Increased Flight Initiation Distance (FID) in Golden Marmots (Marmota caudata aurea) Responding to Domestic Dogs in A Landscape of Human Disturbance


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
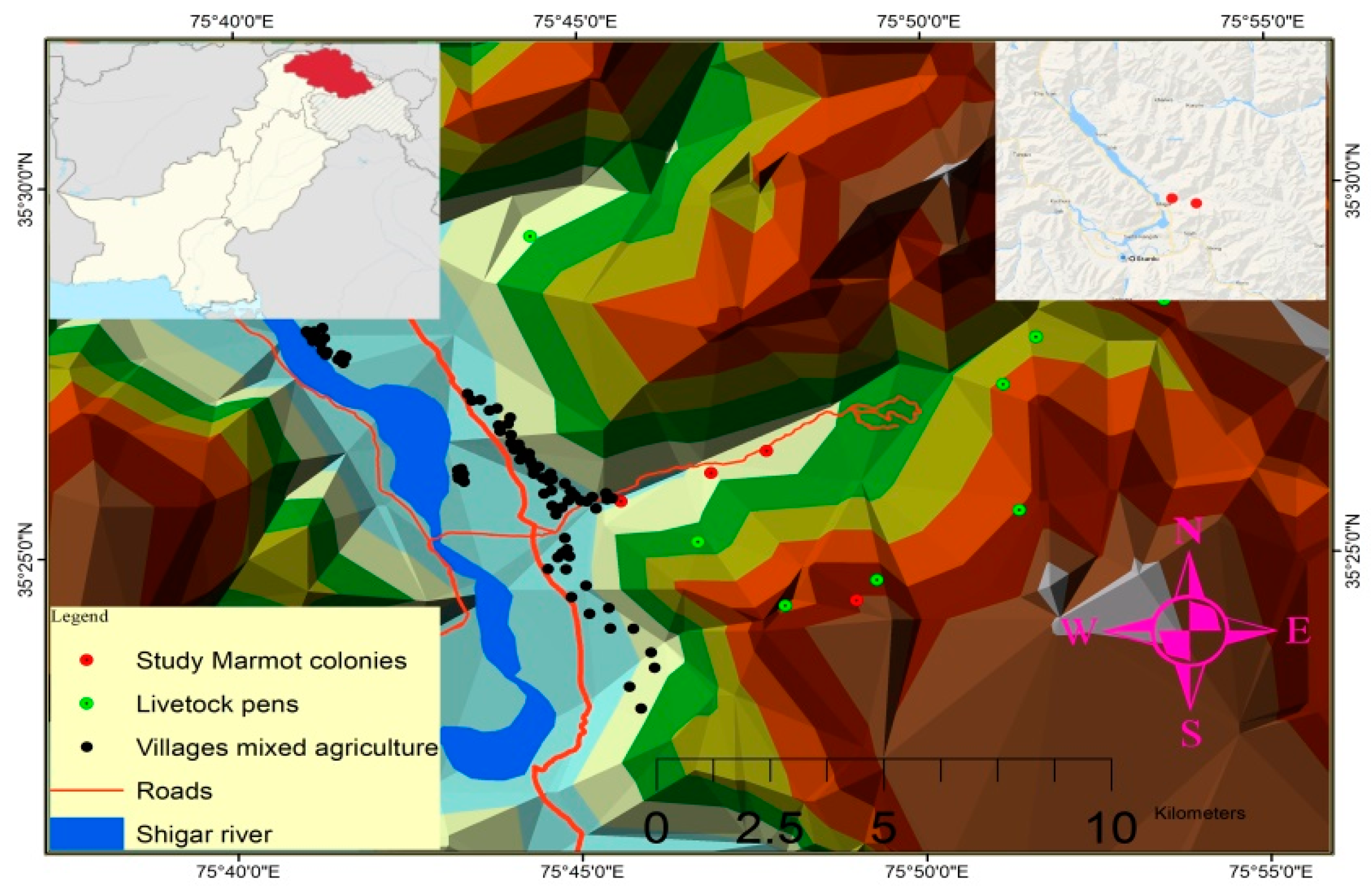
2.1. Study Area
2.2. Live Capture and Marking of Marmots
2.3. Field Identification of Unmarked Individuals
2.4. Assessment of Flight Initiation Distance (FID)
2.5. Data Analyses
2.6. Ethical Approval.
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stankowich, T.; Coss, R.G. Effects of predator behavior and proximity on risk assessment by Columbian black-tailed deer. Behav. Ecol. 2005, 17, 246–254. [Google Scholar] [CrossRef]
- Ydemberg, R.; Dill, L. The economics of fleeing from predators. Adv. Study Behav. 1986, 16, 229–249. [Google Scholar]
- Stankowich, T.; Blumstein, D.T. Fear in animals: a meta-analysis and review of risk assessment. Proc. R. Soc. Lond. B Biol. Sci. 2005, 272, 2627–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frid, A.; Dill, L. Human-caused disturbance stimuli as a form of predation risk. Conserv. Ecol. 2002, 6, 11. [Google Scholar] [CrossRef]
- Li, C.; Monclús, R.; Maul, T.L.; Jiang, Z.; Blumstein, D.T. Quantifying human disturbance on antipredator behavior and flush initiation distance in yellow-bellied marmots. Appl. Anim. Behav. Sci. 2011, 129, 146–152. [Google Scholar] [CrossRef]
- Weston, M.A.; Stankowich, T. Dogs as agents of disturbance. In Free-Ranging Dogs and Wildlife Conservation; Gompper, M.E., Ed.; Oxford University Press: Newyork City, NY, USA, 2013. [Google Scholar]
- Silva-Rodríguez, E.A.; Sieving, K.E. Domestic dogs shape the landscape-scale distribution of a threatened forest ungulate. Biol. Conserv. 2012, 150, 103–110. [Google Scholar] [CrossRef]
- Shannon, G.; Crooks, K.R.; Wittemyer, G.; Fristrup, K.M.; Angeloni, L.M. Road noise causes earlier predator detection and flight response in a free-ranging mammal. Behav. Ecol. 2016, 27, 1370–1375. [Google Scholar] [CrossRef] [Green Version]
- Baskin, L.M.; Hjalten, J. Fright and flight behavior of reindeer. Alces 2001, 37, 435–445. [Google Scholar]
- Kloppers, E.L.; St. Clair, C.C.; Hurd, T.E. Predator-resembling aversive conditioning for managing habituated wildlife. Ecol. Soc. 2005, 10. [Google Scholar] [CrossRef]
- Stankowich, T. Ungulate flight responses to human disturbance: a review and meta-analysis. Biol. Conserv. 2008, 141, 2159–2173. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Hiding time in refuge. In Escaping from Predators: An Integrative View of Escape Decisions; Cambridge University Press: Newyork City, NY, USA, 2015; pp. 227–261. [Google Scholar]
- Marealle, W.N.; Fossøy, F.; Holmern, T.; Stokke, B.G.; Røskaft, E. Does illegal hunting skew Serengeti wildlife sex ratios? Wildl. Biol. 2010, 16, 419–430. [Google Scholar] [CrossRef]
- Thiel, D.; Jenni-Eiermann, S.; Palme, R. Evidence for Physiological Stress in Capercaillie Tetrao Urogallus Due to Human Winter Recreation. Behavioral and Physiological Effects in Capercaillie (Tetrao urogallus) Caused by Human Disturbance. Ph.D. Thesis, University of Zurich, Rämistrasse, Zurich, 2007. [Google Scholar]
- Weston, M.; McLeod, E.M.; Blumstein, D.; Guay, P.-J. A review of flight-initiation distances and their application to managing disturbance to Australian birds. Emu-Austral Ornithol. 2012, 112, 269–286. [Google Scholar] [CrossRef] [Green Version]
- Stankowich, T.; Reimers, E. Escape Decisions in mammals. In Escaping from Predators: An Integrative View of Escape Decisions; Cambridge University Press: London, UK, 2015. [Google Scholar]
- Kenward, R. Hawks and doves: factors affecting success and selection in goshawk attacks on woodpigeons. J. Anim. Ecol. 1978, 449–460. [Google Scholar] [CrossRef]
- Rocha, C.; Bergallo, H. Thermal biology and flight distance of Tropidurus oreadicus (Sauria Iguanidae) in an area of Amazonian Brazil. Ethol. Ecol. Evol. 1990, 2, 263–268. [Google Scholar] [CrossRef]
- Abrahams, M.V. The interaction between antipredator behaviour and antipredator morphology: experiments with fathead minnows and brook sticklebacks. Can. J. Zool. 1995, 73, 2209–2215. [Google Scholar] [CrossRef] [Green Version]
- Matson, K.D.; Cohen, A.A.; Klasing, K.C.; Ricklefs, R.E.; Scheuerlein, A. No simple answers for ecological immunology: relationships among immune indices at the individual level break down at the species level in waterfowl. Proc. R. Soc. B Biol. Sci. 2005, 273, 815–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairbanks, B.; Dobson, F.S. Mechanisms of the group-size effect on vigilance in Columbian ground squirrels: dilution versus detection. Anim. Behav. 2007, 73, 115–123. [Google Scholar] [CrossRef]
- Stankowich, T.; Coss, R.G. Effects of risk assessment, predator behavior, and habitat on escape behavior in Columbian black-tailed deer. Behav. Ecol. 2006, 18, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Louis, S.; Le Beere, M. Adjustment in flight distance from humans by Marmota marmota. Can.J. Zool-Revue Canadienne De Zoologie 2000, 78, 556–563. [Google Scholar] [CrossRef]
- Weterings, M.J.; Zaccaroni, M.; van der Koore, N.; Zijlstra, L.M.; Kuipers, H.J.; van Langevelde, F.; van Wieren, S.E. Strong reactive movement response of the medium-sized European hare to elevated predation risk in short vegetation. Anim. Behav. 2016, 115, 107–114. [Google Scholar] [CrossRef]
- Zhang, H.; Li, W.; Hu, Y.; Zhang, Y. Opposite companion effect on flight initiation distance in sympatric species: plateau pika (Ochotona curzoniae) and White-rumped Snowfinch (Onychostruthus taczanowskii). Can. J. Zool. 2015, 94, 109–114. [Google Scholar] [CrossRef]
- McLean, B.S. Urocitellus parryii (Rodentia: Sciuridae). Mammalian Species 2018, 50, 84–99. [Google Scholar] [CrossRef] [Green Version]
- Holtmeier, F.-K. Animals’ Influence on the Landscape and Ecological Importance; Springer: Berlin, Germany, 2015. [Google Scholar]
- Blumstein, D.T.; Arnold, W. Ecology and social behavior of golden marmots (Marmota caudata aurea). J. Mammal. 1998, 79, 873–886. [Google Scholar] [CrossRef]
- Qureshi, B.U.D. Identification, Population and Food Habits of Marmots in District Neelum, Azad Jammu and Kashmir; Pir Mahal Ali Sha Arid Agriculture University Rawalpindi: Punjab, Pakistan, 2015. [Google Scholar]
- Shabbir, S.; Anwar, M.; Hussain, I.; Nawaz, M. Food habits and diet overlap of two sympatric carnivore species in Chitral, Pakistan. J. Anim. Plants Sci. 2013, 23, 100–106. [Google Scholar]
- Patil, V.; Morrison, S.; Karels, T.; Hik, D. Winter weather versus group thermoregulation: what determines survival in hibernating mammals? Oecologia 2013, 173, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Anwar, M.B.; Nadeem, M.S.; Beg, M.A.; Kayani, A.R.; Muhammad, G. A photographic key for the identification of mammalian hairs of prey species in snow leopard (Panthera uncia) habitats of Gilgit-Baltistan Province of Pakistan. Pak. J. Zool. 2012, 44, 737–743. [Google Scholar]
- Armitage, K.B. Marmot Biology: Sociality, Individual Fitness, and Population Dynamics; Cambridge University Press: Newyork City, NY, USA, 2014. [Google Scholar]
- Abbas, Z.; Khan, S.M.; Alam, J.; Khan, S.W.; Abbasi, A.M. Medicinal plants used by inhabitants of the Shigar Valley, Baltistan region of Karakorum range-Pakistan. J. Ethnobiolo. Ethnomed. 2017, 13, 53. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Farooq, M.; Ahmed, M.; Zafar, M.U.; Akbar, M. Phytosociology and Structure of Central Karakoram National Park (CKNP) of Northern Areas of Pakistan. World Appl. Sci. J. 2010, 9, 1443–1449. [Google Scholar]
- Brady, K.M.; Armitage, K. Scent-marking in the yellow-bellied marmot (Marmota flaviventris). Ethol. Ecol. Evol. 1999, 11, 35–47. [Google Scholar] [CrossRef]
- Jiang, G.; Ma, J.; Zhang, M.; Stott, P. Multi-Scale Foraging Habitat Use and Interactions by Sympatric Cervids in Northeastern China. J. Wildl. Manag. 2010, 74, 678–689. [Google Scholar] [CrossRef]
- Blumstein, D.T.; Runyan, A.; Seymour, M.; Nicodemus, A.; Ozgul, A.; Ransler, F.; Im, S.; Stark, T.; Zugmeyer, C.; Daniel, J.C. Locomotor ability and wariness in yellow-bellied marmots. Ethology 2004, 110, 615–634. [Google Scholar]
- Blumstein, D.; Lea, A.; Olson, L.; Martin, J. Heritability of anti-predatory traits: Vigilance and locomotor performance in marmots. J. Evol. Biol. 2010, 23, 879–887. [Google Scholar] [PubMed]
- Blumstein, D.T.; Samia, D.S.; Stankowich, T.; Cooper, W.E., Jr. 16 Best practice for the study of escape behavior. In Escaping from Predators: An Integrative View of Escape Decisions; Cambridge University Press: Newyork City, NY, USA, 2015; Volume 407, pp. 410–416. [Google Scholar]
- Cooper, W.E.; Cooper, W.E., Jr.; Blumstein, D.T. Escaping from Predators: An Integrative View of Escape Decisions; Cambridge University Press: Newyork City, NY, USA, 2015; in press. [Google Scholar]
- Blumstein, D.T.; Daniel, J.C.; Springett, B.P. A test of the multi-predator hypothesis: rapid loss of antipredator behavior after 130 years of isolation. Ethology 2004, 110, 919–934. [Google Scholar]
- Blumstein, D.T.; Im, S.; Nicodemus, A.; Zugmeyer, C. Yellow-bellied marmots (Marmota flaviventris) hibernate socially. J. Mammal. 2004, 85, 25–29. [Google Scholar]
- Runyan, A.M.; Blumstein, D.T. Do individual differences influence flight initiation distance? J. Wildli. Manag. 2004, 68, 1124–1129. [Google Scholar]
- Blumstein, D.T.; Diaz, A.; Yin, L. Marmots do not consistently use their left eye to respond to an approaching threat but those that did fled sooner. Curr. Zool. 2018, 64, 727–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, K.N.; Chris, T. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–302. [Google Scholar] [CrossRef]
- Coppes, J.; Braunisch, V. Managing visitors in nature areas: where do they leave the trails? A spatial model. Wildl. Biol. 2013, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Focardi, S.; Rizzotto, M. Optimal strategies and complexity: A theoretical analysis of the anti-predatory behavior of the hare. Bull. Math. Biol. 1999, 61, 829–848. [Google Scholar] [CrossRef]
- Favreau, F.-R.; Pays, O.; Fritz, H.; Goulard, M.; Best, E.C.; Goldizen, A.W. Predators, food and social context shape the types of vigilance exhibited by kangaroos. Anim. Behav. 2015, 99, 109–121. [Google Scholar] [CrossRef]
- Ferretti, F.; Costa, A.; Corazza, M.; Pietrocini, V.; Cesaretti, G.; Lovari, S. Males are faster foragers than females: intersexual differences of foraging behaviour in the Apennine chamois. Behav. Ecol. Sociobiol. 2014, 68, 1335–1344. [Google Scholar] [CrossRef]
- Simpson, S.D.; Radford, A.N.; Nedelec, S.L.; Ferrari, M.C.; Chivers, D.P.; McCormick, M.I.; Meekan, M.G. Anthropogenic noise increases fish mortality by predation. Nat. Commun. 2016, 7, ncomms10544. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Variable Name | Description | Factor Levels |
|---|---|---|
| Treatment | Focal marmot approach by humans/dog | Pedestrian only |
| Pedestrian and dog | ||
| Human presence | Presence of additional pedestrians within view (e.g., tourist hikers, livestock farmers) | Binary—humans present (1) |
| Humans absent (0) | ||
| Agesex * | Combined age and sex class of focal marmot | Adult female |
| Adult male | ||
| Juvenile * | ||
| Yearling female Yearling male | ||
| Distance to road | Distance (m) from center of colony to nearest road measured using Arc GIS (10.2) near analysis tool | Continuous |
| Group size | Number of conspecifics within 10 m of focal marmot | Continuous |
| Distance to refuge | Distance (m) from focal marmot’s initial position to the nearest burrow | Continuous |
| Burrow type | Comprised of earthen or rocky type burrows | Earthen |
| Rocky | ||
| Vegetation height | Grass/shrub height estimated by eye | Tall (>30 cm) |
| Short (0–30 cm) | ||
| Viewshed | A measure of visibility on a TIN from the center of each colony to several observation points. Higher values represent greater visibility (i.e., of colony to predators). | Continuous |
| Slope | Slope of colony derived from a DEM (50 m) within the 3D Analyst extension of Arc GIS (10.2) | Continuous |
| Elevation (m.a.s.l) | Elevation in meters above sea level | Continuous |
| Year | Year of study | 2015 |
| (4-month study period per year) | 2016 | |
| 2017 | ||
| Starting Distance (SD) | Distance (m) between experimenter and focal marmot when the former begins approach | Continuous |
| Variable | Mean (± SE) | Minimum Value | Maximum Value |
|---|---|---|---|
| Elevation (m.a.s.l) | 2858.10 ± 29.99 | 2207 | 4792 |
| Viewshed (°) | 29.32 ± 14.82 | 20.12 | 50.88 |
| Distance to refuge (m) | 1.03 ± 4.54 | 0 | 221 |
| Distance to roads (m) | 811.63 ± 44.4 | 23.3 | 7987.5 |
| GLM | GLMM | ||||
|---|---|---|---|---|---|
| Variance Component | Variance | % of Total Variance | |||
| Colony ID | 0.162 | 0.02 | |||
| Residual | 3.731 | 0.04 | |||
| Explanatory variable (post hoc pairwise comparisons) | Estimate | Std. Error | % deviance explained * | t | P |
| Treatment (Dog versus pedestrian) | 0.527 | 0.186 | 4.380 | 2.823 | * |
| Vegetation height (Long versus short) | 0.660 | 0.100 | 6.360 | −6.563 | ** |
| Burrow type (Earthen versus rocky) | 0.386 | 0.120 | 16.040 | 3.269 | ** |
| Age/sex class | 35.470 | ||||
| AF vs. J | −0.014 | 0.338 | Na | −4.110 | *** |
| AM vs. J | −0.917 | 0.241 | Na | 3.807 | *** |
| AF vs. YM | −2.273 | 0.578 | Na | −4.725 | ** |
| AM vs. YF | 2.729 | 0.578 | Na | 4.725 | *** |
| AM vs. YM | −1.481 | 0.578 | Na | −4.725 | *** |
| Year | 8.500 | ||||
| 2015 vs. 2016 | −0.409 | 0.151 | Na | −2.703 | ** |
| 2017 vs. 2016 | −0.622 | 0.203 | Na | −3.060 | ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaman, M.; Tolhurst, B.A.; Zhu, M.; Jiang, G. Increased Flight Initiation Distance (FID) in Golden Marmots (Marmota caudata aurea) Responding to Domestic Dogs in A Landscape of Human Disturbance. Animals 2019, 9, 605. https://doi.org/10.3390/ani9090605
Zaman M, Tolhurst BA, Zhu M, Jiang G. Increased Flight Initiation Distance (FID) in Golden Marmots (Marmota caudata aurea) Responding to Domestic Dogs in A Landscape of Human Disturbance. Animals. 2019; 9(9):605. https://doi.org/10.3390/ani9090605
Chicago/Turabian StyleZaman, Muhammad, Bryony A. Tolhurst, Mengyan Zhu, and Guangshun Jiang. 2019. "Increased Flight Initiation Distance (FID) in Golden Marmots (Marmota caudata aurea) Responding to Domestic Dogs in A Landscape of Human Disturbance" Animals 9, no. 9: 605. https://doi.org/10.3390/ani9090605
APA StyleZaman, M., Tolhurst, B. A., Zhu, M., & Jiang, G. (2019). Increased Flight Initiation Distance (FID) in Golden Marmots (Marmota caudata aurea) Responding to Domestic Dogs in A Landscape of Human Disturbance. Animals, 9(9), 605. https://doi.org/10.3390/ani9090605