Antioxidative Effects of Olive Polyphenols Compared to Vitamin E in Piglets Fed a Diet Rich in N-3 PUFA


Simple Summary
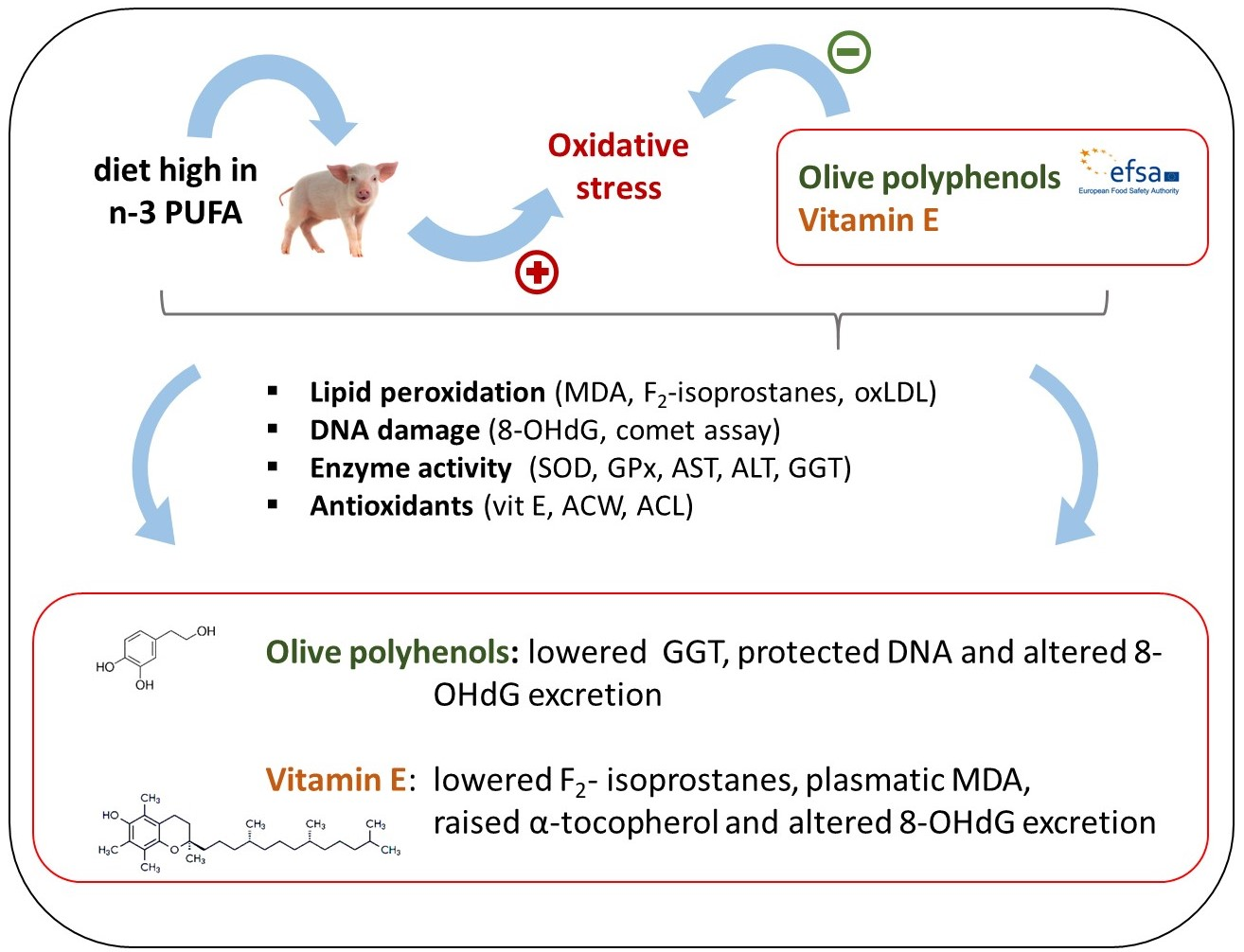
Abstract

1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design and Dietary Treatments
2.2. Preparation and Standardization of Olive Leaf Extract
2.3. Sample Collection
2.4. Chemical Analyses
2.5. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Extent of Lipid Oxidation
3.3. Level of Antioxidants and Antioxidant Enzymes
3.4. Rate of DNA Damage and Activity of Liver Enzymes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vissers, M.N.; Zock, P.L.; Katan, M.B. Bioavailability and antioxidant effects of olive oil phenols in humans: A review. Eur. J. Clin. Nutr. 2004, 58, 955–965. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA) Panel on Dietetic Products Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to polyphenols in olive and protection of LDL particles from oxidative damage. EFSA J. 2011, 9, 2033. [Google Scholar]
- Paiva-Martins, F.; Barbosa, S.; Silva, M.; Monteiro, D.; Pinheiro, V.; Mourão, J.L.; Fernandes, J.; Rocha, S.; Belo, L.; Santos-Silva, A. The effect of olive leaf supplementation on the constituents of blood and oxidative stability of red blood cells. J. Funct. Foods 2014, 9, 271–279. [Google Scholar] [CrossRef]
- Botsoglou, E.; Govaris, A.; Ambrosiadis, I.; Fletouris, D. Lipid and protein oxidation of α-linolenic acid-enriched pork during refrigerated storage as influenced by diet supplementation with olive leaves (Olea europea L.) or α-tocopheryl acetate. Meat Sci. 2012, 92, 525–532. [Google Scholar] [CrossRef]
- Paiva-Martins, F.; Ribeirinha, T.; Silva, A.; Gonçalves, R.; Pinheiro, V.; LMourão, J.; Outor-Monteiro, D. Effects of the dietary incorporation of olive leaves on growth performance, digestibility, blood parameters and meat quality of growing pigs. J. Sci. Food Agric. 2014, 94, 3023–3029. [Google Scholar] [CrossRef]
- Frankič, T.; Levart, A.; Salobir, J. The effect of vitamin E and plant extract mixture composed of carvacrol, cinnamaldehyde and capsaicin on oxidative stress induced by high PUFA load in young pigs. Animal 2010, 4, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and antioxidants in disease: Oxidative stress in farm animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Swine; National Academies Press: Washington, DC, USA, 2012; ISBN 978-0-309-22423-9. [Google Scholar]
- Naumann, C.; Bassler, R. Methodenbuch Band LLL: Die Chemische Untersuchung von Futtermitteln; VDLUFA-Verlag: Darmstadt, Germany, 1997. [Google Scholar]
- Voljč, M.; Frankič, T.; Levart, A.; Nemec, M.; Salobir, J. Evaluation of different vitamin e recommendations and bioactivity of α-tocopherol isomers in broiler nutrition by measuring oxidative stress in vivo and the oxidative stability of meat. Poult. Sci. 2011, 90, 1478–1488. [Google Scholar] [CrossRef]
- Frankič, T.; Salobir, J. In vivo antioxidant potential of Sweet chestnut (Castanea sativa Mill.) wood extract in young growing pigs exposed to n-3 PUFA-induced oxidative stress. J. Sci. Food Agric. 2011, 91, 1432–1439. [Google Scholar] [CrossRef]
- Surai, P.F. Polyphenol compounds in the chicken/animal diet: From the past to the future. J. Anim. Physiol. Anim. Nutr. (Berl.) 2014, 98, 19–31. [Google Scholar] [CrossRef]
- Halliwell, B. Establishing the Significance and Optimal Intake of Dietary Antioxidants: The Biomarker Concept. Nutr. Rev. 2009, 57, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Caruso, D.; Galli, C.; Viappiani, S.; Galli, G.; Sala, A. Olive oils rich in natural catecholic phenols decrease isoprostane excretion in humans. Biochem. Biophys. Res. Commun. 2000, 278, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G.; Atkinson, J. Vitamin E, antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef]
- Covas, M.I.; Nyyssönen, K.; Poulsen, H.E.; Kaikkonen, J.; Zunft, H.J.F.; Kiesewetter, H.; Gaddi, A.; De La Torre, R.; Mursu, J.; Bäumler, H.; et al. The effect of polyphenols in olive oil on heart disease risk factors: A randomized trial. Ann. Intern. Med. 2006, 145, 333–341. [Google Scholar] [CrossRef]
- Frankič, T.; Salobir, K.; Salobir, J. The comparison of in vivo antigenotoxic and antioxidative capacity of two propylene glycol extracts of Calendula officinalis (marigold) and vitamin e in young growing pigs. J. Anim. Physiol. Anim. Nutr. (Berl.) 2009, 93, 688–694. [Google Scholar] [CrossRef]
- Andreadou, I.; Iliodromitis, E.K.; Mikros, E.; Constantinou, M.; Agalias, A.; Magiatis, P.; Skaltsounis, A.L.; Kamber, E.; Tsantili-Kakoulidou, A.; Kremastinos, D.T. The Olive Constituent Oleuropein Exhibits Anti-Ischemic, Antioxidative, and Hypolipidemic Effects in Anesthetized Rabbits. J. Nutr. 2006, 136, 2213–2219. [Google Scholar] [CrossRef]
- Al-Azzawie, H.F.; Alhamdani, M.S.S. Hypoglycemic and antioxidant effect of oleuropein in alloxan-diabetic rabbits. Life Sci. 2006, 78, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Buckley, D.J.; Morrissey, P.A.; Gray, J.I. Influence of dietary vitamin E on the oxidative stability and quality of pig meat. J. Anim. Sci. 1995, 73, 3122–3130. [Google Scholar] [CrossRef]
- Traber, G.; Kayden, J. Preferential incorporation in human Iipoproteins13 of a-tocopherol vs ‘ y-tocopherol. Am. J. Clin. Nutr. 1989, 49, 517–526. [Google Scholar] [CrossRef]
- De Moffarts, B.; Kirschvink, N.; Art, T.; Pincemail, J.; Lekeux, P. Effect of oral antioxidant supplementation on blood antioxidant status in trained thoroughbred horses. Vet. J. 2005, 169, 65–74. [Google Scholar] [CrossRef]
- Juśkiewicz, J.; Zduńczyk, Z.; Zary-Sikorska, E.; Król, B.; Milala, J.; Jurgoński, A. Effect of the dietary polyphenolic fraction of chicory root, peel, seed and leaf extracts on caecal fermentation and blood parameters in rats fed diets containing prebiotic fructans. Br. J. Nutr. 2011, 105, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Free radicals and antioxidants—Quo vadis? Trends Pharmacol. Sci. 2011, 32, 125–130. [Google Scholar] [CrossRef]
- Masella, R.; Varì, R.; D’Archivio, M.; Di Benedetto, R.; Matarrese, P.; Malorni, W.; Scazzocchio, B.; Giovannini, C. Extra Virgin Olive Oil Biophenols Inhibit Cell-Mediated Oxidation of LDL by Increasing the mRNA Transcription of Glutathione-Related Enzymes. J. Nutr. 2004, 134, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Cumaoǧlu, A.; Rackova, L.; Stefek, M.; Kartal, M.; Maechler, P.; Karasu, Ç. Effects of olive leaf polyphenols against H2O2 toxicity in insulin secreting β-cells. Acta Biochim. Pol. 2011, 58, 45–50. [Google Scholar] [CrossRef]
- Hassen, I.; Casabianca, H.; Hosni, K. Biological activities of the natural antioxidant oleuropein: Exceeding the expectation—A mini-review. J. Funct. Foods 2015, 18, 926–940. [Google Scholar] [CrossRef]
- Hamden, K.; Allouche, N.; Damak, M.; Elfeki, A. Hypoglycemic and antioxidant effects of phenolic extracts and purified hydroxytyrosol from olive mill waste in vitro and in rats. Chem. Biol. Interact. 2009, 180, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Oliveras-López, M.J.; Berná, G.; Jurado-Ruiz, E.; López-García de la Serrana, H.; Martín, F. Consumption of extra-virgin olive oil rich in phenolic compounds has beneficial antioxidant effects in healthy human adults. J. Funct. Foods 2014, 10, 475–484. [Google Scholar] [CrossRef]
- Romeu, M.; Rubió, L.; Sánchez-Martos, V.; Castañer, O.; De La Torre, R.; Valls, R.M.; Ras, R.; Pedret, A.; Catalán, Ú.; López De Las Hazas, M.D.C.; et al. Virgin Olive Oil Enriched with Its Own Phenols or Complemented with Thyme Phenols Improves DNA Protection against Oxidation and Antioxidant Enzyme Activity in Hyperlipidemic Subjects. J. Agric. Food Chem. 2016, 64, 1879–1888. [Google Scholar] [CrossRef]
- Serini, S.; Fasano, E.; Piccioni, E.; Cittadini, A.R.M.; Calviello, G. Dietary n-3 polyunsaturated fatty acids and the paradox of their health benefits and potential harmful effects. Chem. Res. Toxicol. 2011, 24, 2093–2105. [Google Scholar] [CrossRef]
- Fabiani, R.; Rosignoli, P.; De Bartolomeo, A.; Fuccelli, R.; Servili, M.; Montedoro, G.F.; Morozzi, G. Oxidative DNA damage is prevented by extracts of olive oil, hydroxytyrosol, and other olive phenolic compounds in human blood mononuclear cells and HL60 cells. J. Nutr. 2008, 138, 1411–1416. [Google Scholar] [CrossRef]
- Lim, J.S.; Yang, J.H.; Chun, B.Y.; Kam, S.; Jacobs, D.R.; Lee, D.H. Is serum γ-glutamyltransferase inversely associated with serum antioxidants as a marker of oxidative stress? Free Radic. Biol. Med. 2004, 37, 1018–1023. [Google Scholar] [CrossRef] [PubMed]
- De Bock, M.; Derraik, J.G.B.; Brennan, C.M.; Biggs, J.B.; Morgan, P.E.; Hodgkinson, S.C.; Hofman, P.L.; Cutfield, W.S. Olive (Olea europaea L.) Leaf Polyphenols Improve Insulin Sensitivity in Middle-Aged Overweight Men: A Randomized, Placebo-Controlled, Crossover Trial. PLoS ONE 2013, 8, e57622. [Google Scholar] [CrossRef]
- Botsoglou, E.; Govaris, A.; Christaki, E.; Botsoglou, N. Effect of dietary olive leaves and/or α-tocopheryl acetate supplementation on microbial growth and lipid oxidation of turkey breast fillets during refrigerated storage. Food Chem. 2010, 121, 17–22. [Google Scholar] [CrossRef]
- Paiva-Martins, F.; Fernandes, J.; Rocha, S.; Nascimento, H.; Vitorino, R.; Amado, F.; Borges, F.; Belo, L.; Santos-Silva, A. Effects of olive oil polyphenols on erythrocyte oxidative damage. Mol. Nutr. Food Res. 2009, 53, 609–616. [Google Scholar] [CrossRef]
- Zontar, T.P.; Salobir, J.; Rezar, V. Time dependent formation of markers of oxidative stress induced by a high fat diet supplemented or unsupplemented with vitamin E in pigs. Acta Agric. Slov. 2009, 94, 103–110. [Google Scholar]
- Dittmar, M.; Knuth, M.; Beineke, M.; Epe, B. Role of Oxidative DNA Damage and Antioxidative Enzymatic Defence Systems in Human Aging. Open Anthropol. J. 2008, 1, 38–45. [Google Scholar] [CrossRef]
- Lee, J.; Koo, N.; Min, D.B. Species, Aging, and Antioxidative Nutraceuticals. Compr. Rev. Food Sci. Food Saf. 2004, 3, 21–33. [Google Scholar] [CrossRef]

{kind=link}
| Component | Cont− | Cont+, Vit-E, OLE-1, OLE-2, OLE-3 |
|---|---|---|
| Composition (g/kg) | ||
| Wheat starch | 379.37 | 219.82 |
| Soybean meal | 220.75 | 247.87 |
| Skimmed milk powder | 125.69 | 141.15 |
| Full-fat milk powder | 61.82 | 69.42 |
| Maize | 112.56 | 126.38 |
| Wheat bran | 47.05 | 52.83 |
| Rapeseed oil | 18.99 | 21.32 |
| Linseed oil | - | 83.30 |
| Sodium chloride | 2.43 | 2.73 |
| Limestone | 9.72 | 10.92 |
| Monocalcium phosphate | 8.67 | 9.73 |
| L-lysine | 4.02 | 4.51 |
| DL-methionine | 2.35 | 2.63 |
| Threonine | 1.80 | 2.02 |
| Premix 1 | 4.78 | 5.37 |
| Chemical composition of diets (g/kg) | ||
| Dry matter | 903 | 917 |
| Crude protein | 184.4 | 205.3 |
| Crude fat | 37.28 | 125.21 |
| Crude ash | 46.96 | 52.03 |
| NDF 2 | 67.91 | 72.85 |
| ADF 3 | 23.49 | 23.96 |
| Ca | 8.83 | 9.67 |
| P | 5.85 | 6.44 |
| Metabolisable energy (MJ/kg) | 14.99 | 16.84 |
| Fatty acids (g/kg) 4 | ||
| C16:0 | 6.09 | 12.09 |
| C18:0 | 1.68 | 5.55 |
| C18:1 | 13.84 | 34.38 |
| C18:2 n-6 | 8.08 | 21.30 |
| C18:3 n-3 | 1.55 | 39.59 |
| n-6 PUFA 5 | 8.12 | 21.41 |
| n-3 PUFA 5 | 1.55 | 39.62 |
| n-6/n-3 PUFA 5 | 5.24 | 0.54 |
| Sum of SFA 6 | 11.05 | 22.12 |
| Sum of MUFA 7 | 14.63 | 35.71 |
| Sum of PUFA 5 | 9.73 | 61.12 |
| Daily Intake | Diets | |||||
|---|---|---|---|---|---|---|
| Cont− | Cont+ | Vit-E | OLE-1 | OLE-2 | OLE-3 | |
| Feed (g) 1 | 473.1 | 421.4 | 421.4 | 421.4 | 421.4 | 421.4 |
| Hydroxytyrosol EQ (mg) 2 | - | - | - | 3.84 | 38.4 | 96.0 |
| α-tocopherol (mg) | 12.91 | 10.65 | 102.40 | 11.77 | 12.17 | 12.22 |
| γ-tocopherol (mg) | 4.28 | 17.13 | 17.96 | 17.96 | 18.50 | 17.72 |
| Vitamin E (IU) 3 | 16.8 | 12.3 | 105.4 | 14.8 | 15.4 | 15.5 |
| Energy from fat (%) 4 | 11.2 | 29.4 | 29.4 | 29.4 | 29.4 | 29.4 |
| Energy from PUFA (%) 4 | 2.9 | 15.0 | 15.0 | 15.0 | 15.0 | 15.0 |
| Energy from n-3 PUFA (%) 4 | 0.6 | 10.4 | 10.4 | 10.4 | 10.4 | 10.4 |
| Parameter | Group | p-Value | |||||
|---|---|---|---|---|---|---|---|
| Cont− | Cont+ | Vit-E | OLE-1 | OLE-2 | OLE-3 | ||
| Initial body weight (kg) | 10.6 ± 0.25 | 10.6 ± 0.25 | 10.6 ± 0.23 | 10.6 ± 0.21 | 10.7 ± 0.19 | 10.6 ± 0.22 | 0.996 |
| Final body weight (kg) | 17.8 ± 0.28 | 17.9 ± 0.33 | 18.2 ± 0.30 | 17.9 ± 0.21 | 17.7 ± 0.26 | 17.9 ± 0.31 | 0.823 |
| Average daily gain (g) | 346 ± 4.9 | 348 ± 8.0 | 360 ± 6.1 | 344 ± 9.9 | 330 ± 9.1 | 349 ± 5.6 | 0.169 |
| Average daily feed intake (g) | 535 ± 6.0 b | 477 ± 5.9 a | 481 ± 4.9 a | 479 ± 5.6 a | 477 ± 4.2 a | 479 ± 5.3 a | <0.001 |
| Conversion ratio 1 | 1.55 ± 0.023 b | 1.37 ± 0.030 a | 1.34 ± 0.021 a | 1.40 ± 0.048 ab | 1.45 ± 0.040 ab | 1.38 ± 0.014 a | <0.001 |
| Parameter | Group | p-Value | |||||
|---|---|---|---|---|---|---|---|
| Cont− | Cont+ | Vit-E | OLE-1 | OLE-2 | OLE-3 | ||
| Urinary F2-isoprostanes (ng/24 h) 1 | 1961 ± 165.9 b | 1496 ±130.5 ab | 1270 ± 144.5 a | 1614 ± 204.2 ab | 1456 ± 232.9 ab | 2086 ± 356.8 ab | 0.0483 |
| Serum oxLDL (nmol/mL) 2 | 171 ± 17.0 | 169 ± 10.1 | 167 ± 13.5 | 139 ± 25.5 | 132 ± 23.0 | 136 ± 13.4 | 0.2734 |
| Plasma MDA (nmol/mL) 3 | 0.596 ± 0.045 a | 0.863 ± 0.042 bd | 0.670 ± 0.058 ab | 0.873 ± 0.032 cd | 0.864 ± 0.064 bc | 0.859 ± 0.057 bc | 0.0001 |
| Urinary MDA (µmol/24 h) 3 | 5.57 ± 0.454 | 7.62 ± 0.622 | 5.92 ± 0.624 | 7.83 ± 0.710 | 5.80 ± 0.734 | 7.62 ± 0.553 | 0.0169 |
| Parameter | Group | p-Value | |||||
|---|---|---|---|---|---|---|---|
| Cont− | Cont+ | Vit-E | OLE-1 | OLE-2 | OLE-3 | ||
| Plasma α-tocopherol (µg/mL) | 1.74 ± 0.143 a | 1.54 ± 0.118 a | 4.52 ± 0.221 b | 1.60 ± 0.169 a | 1.58 ± 0.191 a | 1.39 ± 0.071 a | <0.001 |
| Plasma γ-tocopherol (µg/mL) | 0.084 ± 0.011 a | 0.546 ± 0.046 c | 0.209 ± 0.019 b | 0.460 ± 0.064 c | 0.413 ± 0.046 c | 0.346 ± 0.052 bc | <0.001 |
| Serum ACW (nmol/mL) 1 | 65.7 ± 3.35 | 67.8 ± 2.92 | 73.8 ± 2.42 | 78.7 ± 5.00 | 79.8 ± 3.70 | 71.1 ± 3.31 | 0.0499 |
| Serum ACL (nmol/mL) 2 | 112 ± 3.78 | 123 ± 5.57 | 115 ± 4.65 | 123 ± 4.22 | 123 ± 7.20 | 125 ± 5.20 | 0.2143 |
| Whole blood GPx (U/g HGB) 3 | 261 ± 12.5 | 296 ± 26.1 | 289 ± 12.9 | 241 ± 12.2 | 249 ± 6.0 | 273 ± 14.8 | 0.0404 |
| Whole blood SOD (U/g HGB) 4 | 1839 ± 65.3 | 1781 ± 72.0 | 1687 ± 106.5 | 1743 ± 68.0 | 1793 ± 110.3 | 1858 ± 72.9 | 0.7215 |
| Parameter | Group | p-Value | |||||
|---|---|---|---|---|---|---|---|
| Cont− | Cont+ | Vit-E | OLE-1 | OLE-2 | OLE-3 | ||
| Tail DNA (%) | 7.91 ± 0.257 ad | 9.07 ± 0.305 bc | 9.75 ± 0.324 b | 7.92 ± 0.298 cd | 8.88 ± 0.307 abc | 8.38 ± 0.300 ac | <0.0001 |
| OTM1 | 0.730 ± 0.032 a | 0.830 ± 0.031 a | 0.986 ± 0.032 b | 0.729 ± 0.030 a | 0.820 ± 0.032 a | 0.842 ± 0.030 a | <0.0001 |
| Urinary 8-OHdG (µg/24 h) 2 | 418 ± 41.7 bc | 253 ± 26.9 ad | 389 ± 31.2 c | 345 ± 28.3 bcd | 237 ± 31.9 ad | 236 ± 18.9 a | 0.0001 |
| Serum AST (U/L) 3 | 47.6 ± 3.50 | 64.5 ± 6.28 | 47.6 ± 3.73 | 58.5 ± 7.31 | 58.3 ± 6.86 | 47.8 ± 4.74 | 0.1370 |
| Serum ALT (U/L) 4 | 85.0 ± 9.09 | 93.9 ± 7.95 | 92.8 ± 8.72 | 98.1 ± 7.47 | 95.5 ± 7.35 | 82.6 ± 6.36 | 0.6070 |
| Serum GGT (U/L) 5 | 37.0 ± 2.01 a | 51.9 ± 4.09 b | 35.6 ± 4.13 ab | 38.1 ± 1.65 a | 44.5 ± 4.66 ab | 46.0 ± 2.42 ab | 0.0051 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leskovec, J.; Rezar, V.; Nemec Svete, A.; Salobir, J.; Levart, A. Antioxidative Effects of Olive Polyphenols Compared to Vitamin E in Piglets Fed a Diet Rich in N-3 PUFA. Animals 2019, 9, 161. https://doi.org/10.3390/ani9040161
Leskovec J, Rezar V, Nemec Svete A, Salobir J, Levart A. Antioxidative Effects of Olive Polyphenols Compared to Vitamin E in Piglets Fed a Diet Rich in N-3 PUFA. Animals. 2019; 9(4):161. https://doi.org/10.3390/ani9040161
Chicago/Turabian StyleLeskovec, Jakob, Vida Rezar, Alenka Nemec Svete, Janez Salobir, and Alenka Levart. 2019. "Antioxidative Effects of Olive Polyphenols Compared to Vitamin E in Piglets Fed a Diet Rich in N-3 PUFA" Animals 9, no. 4: 161. https://doi.org/10.3390/ani9040161
APA StyleLeskovec, J., Rezar, V., Nemec Svete, A., Salobir, J., & Levart, A. (2019). Antioxidative Effects of Olive Polyphenols Compared to Vitamin E in Piglets Fed a Diet Rich in N-3 PUFA. Animals, 9(4), 161. https://doi.org/10.3390/ani9040161