
Methanogenesis from Mineral Carbonates, a Potential Indicator for Life on Mars

 ,
,  and
and
Abstract
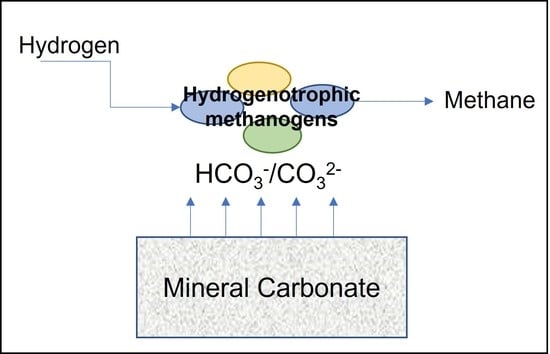
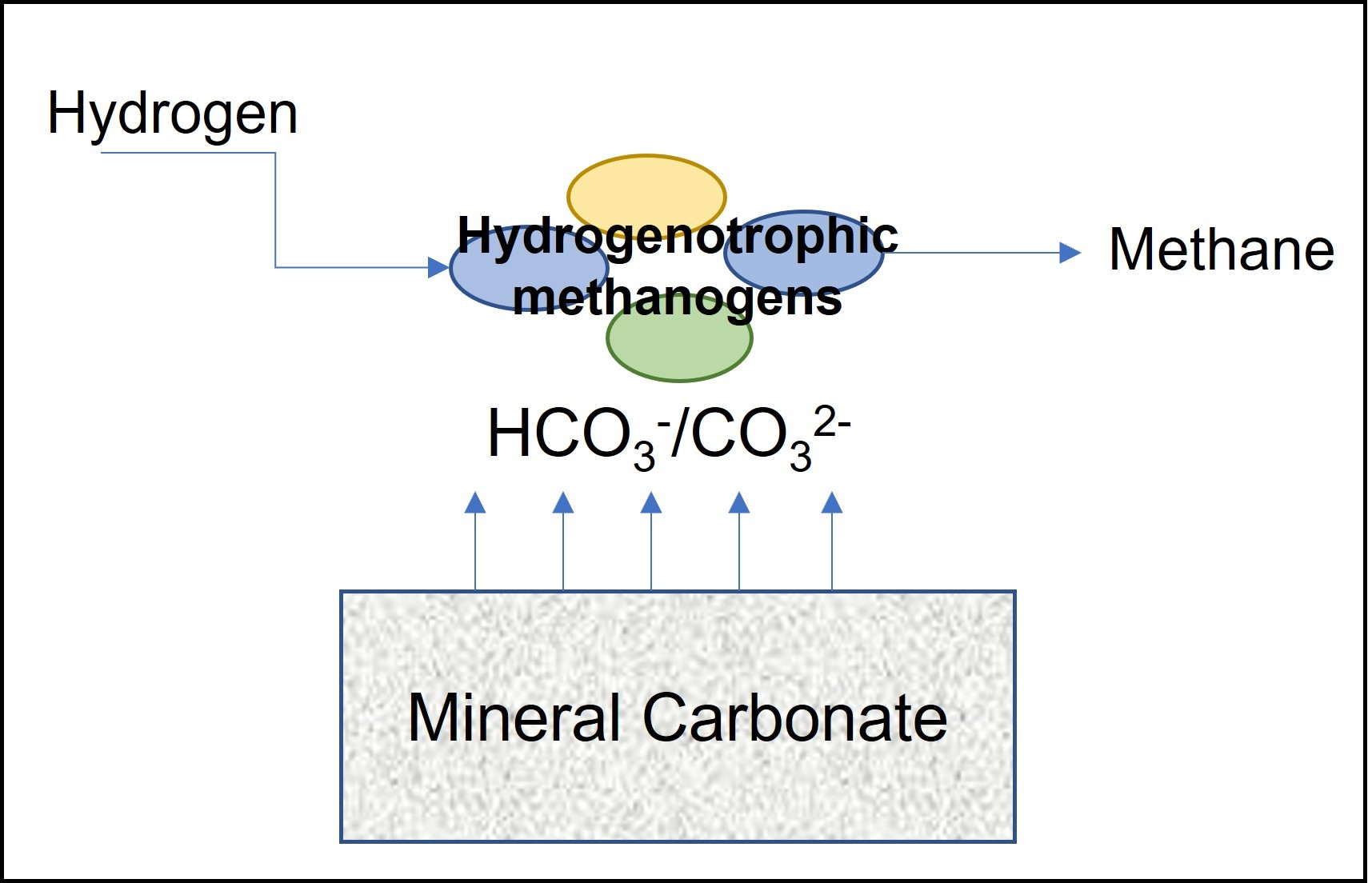{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Release of Dissolved Inorganic Carbon from Carbonate Sources
2.2. Sampling Sites and Preparation of Enrichment Cultures
2.3. Reaction Set-Up and Analysis
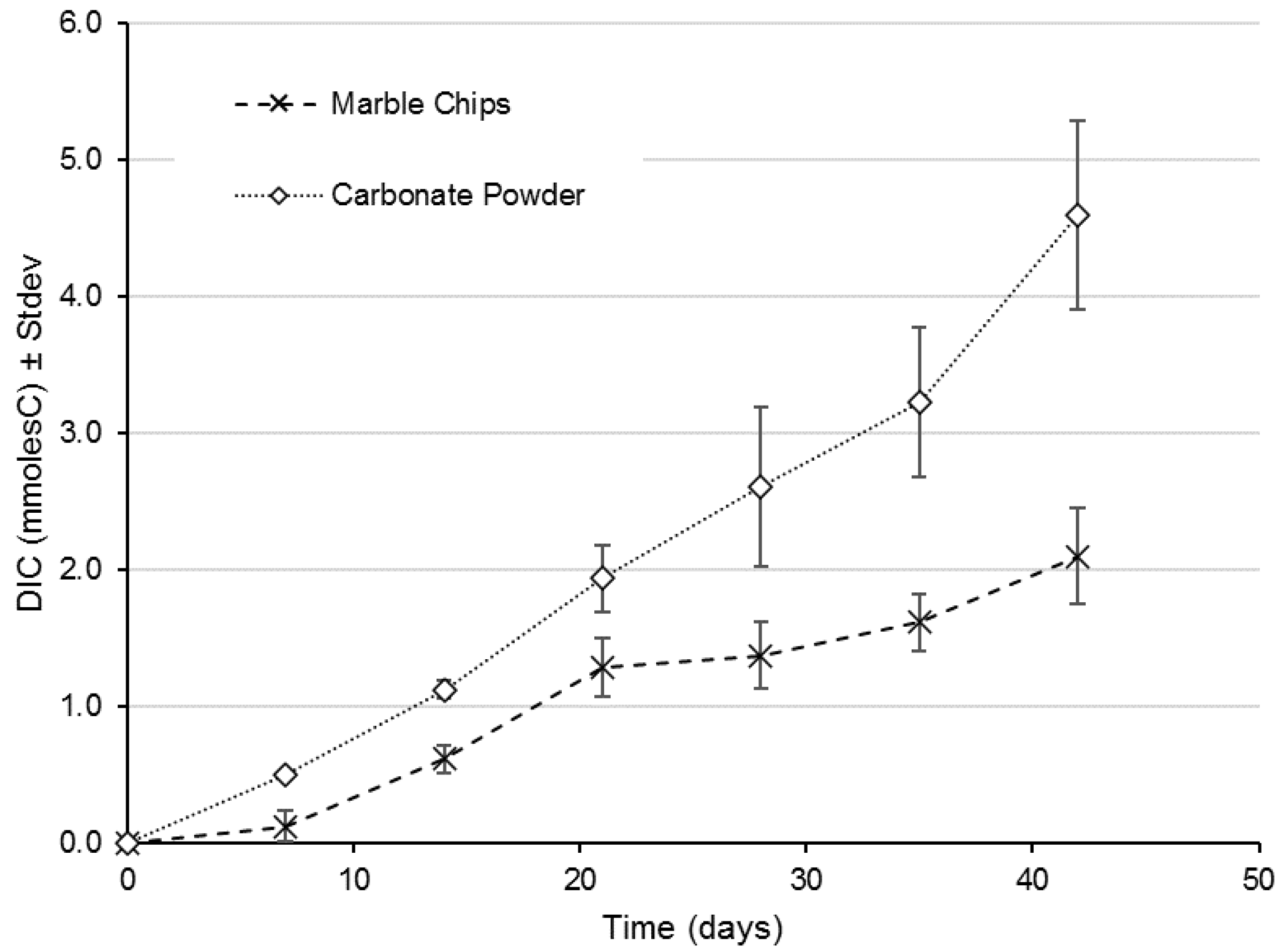
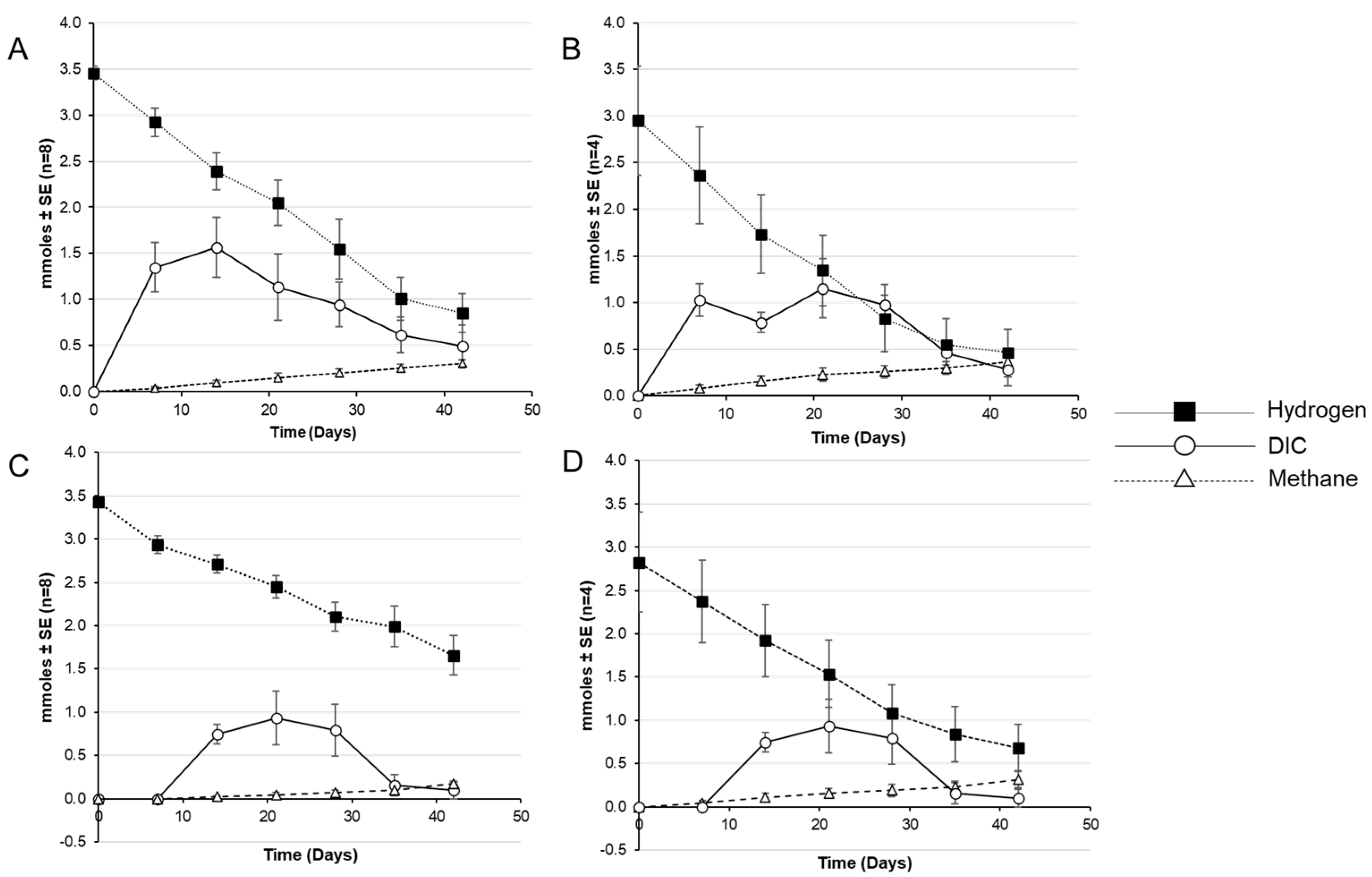
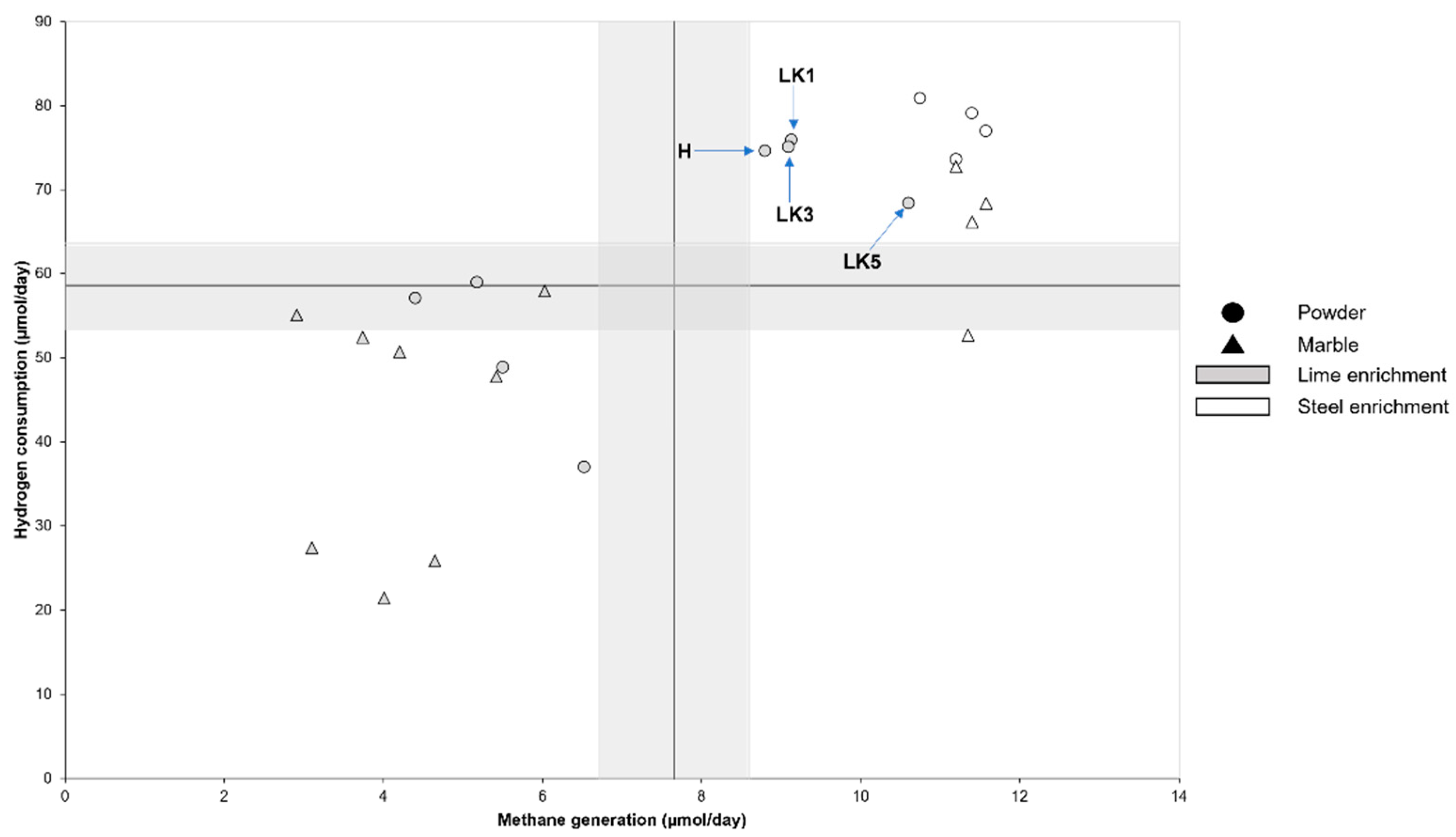
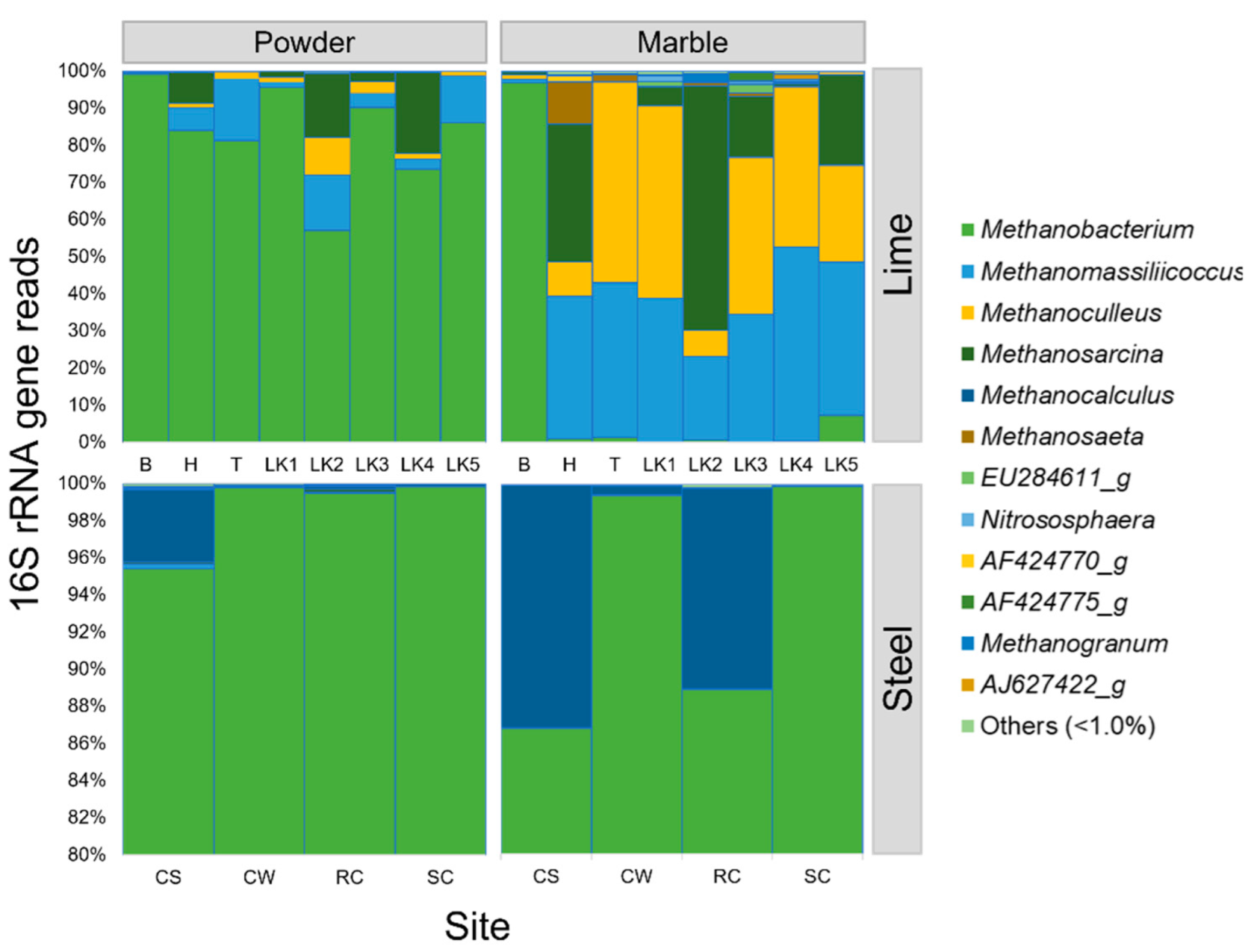
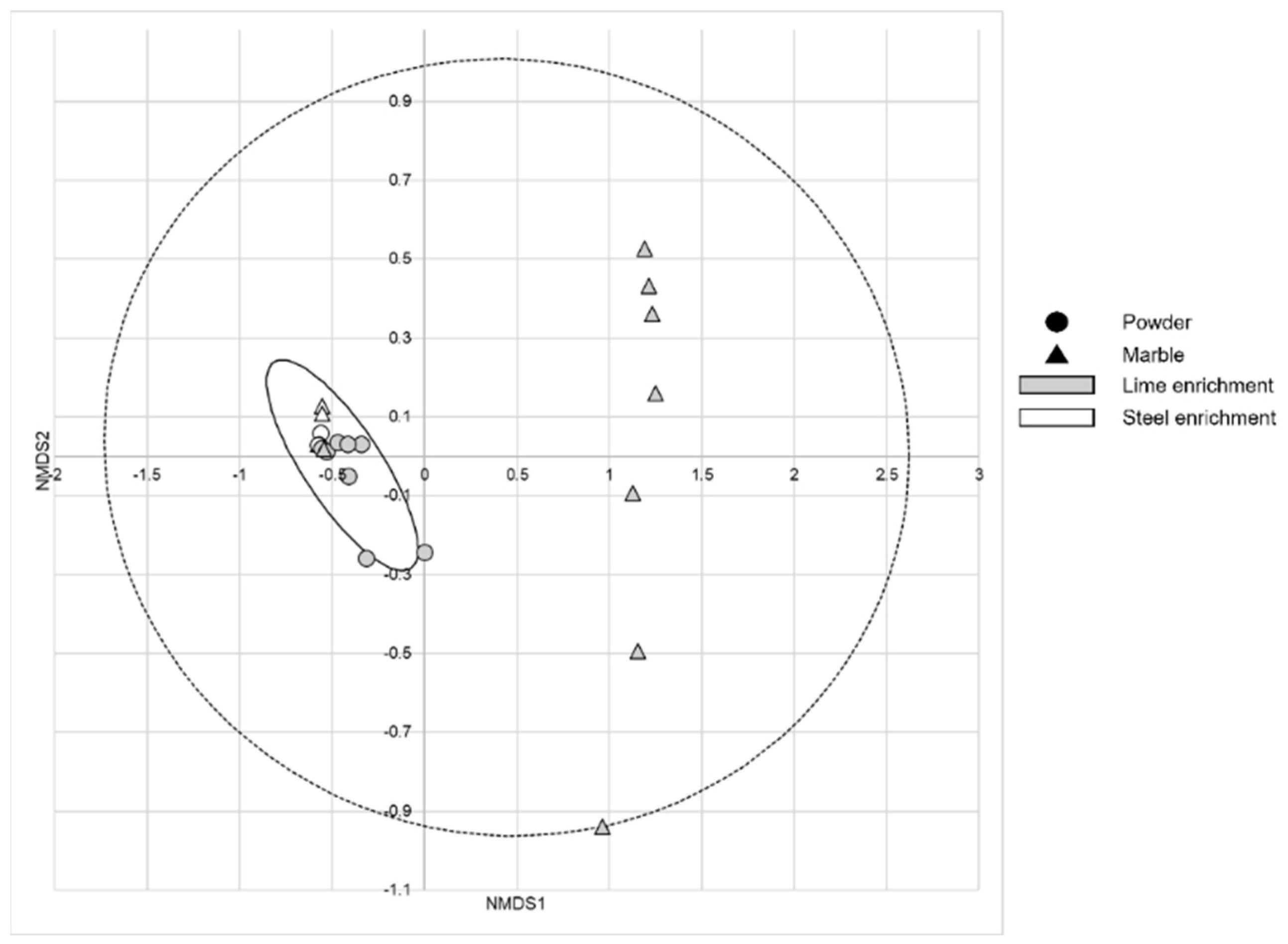
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NASA. NASA Astrobiology Strategy 2015. Edited by L.Hays 2015. Available online: https://nai.nasa.gov/media/medialibrary/2015/10/NASA_Astrobiology_Strategy_2015_151008.pdf (accessed on 8 November 2021).
- Grenfell, J.L. A review of exoplanetary biosignatures. Phys. Rep. 2017, 713, 1–17. [Google Scholar] [CrossRef]
- Williford, K.H.; Farley, K.A.; Stack, K.M.; Allwood, A.C.; Beaty, D.; Beegle, L.W.; Bhartia, R.; Brown, A.J.; de la Torre Juarez, M.; Hamran, S.-E. The NASA Mars 2020 rover mission and the search for extraterrestrial life. In From Habitability to Life on Mars; Elsevier: Amsterdam, The Netherlands, 2018; pp. 275–308. [Google Scholar]
- Banerdt, W.B.; Smrekar, S.E.; Banfield, D.; Giardini, D.; Golombek, M.; Johnson, C.L.; Lognonné, P.; Spiga, A.; Spohn, T.; Perrin, C. Initial results from the InSight mission on Mars. Nat. Geosci. 2020, 13, 183–189. [Google Scholar] [CrossRef]
- Vago, J.L.; Westall, F.; Coates, A.J.; Jaumann, R.; Korablev, O.; Ciarletti, V.; Mitrofanov, I.; Josset, J.-L.; De Sanctis, M.C.; Bibring, J.-P. Habitability on early Mars and the search for biosignatures with the ExoMars Rover. Astrobiology 2017, 17, 471–510. [Google Scholar] [CrossRef] [PubMed]
- Webster, C.R.; Mahaffy, P.R.; Atreya, S.K.; Moores, J.E.; Flesch, G.J.; Malespin, C.; McKay, C.P.; Martinez, G.; Smith, C.L.; Martin-Torres, J.; et al. Background levels of methane in Mars’ atmosphere show strong seasonal variations. Science 2018, 360, 1093–1096. [Google Scholar] [CrossRef]
- Oehler, D.Z.; Etiope, G. Methane Seepage on Mars: Where to Look and Why. Astrobiology 2017, 17, 1233–1264. [Google Scholar] [CrossRef] [PubMed]
- Ferry, J.G. How to Make a Living by Exhaling Methane. Annu. Rev. Biochem. 2010, 64, 453–473. [Google Scholar] [CrossRef]
- Nixon, S.L.; Cousins, C.R.; Cockell, C.S. Plausible microbial metabolisms on Mars: Microbial metabolisms on Mars. Astron. Geophys. 2013, 54, 1.13–1.16. [Google Scholar] [CrossRef]
- Holm, N.G.; Oze, C.; Mousis, O.; Waite, J.H.; Guilbert-Lepoutre, A. Serpentinization and the Formation of H2 and CH4 on Celestial Bodies (Planets, Moons, Comets). Astrobiology 2015, 15, 587–600. [Google Scholar] [CrossRef]
- McMahon, S.; Parnell, J.; Blamey, N.J.F. Evidence for Seismogenic Hydrogen Gas, a Potential Microbial Energy Source on Earth and Mars. Astrobiology 2016, 16, 690–702. [Google Scholar] [CrossRef]
- Blank, J.G.; Green, S.J.; Blake, D.; Valley, J.W.; Kita, N.T.; Treiman, A.; Dobson, P.F. An alkaline spring system within the Del Puerto Ophiolite (California, USA): A Mars analog site. Planet. Space Sci. 2009, 57, 533–540. [Google Scholar] [CrossRef]
- Etiope, G.; Ehlmann, B.L.; Schoell, M. Low temperature production and exhalation of methane from serpentinized rocks on Earth: A potential analog for methane production on Mars. Icarus 2013, 224, 276–285. [Google Scholar] [CrossRef]
- Alexander, W.R.; Milodowski, A.E. Cyprus Natural Analogue Project (CNAP) Phase IV Final Report; Nuclear Decommissioning Agency: Harwell, UK, 2015. [Google Scholar]
- D’Alessandro, W.; Daskalopoulou, K.; Calabrese, S.; Bellomo, S. Water chemistry and abiogenic methane content of a hyperalkaline spring related to serpentinization in the Argolida ophiolite (Ermioni, Greece). Mar. Pet. Geol. 2018, 89, 185–193. [Google Scholar] [CrossRef]
- Ramirez, R.M.; Craddock, R.A. The geological and climatological case for a warmer and wetter early Mars. Nat. Geosci. 2018, 11, 230–237. [Google Scholar] [CrossRef]
- Hu, R.; Kass, D.M.; Ehlmann, B.L.; Yung, Y.L. Tracing the fate of carbon and the atmospheric evolution of Mars. Nat. Commun. 2015, 6, 10003. [Google Scholar] [CrossRef] [PubMed]
- Horgan, B.H.N.; Anderson, R.B.; Dromart, G.; Amador, E.S.; Rice, M.S. The mineral diversity of Jezero crater: Evidence for possible lacustrine carbonates on Mars. Icarus 2020, 339, 113526. [Google Scholar] [CrossRef]
- Burke, I.; Mortimer, R.; Palani, S.; Whittleston, R.; Lockwood, C.; Ashley, D.; Stewart, D. Biogeochemical reduction processes in a hyper-alkaline affected leachate soil profile. Geomicrobiol. J. 2012, 29, 769–779. [Google Scholar] [CrossRef]
- Wormald, R.M.; Rout, S.P.; Mayes, W.; Gomes, H.; Humphreys, P.N. Hydrogenotrophic Methanogenesis under Alkaline Conditions. Front. Microbiol. 2020, 11, 614227. [Google Scholar] [CrossRef]
- Rout, S.P.; Radford, J.; Laws, A.P.; Sweeney, F.; Elmekawy, A.; Gillie, L.J.; Humphreys, P.N. Biodegradation of the Alkaline Cellulose Degradation Products Generated during Radioactive Waste Disposal. PLoS ONE 2014, 9, e107433. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Whiteley, A.S.; O’Donnell, A.G.; Bailey, M.J. Rapid Method for Coextraction of DNA and RNA from Natural Environments for Analysis of Ribosomal DNA- and rRNA-Based Microbial Community Composition. Appl. Environ. Microbiol. 2000, 66, 5488–5491. [Google Scholar] [CrossRef]
- Kral, T.A.; Birch, W.; Lavender, L.E.; Virden, B.T. Potential use of highly insoluble carbonates as carbon sources by methanogens in the subsurface of Mars. Planet. Space Sci. 2014, 101, 181–185. [Google Scholar] [CrossRef]
- Phillips-Lander, C.M.; Parnell, S.R.; McGraw, L.E.; Madden, M.E.E. Carbonate dissolution rates in high salinity brines: Implications for post-Noachian chemical weathering on Mars. Icarus 2018, 307, 281–293. [Google Scholar] [CrossRef]
- Zastro, A.M.; Glotch, T.D. Distinct carbonate lithologies in Jezero crater, Mars. Geophys. Res. Lett. 2021, 48, 9. [Google Scholar] [CrossRef] [PubMed]
- Siegert, M.; Li, X.-F.; Yates, M.D.; Logan, B.E. The presence of hydrogenotrophic methanogens in the inoculum improves methane gas production in microbial electrolysis cells. Front. Microbiol. 2015, 5, 778. [Google Scholar] [CrossRef] [PubMed]
- Maus, I.; Wibberg, D.; Stantscheff, R.; Cibis, K.; Eikmeyer, F.-G.; König, H.; Pühler, A.; Schlüter, A. Complete genome sequence of the hydrogenotrophic archaeon Methanobacterium sp. Mb1 isolated from a production-scale biogas plant. J. Biotechnol. 2013, 168, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Chen, H.; Yu, B.; He, J.; Zheng, P.; Mao, X.; Tian, G.; Yu, J.; Huang, Z.; Luo, J. Dietary pea fiber increases diversity of colonic methanogens of pigs with a shift from Methanobrevibacter to Methanomassiliicoccus-like genus and change in numbers of three hydrogenotrophs. BMC Microbiol. 2017, 17, 17. [Google Scholar] [CrossRef]
- Maus, I.; Wibberg, D.; Stantscheff, R.; Eikmeyer, F.-G.; Seffner, A.; Boelter, J.; Szczepanowski, R.; Blom, J.; Jaenicke, S.; König, H. Complete genome sequence of the hydrogenotrophic, methanogenic archaeon Methanoculleus bourgensis strain MS2T, isolated from a sewage sludge digester. Am. Soc. Microbiol. 2012, 194, 19. [Google Scholar]
- Lambie, S.C.; Kelly, W.J.; Leahy, S.C.; Li, D.; Reilly, K.; McAllister, T.A.; Valle, E.R.; Attwood, G.T.; Altermann, E. The complete genome sequence of the rumen methanogen Methanosarcina barkeri CM1. Stand. Genom. Sci. 2015, 10, 57. [Google Scholar] [CrossRef]
- Ragsdale, S.W.; Pierce, E. Acetogenesis and the Wood–Ljungdahl pathway of CO2 fixation. Biochim. Biophys. Acta BBA Proteins Proteom. 2008, 1784, 1873–1898. [Google Scholar] [CrossRef]
- Borrel, G.; Adam, P.S.; Gribaldo, S. Methanogenesis and the Wood–Ljungdahl Pathway: An Ancient, Versatile, and Fragile Association. Genome Biol. Evol. 2016, 8, 1706–1711. [Google Scholar] [CrossRef]
- Orange, F.; Westall, F.; Disnar, J.R.; Prieur, D.; Bienvenu, N.; Le Romancer, M.; Défarge, C. Experimental silicification of the extremophilic Archaea Pyrococcus abyssi and Methanocaldococcus jannaschii: Applications in the search for evidence of life in early Earth and extraterrestrial rocks. Geobiology 2009, 7, 403–418. [Google Scholar] [CrossRef]
- Joseph, R.G.; Planchon, O.; Duxbury, N.S.; Latif, K.; Kidron, G.J.; Consorti, L.; Armstrong, R.A.; Gibson, C.; Schild, R. Oceans, Lakes, and Stromatolites on Mars. Adv. Astron. 2020, 2020, 6959532. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wormald, R.M.; Hopwood, J.; Humphreys, P.N.; Mayes, W.; Gomes, H.I.; Rout, S.P. Methanogenesis from Mineral Carbonates, a Potential Indicator for Life on Mars. Geosciences 2022, 12, 138. https://doi.org/10.3390/geosciences12030138
Wormald RM, Hopwood J, Humphreys PN, Mayes W, Gomes HI, Rout SP. Methanogenesis from Mineral Carbonates, a Potential Indicator for Life on Mars. Geosciences. 2022; 12(3):138. https://doi.org/10.3390/geosciences12030138
Chicago/Turabian StyleWormald, Richard M., Jeremy Hopwood, Paul N. Humphreys, William Mayes, Helena I. Gomes, and Simon P. Rout. 2022. "Methanogenesis from Mineral Carbonates, a Potential Indicator for Life on Mars" Geosciences 12, no. 3: 138. https://doi.org/10.3390/geosciences12030138
APA StyleWormald, R. M., Hopwood, J., Humphreys, P. N., Mayes, W., Gomes, H. I., & Rout, S. P. (2022). Methanogenesis from Mineral Carbonates, a Potential Indicator for Life on Mars. Geosciences, 12(3), 138. https://doi.org/10.3390/geosciences12030138