRef]
- Blomberg, P.; Itävaara, M.; Marjamaa, K.; Salavirta, H.; Arvas, M.; Miettinen, H.; Vikman, M. Metabolic Pathways of Deep Groundwater Microbiomes and Sulphide Formation at Olkiluoto, 1st ed.; Posiva Ltd.: Eurajoki, Finland, 2017; p. 150. Available online: www.posiva.fi/files/4688/WR_2017-11_web.pdf (accessed on 30 October 2018).
- Miettinen, H.; Bomberg, M.; Nyyssönen, M.; Salavirta, H.; Sohlberg, E.; Vikman, M.; Itävaara, M. The Diversity of Microbial Communities in Olkiluoto Bedrock Groundwaters 2009–2013; Posiva Ltd.: Eurajoki, Finland, 2015; p. 160. Available online: http://www.posiva.fi/files/4125/WR_2015-12.pdf (accessed on 30 October 2018).
- Bomberg, M.; Nyyssönen, M.; Pitkänen, P.; Lehtinen, A.; Itävaara, M. Active microbial communities inhabit sulphate-methane interphase in deep bedrock fracture fluids in Olkiluoto, Finland. BioMed Res. Int. 2015, 2015, 979530. [Google Scholar] [CrossRef] [PubMed]
- Bomberg, M.; Lamminmäki, T.; Itävaara, M. Estimation of microbial metabolism and co-occurrence patterns in fracture groundwaters of deep crystalline bedrock at Olkiluoto, Finland. Biogeosci. Discuss. 2015, 12, 13819–13857. [Google Scholar] [CrossRef]
- Weber, H.S.; Habicht, K.; Thamdrup, B. Anaerobic methanotrophic archaea of the ANME-2d cluster are active in a low-sulfate, iron-rich freshwater sediment. Front. Microbiol. 2017, 8, 619. [Google Scholar] [CrossRef] [PubMed]
- Oni, O.; Miyatake, T.; Kasten, S.; Richter-Heitmann, T.; Fischer, D.; Wagenknecht, L.; Kulkarni, A.; Blumers, M.; Shylin, S.I.; Ksenofontov, V.; et al. Distinct microbial populations are tightly linked to the profile of dissolved iron in the methanic sediments of the Helgoland mud area, North Sea. Front. Microbiol. 2015, 6, 365. [Google Scholar] [CrossRef] [PubMed]
- Purkamo, L.; Bomberg, M.; Nyyssönen, M.; Kukkonen, I.; Ahonen, L.; Kietäväinen, R.; Itävaara, M. Dissecting the deep biosphere: Retrieving authentic microbial communities from packer-isolated deep crystalline bedrock fracture zones. FEMS Microbiol. Ecol. 2013, 85, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Tsitko, I.; Lusa, M.; Lehto, J.; Parviainen, L.; Ikonen, A.T.K.; Lahdenperä, A-M.; Bomberg, M. The variation of microbial communities in a depth profile of an acidic, nutrient-poor boreal bog in Southwestern Finland. Open J. Ecol. 2014, 4, 832–859. [Google Scholar] [CrossRef]
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16s rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [PubMed]
- Bano, N.; Ruffin, S.; Ransom, B.; Hollibaugh, J.T. Phylogenetic composition of Arctic ocean archaeal assemblages and comparison with Antarctic assemblages. Appl. Environ. Microbiol. 2004, 70, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Barns, S.M.; Fundyga, R.E.; Jeffries, M.W.; Pace, N.R. Remarkable archaeal diversity detected in a Yellowstone National Park hot spring environment. Proc. Natl. Acad. Sci. USA 1994, 91, 1609–1613. [Google Scholar] [CrossRef] [PubMed]
- Rajala, P.; Bomberg, M.; Kietäväinen, R.; Kukkonen, I.; Ahonen, L.; Nyyssö∂nen, M.; Itävaara, M. Rapid reactivation of deep subsurface microbes in the presence of C-1 compounds. Microorganisms 2015, 3, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Kuever, J. The Family Desulfobacteraceae. In The Prokaryotes, 4th ed.; Rodenberg, E., DeLong, E.F., Loy, S., Stackebrndt, E., Thompson, F., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2014; pp. 45–74. ISBN 978-3-642-39044-9. [Google Scholar]
- Sun, H.; Spring, S.; Lapidus, A.; Davenport, K.; Glavina Del Rio, T.; Tice, J.; Nolan, M.; Copeland, A.; Cheng, J-F.; Lucas, S.; et al. Complete genome sequence of Desulfarculus baarsii type strain (2st14T). Stand. Genomic Sci. 2010, 3, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Labrenz, M.; Grote, J.; Mammitzsch, K.; Boschker, H.T.S.; Laue, M.; Jost, G.; Glaubitz, S.; Jörgens, K. Sulfurimonas gotlandica sp. nov., a chemoautotrophic and psychrotolerant epsilonproteobacterium isolated from a pelagic redoxcline, and an emended description of the genus Sulfurimonas. Int. J. Syst. Evol. Microbiol. 2013, 63, 4141–4148. [Google Scholar] [CrossRef] [PubMed]
- Löffler, F.E.; Yan, J.; Ritalahti, K.M.; Adrian, L.; Edwards, E.A.; Konstantinidis, K.T.; Müller, J.A.; Fullerton, H.; Zinder, S.H.; Spormann, A.M. Dehalococcoides mccartyi gen. nov., sp. nov., obligately organohalide-respiring anaerobic bacteria relevant to halogen cycling and bioremediation, belong to a novel bacterial class, Dehalococcoidia classis nov., order Dehalococcoidales ord. nov. and family Dehalococcoidaceae fam. nov., within the phylum Chloroflexi. Int. J. Syst. Evol. Microbiol. 2013, 63, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.J.; Saw, J.H.; Lind., A.E.; Lazar, C.S.; Hinrichs, K.-U.; Teske, A.P.; Ettema, T.J.G. Genomic inference of the metabolism of cosmopolitan subsurface Archaea, Hadesarchaea. Nat. Microbiol. 2016, 1, 16002. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.N.; Parks, D.H.; Chadwick, G.L.; Robbins, S.J.; Orphan, V.J.; Golding, S.D.; Tyson, G.W. Methane metabolism in the archaeal phylum Bathyarchaeota revealed by genome-centric metagenomics. Science 2015, 350, 434. [Google Scholar] [CrossRef] [PubMed]
- Lamminmäki, T.; (Posiva Ltd., Eurajoki, Finland). Personal communication, 2018.
- Bell, E.; Lamminmäki, T.; Alneberg, J.; Andersson, A.F.; Qian, C.; Xiong, W.; Hettich, R.L.; Balmer, L.; Frutschi, M.; Sommer, G.; et al. Biogehochemmical cycling by a low-diversity microbial community in deep groundwater. Front. Microbiol. 2018, 9, 2129. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.; Arlinger, J.; Eriksson, S.; Hallbeck, A.; Hallbeck, L.; Johansson, J. Numbers, biomass and cultivable diversity of microbial populations relate to depth and borehole-specific conditions in groundwater from depths of 4–450 m in Olkiluoto, Finland. ISME J. 2008, 2, 760–775. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; Stams, A.J.M. The ecology and biotechnology of sulphate-reducing bacteria. Nat. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Purkamo, L.; Bomberg, M.; Nyyssönen, M.; Ahonen, L.; Kukkonen, I.; Itävaara, M. Response of deep subsurface microbial community to different carbon sources and electron acceptors during ∼2 months incubation in microcosms. Front. Microbiol. 2017, 8, 232. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.H.; Park, M.; Lee, J.R.; Yun, P.-Y.; Jeon, C.O. Brevundimonas aveniformis sp. nov., a stalked species isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 2007, 57, 1561–1565. [Google Scholar] [CrossRef] [PubMed]
- Ino, K.; Hernsdorf, AW.; Konno, U.; Kouduka, M.; Yanagawa, K.; Kato, S.; Sunamura, M.; Hirota, A.; Togo, Y.; Ito, K.; et al. Ecological and genomic profiling of anaerobic methane-oxidizing archaea in a deep granitic environment. ISME J. 2018, 12, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Scheller, S.; Yu, H.; Chadwick, G.L.; McGlynn, S.E.; Orphan, V.J. Artificial electron acceptors decouple archaeal methane oxidation from sulfate reduction. Science 2016, 351, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Rasigraf, O.; Sapart, C.J.; Jilbert, T.; Jetten, M.S.; Röckmann, T.; van der Veen, C.; Bândă, N.; Kartal, B.; Ettwig, K.F.; et al. Iron-mediated anaerobic oxidation of methane in brackish coastal sediments. Environ. Sci. Technol. 2015, 49, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Beal, E.J.; House, C.H.; Orphan, V.J. Manganese- and iron-dependent marine methane oxidation. Science 2009, 325, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Hatzinger, P.B.; Palmer, P.; Smith, R.L.; Peñrrieta, C.T.; Yoshinari, T. Applicability of tetrazolium salts for the measurement of respiratory activity and viability of groundwater bacteria. J. Microbiol. Methods 2003, 52, 47–58. [Google Scholar] [CrossRef]
- Bomberg, M.; Raulio, M.; Jylhä, S.; Mueller, C.W.; Höschen, C.; Rajala, P.; Purkamo, L.; Kietäväinen, R.; Ahonen, L.; Itävaara, M. CO2 and carbonate as substrate for the activation of the microbial community in 180 m deep bedrock fracture fluid of Outokumpu Deep Drill Hole, Finland. AIMS Microbiol. 2017, 3, 846–871. [Google Scholar] [CrossRef]
- Rajala, P.; Bomberg, M. Reactivation of deep subsurface microbial community in response to methane or methanol amendment. Front. Microbiol. 2017, 8, 431. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Perner, M. The globally widespread genus Sulfurimonas: Versatile energy metabolisms and adaptations to redox clines. Front. Microbiol. 2015, 6, 989. [Google Scholar] [CrossRef] [PubMed]
- Sias, S.R.; Ingraham, J.L. Isolation and analysis of mutants of Pseudomonas aeruginosa unable to assimilate nitrate. Arch. Microbiol. 1979, 122, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Bedzyk, L.; Wang, T.; Ye, R.W. The periplasmic nitrate reductase in Pseudomonas sp. G-179 catalyzes the first step of denitrification. J. Bacteriol. 1999, 181, 2801–2806. [Google Scholar]
- Silver, B.J.; Raymond, R.; Sigman, D.M.; Prokopeko, M.; Sherwood Lollar, B.; Lacrampe-Couloume, G.; Fogel, M.L.; Pratt, L.M.; Lefticariu, L.; Onstott, T.C. The origin of NO3− and N2 in deep subsurface fracture water of South Africa. Chem. Geol. 2012, 294, 51–62. [Google Scholar] [CrossRef]
- Kutvonen, H.; Rajala, P.; Carpén, L.; Bomberg, M. Nitrate and ammonia as nitrogen sources for deep subsurface microorganisms. Front. Microbiol. 2015, 6, 1079. [Google Scholar] [CrossRef] [PubMed]
- Castelle, C.J.; Wrighton, K.C.; Thomas, B.C.; Hug, L.A.; Brown, C.T.; Wilkins, M.J. Genomic expansion of domain archaea highlights roles for organisms from new phyla in anaerobic carbon cycling. Curr. Biol. 2015, 25, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Wrighton, K.C.; Thomas, B.C.; Sharon, I.; Miller, C.S.; Castelle, C.J.; VerBerkmoes, N.C.; Wilkins, M.J.; Hettich, R.L.; Lipton, M.S.; Williams, K.H.; et al. Fermentation, hydrogen, and sulfur metabolism in multiple uncultivated bacterial phyla. Science 2012, 337, 1661–1665. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.T.; Hug, L.A.; Thomas, B.C.; Sharon, T.; Castelle, C.J.; Singh, A.; Wilkins, M.J.; Wrighton, K.C.; Williams, K.H.; Banfield, J.F. Unusual biology across a group comprising more than 15% of domain Bacteria. Nature 2015, 523, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Nelson, W.C.; Stegen, J.C. The reduced genomes of Parcubacteria (OD1) contain signatures of a symbiotic lifestyle. Front. Microbiol. 2015, 6, 713. [Google Scholar] [CrossRef] [PubMed]
- Kizina, J. Insights into the Biology of Candidate Division OP3 LiM Populations. Ph.D. Thesis, University of Bremen, Bremen, Germany, 18 September 2017. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drillhole | OL-KR6 | OL-KR15 | Method |
| Drillhole section (m) | 125–130 | 446–460 | |
| Sampling date | 23 July 2017 | 31 January 2017 | |
| Vertical depth (m b.s.l.) | 94 | 434 | |
| EC (mS m−1) | 1270 | 2700 | SFS-EN-27888 |
| pH | 7.8 | 8.1 | ISO-10532 |
| Alkalinity (mmol L−1) | 2.2 | 0.2 | Titration |
| HCO3 (mg L−1) | 134 | 12 | Calculated from alkalinity |
| NPOC (mg L−1) | 4 | 11 | SFS-EN 1484 |
| (mg L−1) | <0.2 | <0.4 | IC, conductivity detector |
| Ntot (mg L−1) | 0.35 | 1.3 | IC, conductivity detector |
| (mg L−1) | 0.34 | 0.04 | SFS 3032 to an appropriate extent |
| Stot (mg L−1) | 140 | 2.7 | H2O2 oxidation + IC |
| (mg L−1) | 435 | 6.3 | IC, Conductivity detector |
| (mg L−1) | na | 0.25 | Spectrophotometry SFS 3038 to an appropriate extent |
| Fetot (mg L−1) | 0.3 | 0.06 | ICP-OES SFS-EN ISO 11885 |
| Fe(II) (mg L−1) | na | 0.07 | Spectrophotometry, ASTM E1615-08 to an appropriate extent |
| K (mg L−1) | 19 | 11 | ICP-OES SFS-EN ISO 11885 |
| Mg (mg L−1) | 173 | 34 | ICP-OES SFS-EN ISO 11885 |
| Ca (mg L−1) | 620 | 2050 | ICP-OES SFS-EN ISO 11885 |
| Cl (mg L−1) | 3650 | 9050 | SFS 3006 to an appropriate extent |
| Na (mg L−1) | 1510 | 3230 | ICP-OES SFS-EN ISO 11885 |
| Acetate (mg L−1) | <0.3 | <0.6 | IC, conductivity detector |
| Activating Electron Donors and Acceptors (Final Concentration in the Fracture Fluid Samples) | OL-KR6/125–130 m | OL-KR15/446–460 m |
|---|---|---|
| No activation | X | X |
| Electron donors | ||
| Acetate 1 mM | X | X |
| Hydrogen 0.08 mM | X | X |
| Sulphide 0.15 mM | X | X |
| Methane 1 mM | X | X |
| Terminal electron acceptors | ||
| Sodium nitrate 1 mM | X | X |
| Sodium sulphate 1 mM | X | |
| Combination of electron donors and acceptors | ||
| Acetate 1 mM and Sodium sulphate 1 mM | X | |
| Hydrogen 0.08 mM and Sodium sulphate 1 mM | X | |
| Methane 1 mM and Sodium sulphate 1 mM | X | |
| Methane 1 mM and Nitrate 1 mM | X | X |
| OL-KR6 A/B | OL-KR15 A/B | |
| Bacteria | ||
| Sequence reads originally | 6711/6520 | 16,958/5608 |
| Observed OTUs | 1641/1705 | 329/337 |
| Chao1 | 3033/2976 | 573/518 |
| Singletons | 869/881 | 142/131 |
| Doubletons | 270/304 | 40/46 |
| H’ | 9.7/9.8 | 6.2/6.4 |
| Archaea | ||
| Sequence reads originally | 13,699/13,513 | 15,872/16,827 |
| Observed OTUs | 1592/1545 | 284/289 |
| Chao1 | 2271/2259 | 421/417 |
| Singletons | 625/610 | 97/103 |
| Doubletons | 286/259 | 33/40 |
| H′ | 8.6/8.6 | 5.8/5.8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miettinen, H.; Bomberg, M.; Vikman, M. Acetate Activates Deep Subsurface Fracture Fluid Microbial Communities in Olkiluoto, Finland. Geosciences 2018, 8, 399. https://doi.org/10.3390/geosciences8110399
Miettinen H, Bomberg M, Vikman M. Acetate Activates Deep Subsurface Fracture Fluid Microbial Communities in Olkiluoto, Finland. Geosciences. 2018; 8(11):399. https://doi.org/10.3390/geosciences8110399
Chicago/Turabian StyleMiettinen, Hanna, Malin Bomberg, and Minna Vikman. 2018. "Acetate Activates Deep Subsurface Fracture Fluid Microbial Communities in Olkiluoto, Finland" Geosciences 8, no. 11: 399. https://doi.org/10.3390/geosciences8110399
APA StyleMiettinen, H., Bomberg, M., & Vikman, M. (2018). Acetate Activates Deep Subsurface Fracture Fluid Microbial Communities in Olkiluoto, Finland. Geosciences, 8(11), 399. https://doi.org/10.3390/geosciences8110399