Paleoenvironment Variability during Termination I at the Reykjanes Ridge, North Atlantic

 ,
, 
Abstract
1. Introduction
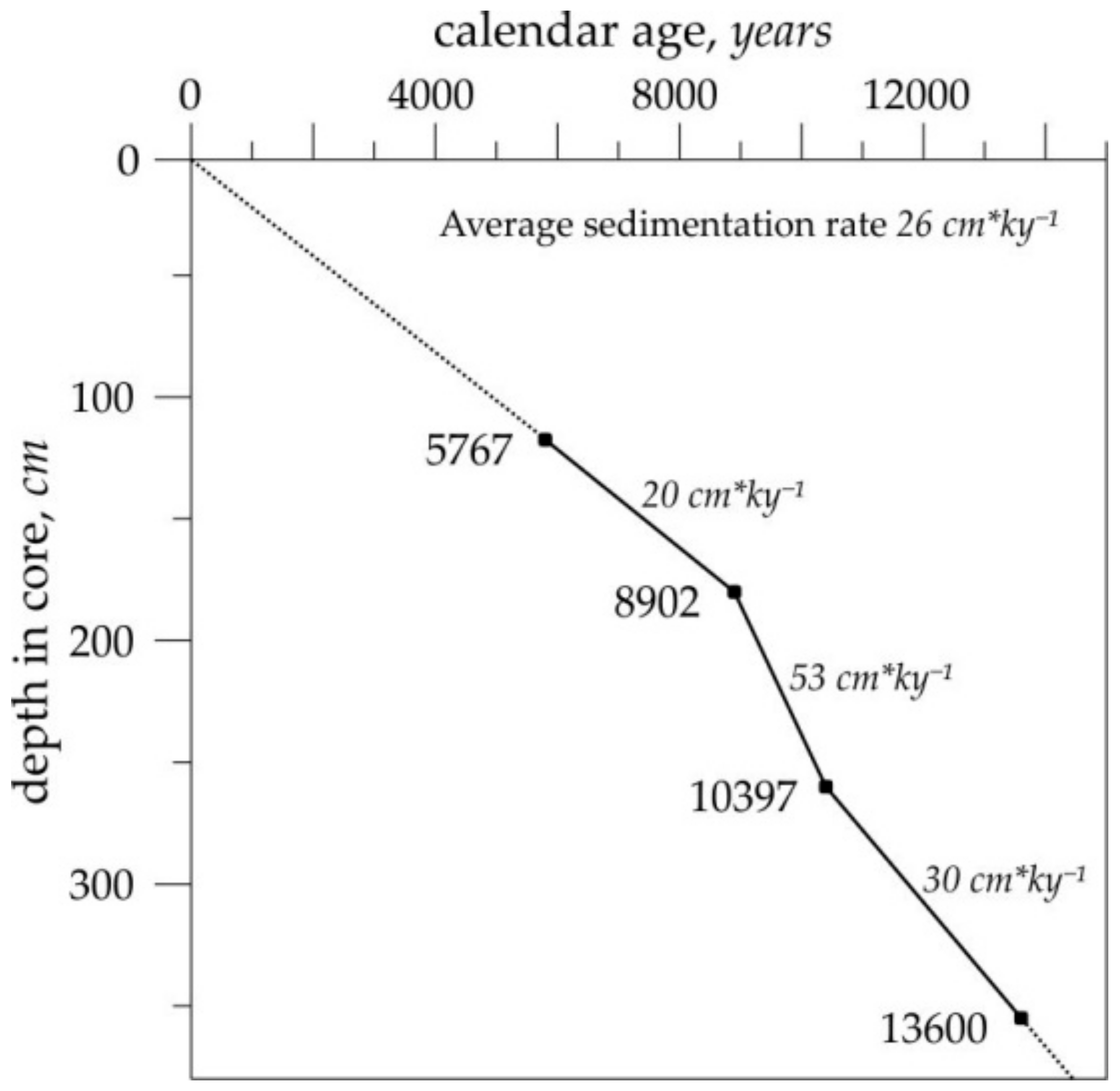
2. Material and Methods
3. Results and Discussion
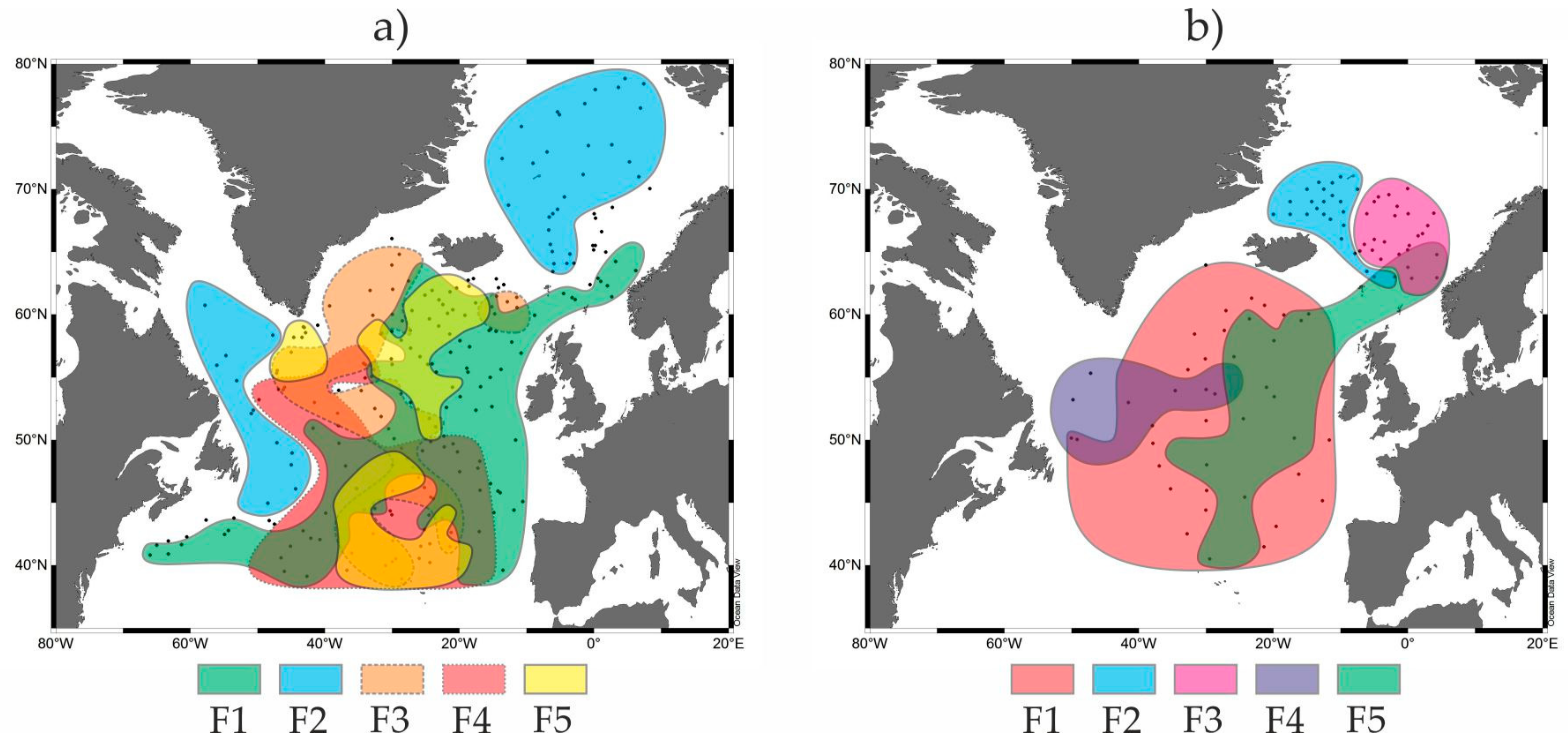
3.1. Modern Distribution of the Radiolarian and Planktic Foraminiferal Factors (=Assemblages) as a Base for the Paleotemperature Estimates
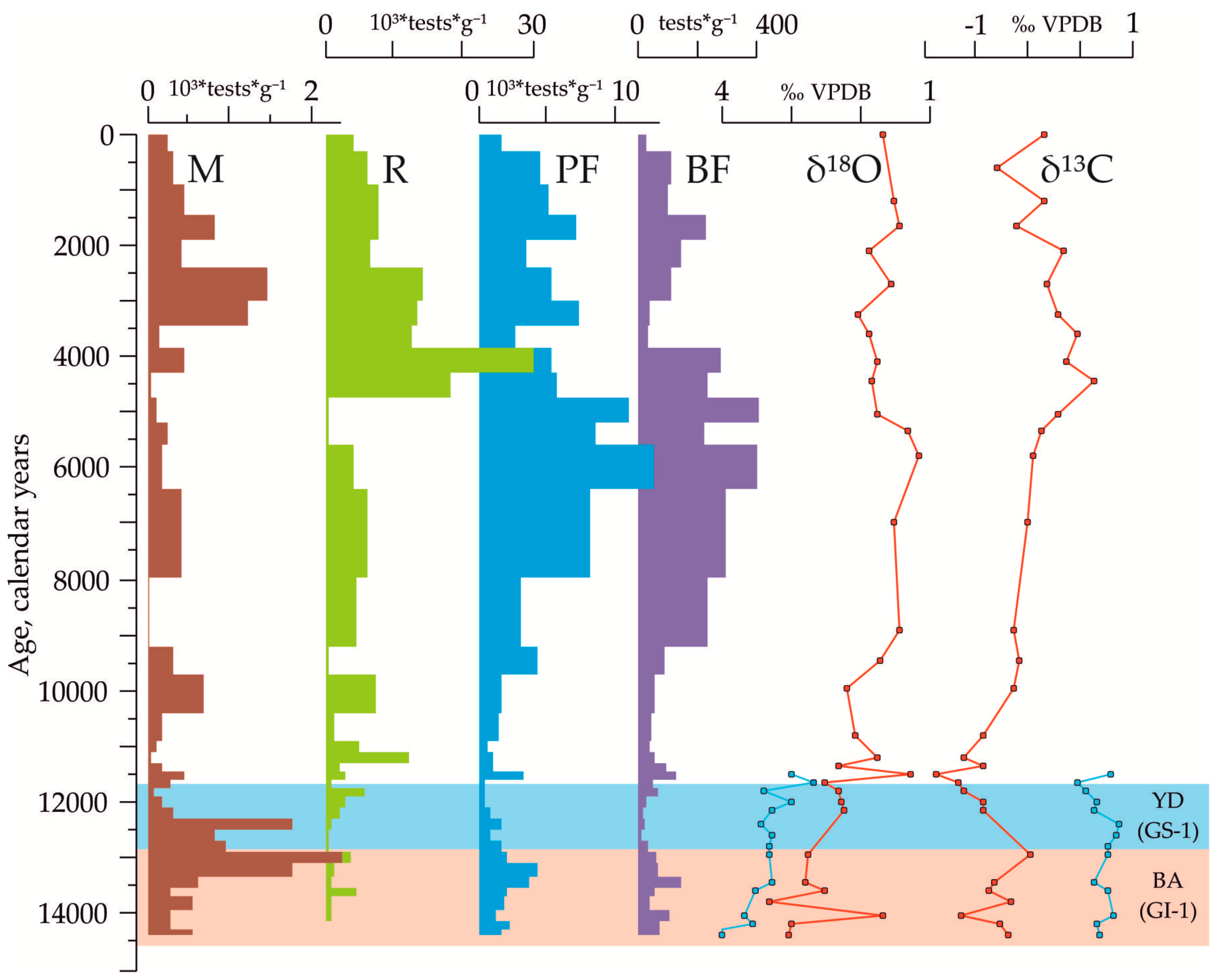
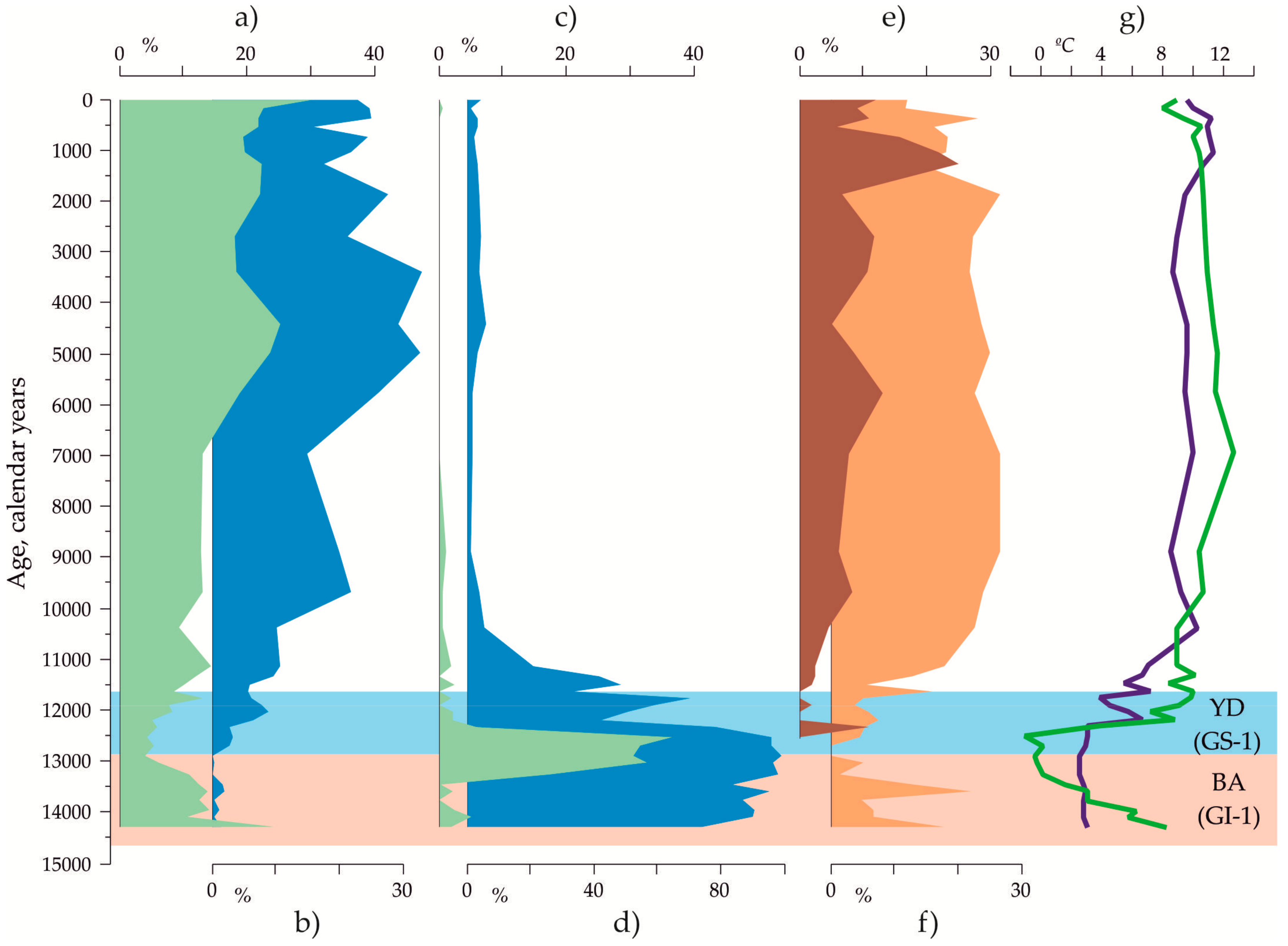
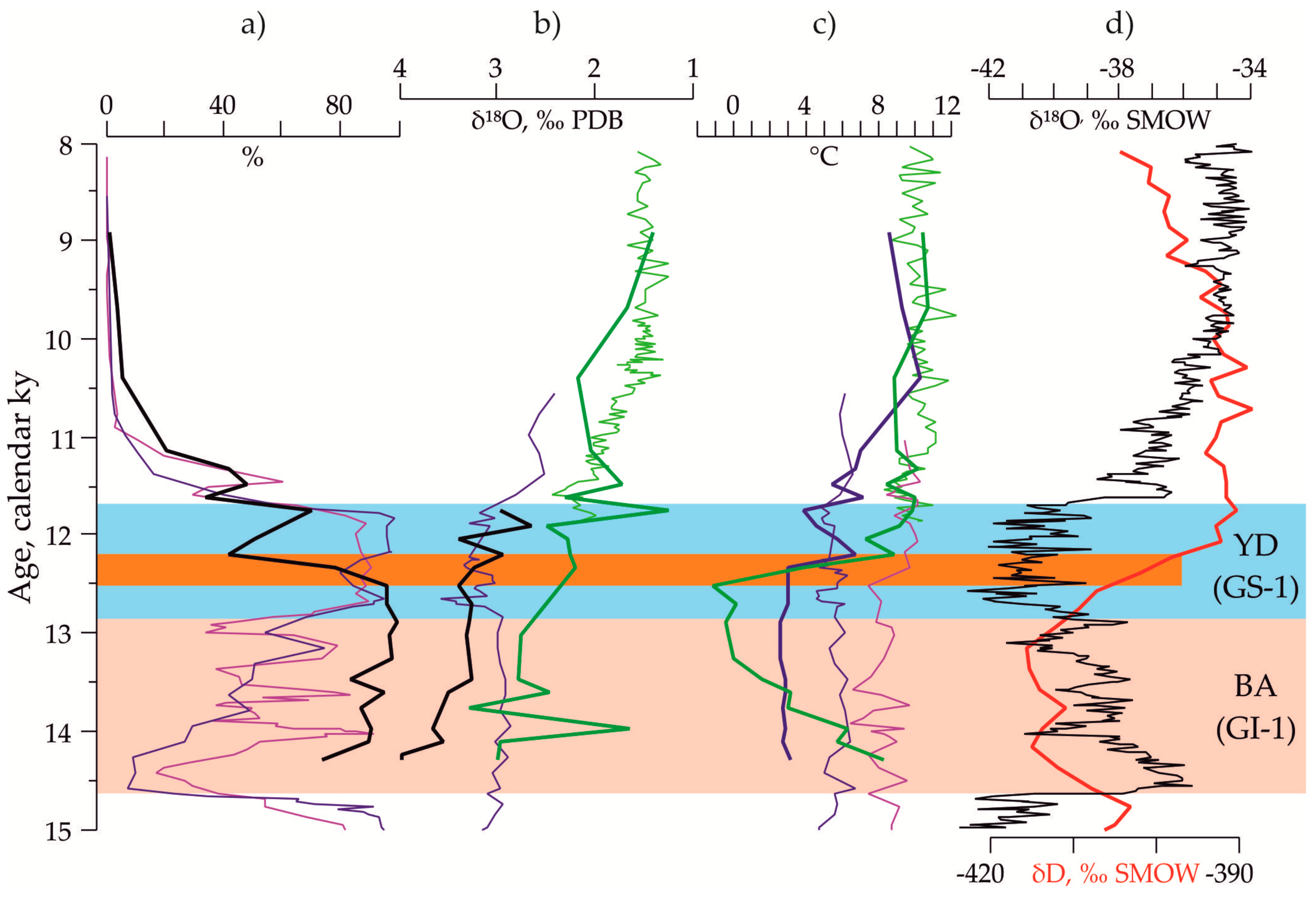
3.2. Indications of the Warm and Cold-Water Events in the Down-Core Records Covering Termination I
3.3. The Stable Oxygen and Carbon Isotope Records in the Core AMK-340
3.4. Paleotemperature Estimates for the Core AMK-340 and Correlation with Other Paleoclimatic Archives
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Delworth, T.L.; Zeng, F. The impact of the North Atlantic oscillation on climate through its influence on the Atlantic meridional overturning circulation. J. Clim. 2016, 29, 941–962. [Google Scholar] [CrossRef]
- Löfverström, M.; Lora, J.M. Abrupt regime shifts in the North Atlantic atmospheric circulation over the last deglaciation. Geophys. Res. Lett. 2017, 44, 8047–8055. [Google Scholar] [CrossRef]
- Knorr, G.; Lohmann, G. Rapid transitions in the Atlantic thermohaline circulation triggered by global warming and meltwater during the last deglaciation. Geochem. Geophys. Geosyst. 2007, 8, Q12006. [Google Scholar] [CrossRef]
- Overpeck, J.T.; Peterson, L.C.; Kipp, N.; Rind, D. Climate change in the circum-North Atlantic region during the last deglaciation. Nature 1989, 338, 553–557. [Google Scholar] [CrossRef]
- Bauch, H.A.; Kandiano, E.S.; Helmke, J.P. Contrasting ocean changes between the subpolar and polar North Atlantic during the past 135 ka. Geophys. Res. Lett. 2012, 39, L11604. [Google Scholar] [CrossRef]
- Matul, A.G. On the problem of paleoceanological evolution of the Reykjanes Ridge region (North Atlantic) during the last deglaciation based on a study of radiolaria. Oceanology 1995, 34, 806–814. [Google Scholar]
- Matul, A.G.; Yushina, I.G. Radiolarians in North Atlantic sediments. Ber. Polarforsch. 1999, 306, 35–45. [Google Scholar]
- Sieger, R.; Grobe, H. PanTool—A Swiss Army Knife for Data Conversion and Recalculation. Available online: https://doi.org/10.1594/PANGAEA.787549 (accessed on 31 July 2018).
- Krauss, W. Currents and mixing in the Irminger Sea and in the Iceland Basin. J. Geophys. Res. 1995, 100, 10851–10871. [Google Scholar] [CrossRef]
- Våge, K.; Pickart, R.S.; Sarafanov, A.; Knutsen, Ø.; Mercier, H.; Lherminier, P.; van Aken, H.M.; Meincke, J.; Quadfasel, D.; Bacon, S. The Irminger Gyre: Circulation, convection, and interannual variability. Deep-Sea Res. Part I 2011, 58, 590–614. [Google Scholar] [CrossRef]
- Locarnini, R.A.; Mishonov, A.V.; Antonov, J.I.; Boyer, T.P.; Garcia, H.E.; Baranova, O.K.; Zweng, M.M.; Paver, C.R.; Reagan, J.R.; Johnson, D.R.; et al. World Ocean Atlas 2013. NOAA, U.S. Department of Commerce: Washington, DC, USA, 2013; NOAA Atlas NESDIS 73, Volume 1: Temperature. Available online: https://www.nodc.noaa.gov/OC5/woa13 (accessed on 31 July 2018).
- Weinelt, M.; Vogelsang, E.; Kucera, M.; Pflaumann, U.; Sarnthein, M.; Voelker, A.; Erlenkeuser, H.; Malmgren, B.A. Variability of North Atlantic heat transfer during MIS 2. Paleoceanography 2003, 18, 1071. [Google Scholar] [CrossRef]
- Thornalley, D.J.R.; McCave, I.N.; Elderfield, H. Freshwater input and abrupt deglacial climate change in the North Atlantic. Paleoceanography 2010, 25, PA1201. [Google Scholar] [CrossRef]
- Thornalley, D.J.R.; Elderfield, H.; Mc Cave, I.N. Reconstructing North Atlantic deglacial surface hydrography and its link to the Atlantic overturning circulation. Global Planet. Chang. 2011, 79, 163–175. [Google Scholar] [CrossRef]
- Zweng, M.M.; Reagan, J.R.; Antonov, J.I.; Locarnini, R.A.; Mishonov, A.V.; Boyer, T.P.; Garcia, H.E.; Baranova, O.K.; Johnson, D.R.; Seidov, D.; et al. World Ocean Atlas 2013. NOAA, Department of Commerce: Washington, DC, USA, 2013; NOAA Atlas NESDIS 74, Volume 2: Salinity. Available online: https://www.nodc.noaa.gov/OC5/woa13 (accessed on 31 July 2018).
- Stuiver, M.; Reimer, P.J.; Reimer, R.W. CALIB 7.1 [WWW program], 2018. Available online: http://calib.org (accessed on 31 July 2018).
- Rasmussen, S.O.; Bigler, M.; Blockley, S.P.; Blunier, T.; Buchardt, S.L.; Clausen, H.B.; Cvijanovic, I.; Dahl-Jensen, D.; Johnsen, S.J.; Fischer, H.; et al. A stratigraphic framework for abrupt climatic changes during the Last Glacial period based on three synchronized Greenland ice-core records: Refining and extending the INTIMATE event stratigraphy. Quat. Sci. Rev. 2014, 106, 14–28. [Google Scholar] [CrossRef]
- Khusid, T.A.; Matul, A.; Barash, M.S.; Gablina, I.F. Benthic foraminiferal records of the last deglaciation on the Reykjanes Ridge. J. Ocean. Res. 2018. submitted for publication. [Google Scholar]
- Gooday, A.J.; Lambshead, P.J.D. Influence of seasonally deposited phytodetritus on benthic foraminiferal populations in the bathyal northeast Atlantic: The species response. Mar. Ecol. Prog. Ser. 1989, 58, 53–67. [Google Scholar] [CrossRef]
- Suhr, S.B.; Pond, D.; Gooday, A.J.; Smith, C.R. Selective feeding by benthic foraminifera on phytodetritus on the western Antarctic Peninsula shelf: Evidence from fatty acid biomarker analysis. Mar. Ecol. Prog. Ser. 2003, 262, 153–162. [Google Scholar] [CrossRef]
- Matul, A.; Mohan, R. Distribution of polycystine radiolarians in bottom surface sediments and its relation to summer sea temperature in the high-latitude North Atlantic. Front. Mar. Sci. 2017, 4, 330. [Google Scholar] [CrossRef]
- Index of /Pub/Data/Paleo/Paleocean/Fossil_Plankton/Russian_Fossil_Plankton. Available online: https://www1.ncdc.noaa.gov/pub/data/paleo/paleocean/fossil_plankton/russian_fossil_plankton (accessed on 31 July 2018).
- Pflaumann, U.; Duprat, J.M.; Pujol, C.; Labeyrie, L.D. Distribution of Planktic Foraminifera in Surface Sediments of the Atlantic Ocean. 1996. Available online: https://doi.org/10.1594/PANGAEA.51621 (accessed on 31 July 2018).
- Zas’ko, D.N. Quantitative distribution of radiolarians in the North Atlantic plankton. Oceanology 2001, 41, 86–93. [Google Scholar]
- Rebotim, A.; Voelker, A.H.L.; Jonkers, L.; Waniek, J.J.; Meggers, H.; Schiebel, R.; Fraile, I.; Schulz, M.; Kucera, M. Factors controlling the depth habitat of planktonic foraminifera in the subtropical eastern North Atlantic. Biogeosciences 2017, 14, 827–859. [Google Scholar] [CrossRef]
- Colebrook, J.M. Continuous plankton records: Seasonal variations in the distribution and abundance of plankton in the North Atlantic Ocean and the North Sea. J. Plankton Res. 1982, 4, 435–462. [Google Scholar] [CrossRef]
- Parsons, T.R.; Takahashi, M.; Hargrave, B. Biological Oceanographic Processes, 3rd ed.; Pergamon Press: Oxford, UK, 1984. [Google Scholar]
- Zielinski, U.; Gersonde, R.; Sieger, R.; Fütterer, D.K. Quaternary surface water temperature estimations: Calibration of a diatom transfer function for the Southern Ocean. Paleoceanography 1998, 13, 365–383. [Google Scholar] [CrossRef]
- Zielinski, U. Quantitative estimation of palaeoenvironment the Antarctic surface water in the late transfer functions with diatoms. Ber. Polarforsch. 1993, 126, 1–148. [Google Scholar]
- Imbrie, J.; Kipp, N.G. A new micropaleontological method for paleoclimatology: Application to a Late Pleistocene Caribbean core. In The Late Cenozoic Glacial Ages; Turekian, K.K., Ed.; Yale University Press: New Haven, CT, USA, 1971; pp. 71–181. [Google Scholar]
- Costello, M.J.; Tsai, P.; Wong, P.S.; Cheung, A.K.L.; Basher, Z.; Chaudhary, C. Marine biogeographic realms and species endemicity. Nat. Commun. 2017, 8, 1057. [Google Scholar] [CrossRef] [PubMed]
- Boltovskoy, D.; Correa, N. Biogeography of Radiolaria Polycystina (Protista) in the World Ocean. Prog. Oceanogr. 2016, 149, 82–105. [Google Scholar] [CrossRef]
- Boltovskoy, D. Foraminifers (Planktonic). In Encyclopedia of Marine Geosciences; Harff, J., Mesched, M., Petersen, S., Thiede, J., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 2014; pp. 1–9. [Google Scholar]
- Kipp, N.G. New transfer function for estimating past sea-surface conditions from sea-bed distribution of planktonic foraminiferal assemblages in the North Atlantic. Mem. Geol. Soc. Am. 1976, 145, 9–42. [Google Scholar]
- Bjørklund, K.R.; Cortese, G.; Swanberg, N.R.; Schrader, H.J. Radiolarian faunal provinces in surface sediments of the Greenland, Iceland and Norwegian (GIN) Seas. Mar. Micropaleontol. 1998, 35, 105–140. [Google Scholar] [CrossRef]
- Clark, P.U.; Shakun, J.D.; Baker, P.A.; Bartlein, P.J.; Brewer, S.; Brook, E.; Carlson, A.E.; Cheng, H.; Kaufman, D.S.; Liu, Z.; et al. Global climate evolution during the last deglaciation. Proc. Natl. Acad. Sci. USA 2012, 109, E1134–E1142. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, K.R.; Hatakeda, K.; Kruglikova, S.B.; Matul, A.G. Amphimelissa setosa (Cleve) (Polycystina, Nassellaria)—A stratigraphic and paleoecological marker of migrating polar environments in the northern hemisphere during the Quaternary. Stratigraphy 2015, 12, 23–37. [Google Scholar]
- Hillaire-Marcel, C.; de Vernal, A. 2008. Stable isotope clue to episodic sea ice formation in the glacial North Atlantic. Earth Planet. Sci. Let. 2008, 268, 143–150. [Google Scholar] [CrossRef]
- Andrews, J.; Jennings, A.E.; Kerwin, M.; Kirby, M.; Manley, W.; Miller, G.H.; Bond, G.; Mac Lean, B. A Heinrich-like event, H-0 (DC-0): Source(s) for detrital carbonate in the North Atlantic during the Younger Dryas Chronozone. Paleoceanography 1995, 10, 943–952. [Google Scholar] [CrossRef]
- Alley, R.B.; Clark, P. The deglaciation of the northern hemisphere: A global perspective. Ann. Rev. Earth Planet. Sci. 1999, 27, 149–182. [Google Scholar] [CrossRef]
- Keigwin, L.D.; Jones, G.A. The marine record of deglaciation from the continental margin off Nova Scotia. Paleoceanography 1995, 10, 973–985. [Google Scholar] [CrossRef]
- Jansen, E.; Andersson, C.; Moros, M.; Nisancioglu, K.H.; Nyland, B.F.; Telford, R.J. The Early to Mid-Holocene thermal optimum in the North Atlantic. In Natural Climate Variability and Global Warming: A Holocene Perspective; Battarbee, R.W., Binney, H.A., Eds.; Wiley-Blackwell: Chichester, UK, 2008; pp. 123–137. [Google Scholar]
- Ravelo, A.C.; Hillaire-Marcel, C. The use of oxygen and carbon isotopes of foraminifera in paleoceanography. In Developments in Marine Geology; Hillaire-Marcel, C., de Vernal, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 1, pp. 735–764. [Google Scholar]
- Jonkers, L.; van Heuven, S.; Zahn, R.; Peeters, F.J.C. Seasonal patterns of shell flux, δ18O and δ13C of small and large N. pachyderma (s) and G. bulloides in the subpolar North Atlantic. Paleoceanography 2013, 28, 164–174. [Google Scholar] [CrossRef]
- Frihmat, Y.E.; Hebbeln, D.; EL Jaaidi, B.; Mhammdi, N. Reconstruction of productivity signal and deep-water conditions in Moroccan Atlantic margin (~35° N) from the last glacial to the Holocene. SpringerPlus 2015, 4, 69. [Google Scholar] [CrossRef] [PubMed]
- NGRIP Members. High-resolution record of Northern Hemisphere climate extending into the last interglacial period. Nature 2004, 431, 147–151. [Google Scholar] [CrossRef] [PubMed]
- EPICA Community Members. Eight glacial cycles from an Antarctic ice core. Nature 2004, 429, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Eynaud, F. Planktonic foraminifera in the Arctic: Potentials and issues regarding modern and Quaternary populations. IOP Conf. Ser. Earth Environ. Sci. 2011, 14, 1–12. [Google Scholar] [CrossRef]
- Pearce, C.; Seidenkrantz, M.-S.; Kuijpers, A.; Masser, G.; Reynisson, N.F.; Kristiansen, S.M. Ocean lead at the termination of the Younger Dryas cold spell. Nat. Commun. 2013, 4, 1664. [Google Scholar] [CrossRef] [PubMed]
- Ebbesen, H.; Hald, M. Unstable Younger Dryas climate in the northeast North Atlantic. Geology 2004, 32, 673–676. [Google Scholar] [CrossRef]
- Rickaby, R.E.M.; Elderfield, H. Evidence from the high-latitude North Atlantic for variations in Antarctic Intermediate water flow during the last deglaciation. Geochem. Geophys. Geosyst. 2005, 6, Q05001. [Google Scholar] [CrossRef]
- Barker, S.; Diz, P.; Vautravers, M.J.; Pike, J.; Knorr, G.; Hall, I.R.; Broecker, W.S. Interhemispheric Atlantic seesaw response during the last deglaciation. Nature 2009, 457, 1097–1102. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth in Core | 14C-Datings, Years | Calendar Age, Years | Dating Code |
|---|---|---|---|
| 118 cm | 5480 ± 50 | 5767 | KIA4187 |
| 181 cm | 8030 ± 60 | 8902 | KIA4188 |
| 260 cm | 9220 ± 60 | 10,397 | KIA4189 |
| 356 cm | 11,830 ± 70 | 13,600 | KIA4190 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matul, A.; Barash, M.S.; Khusid, T.A.; Behera, P.; Tiwari, M. Paleoenvironment Variability during Termination I at the Reykjanes Ridge, North Atlantic. Geosciences 2018, 8, 375. https://doi.org/10.3390/geosciences8100375
Matul A, Barash MS, Khusid TA, Behera P, Tiwari M. Paleoenvironment Variability during Termination I at the Reykjanes Ridge, North Atlantic. Geosciences. 2018; 8(10):375. https://doi.org/10.3390/geosciences8100375
Chicago/Turabian StyleMatul, Alexander, Max S. Barash, Tatyana A. Khusid, Padmasini Behera, and Manish Tiwari. 2018. "Paleoenvironment Variability during Termination I at the Reykjanes Ridge, North Atlantic" Geosciences 8, no. 10: 375. https://doi.org/10.3390/geosciences8100375
APA StyleMatul, A., Barash, M. S., Khusid, T. A., Behera, P., & Tiwari, M. (2018). Paleoenvironment Variability during Termination I at the Reykjanes Ridge, North Atlantic. Geosciences, 8(10), 375. https://doi.org/10.3390/geosciences8100375