The Interaction between Phagocytes and Streptococcus agalactiae (GBS) Mediated by the Activated Complement System is the Key to GBS Inducing Acute Bacterial Meningitis of Tilapia


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Fishes
2.2. Ethics Statement
2.3. Challenge and Behavioral Assessments
2.4. Blood Sampling
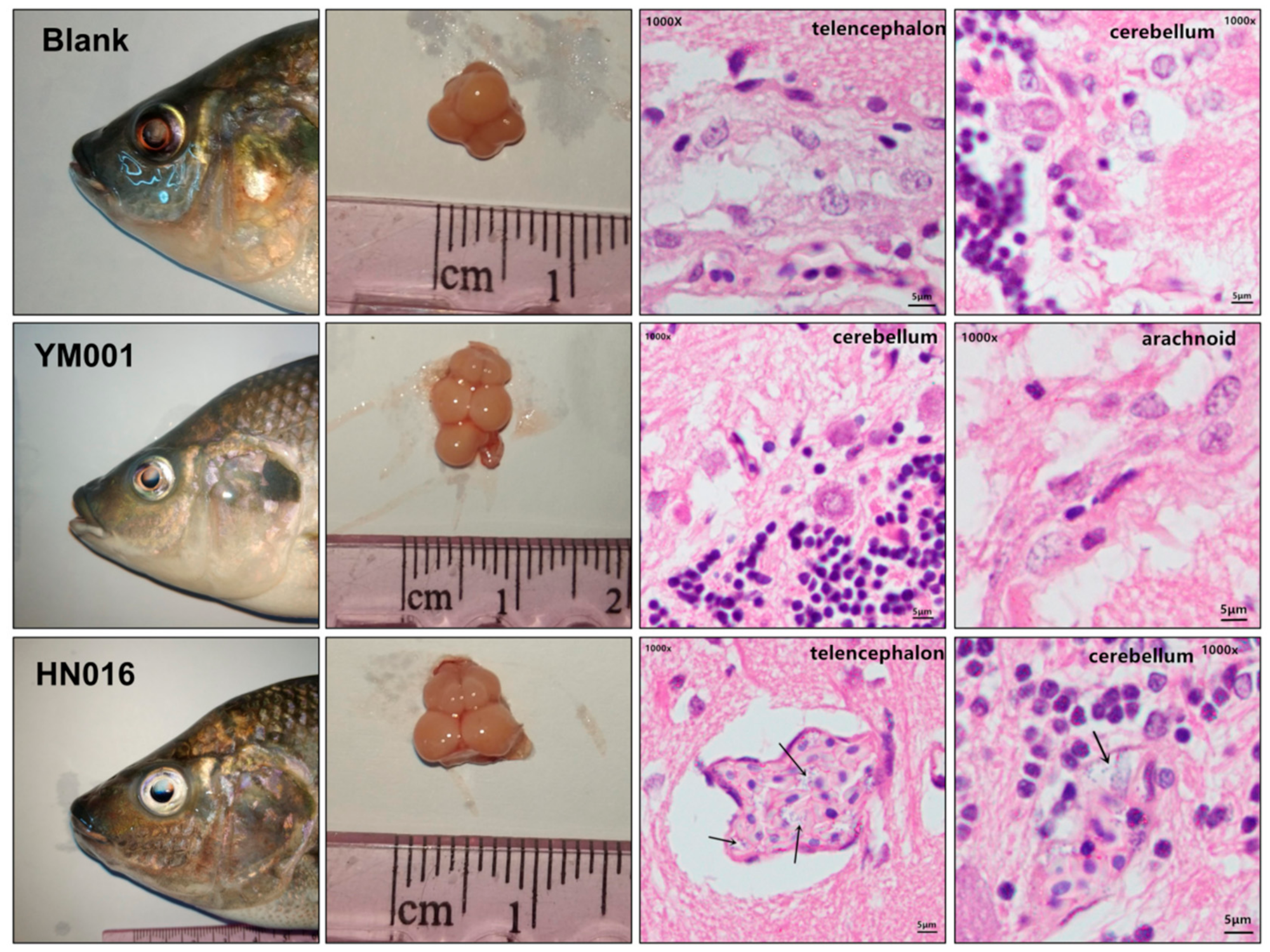
2.5. Tissue Injury Pathological Analysis
2.6. Dual RNA-seq Transcriptional Analysis
2.7. TMT Protein Labeling and LC–MS/MS Proteomic Analyses
2.8. Validation of Gene Expression Level by Real-Time Quantitative PCR (RT-qPCR)
3. Results
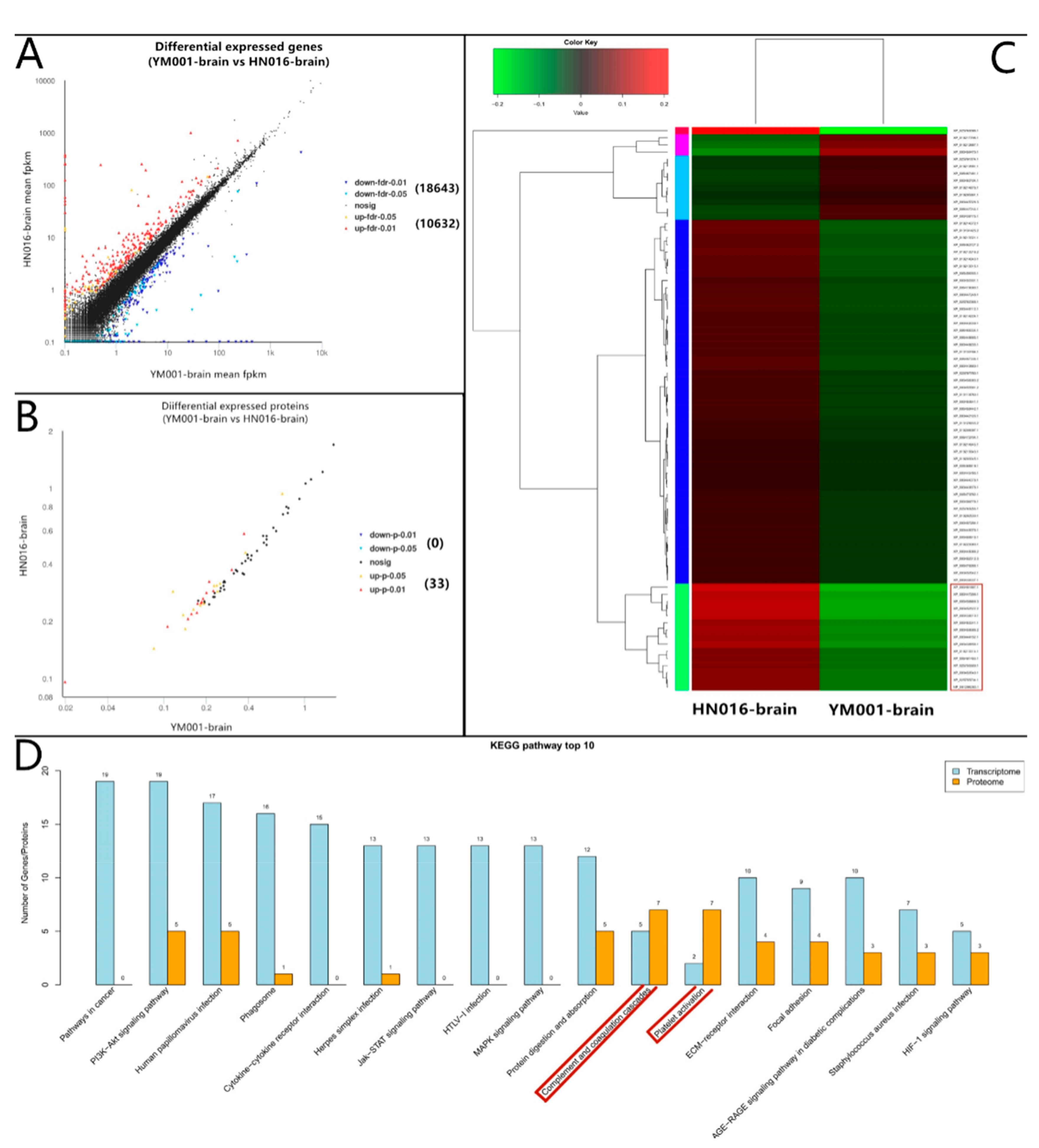
3.1. Overviews of RNA Transcriptomic and Quantitative Proteomic Analyses Profiles
3.2. Histopathological Analysis
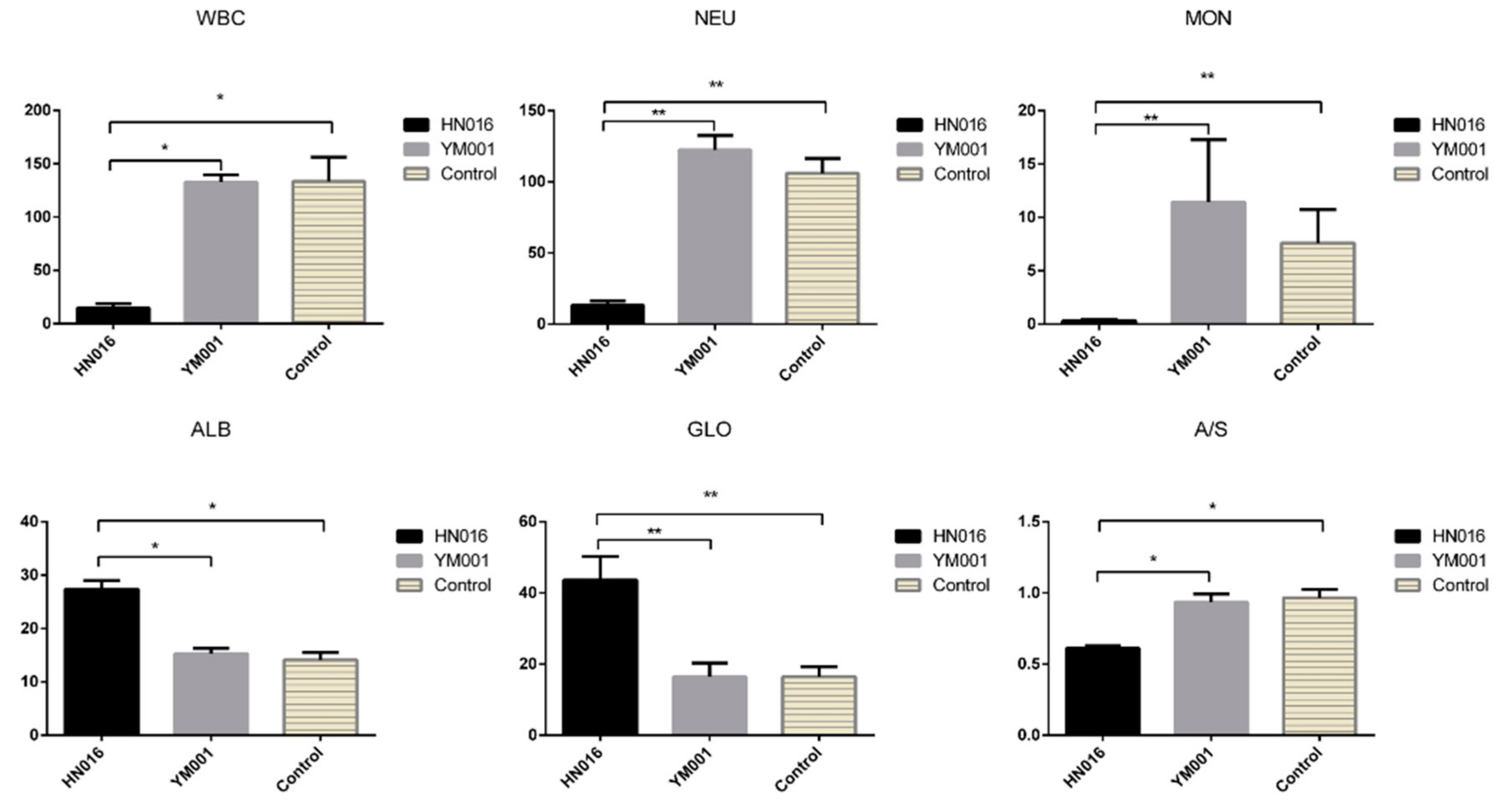
3.3. Effects of HN016 and YM001 on Blood Parameters of Tilapia
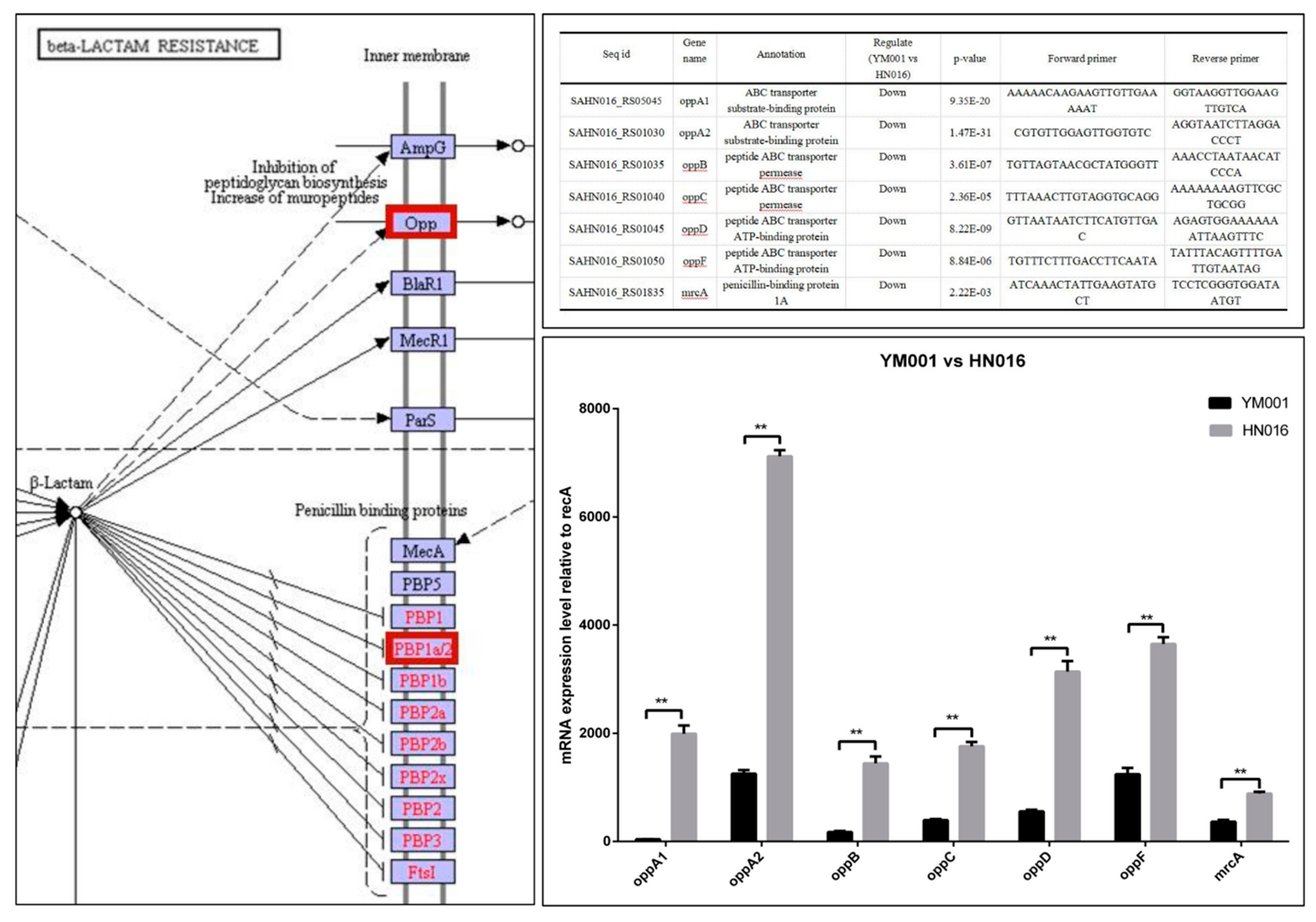
3.4. Bioinformatics Analysis of DEGs and DEPs
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| WBC | white blood cell |
| MON | monocytes |
| NEU | neutrophil |
| DEGs | differential expressed genes |
| DEPs | differential expressed proteins |
| GBS | Streptococcus agalactiae |
| PTS | phosphotransferase system |
References
- Ye, X.; Li, J.; Lu, M.; Deng, G.; Jiang, X.; Tian, Y.; Quan, Y.; Jian, Q. Identification and molecular typing of Streptococcus agalactiae isolated from pond-cultured tilapia in China. Fish. Sci. 2011, 77, 623–632. [Google Scholar] [CrossRef]
- Li, J.; Lü, H.; Zhu, J.; Wang, Y.; Li, X. Aquatic products processing industry in China: Challenges and outlook. Trends Food Sci. Technol. 2009, 20, 73–77. [Google Scholar] [CrossRef]
- Chen, M.; Li, L.-P.; Wang, R.; Liang, W.-W.; Huang, Y.; Li, J.; Lei, A.-Y.; Huang, W.-Y.; Gan, X. PCR detection and PFGE genotype analyses of streptococcal clinical isolates from tilapia in China. Veter Microbiol. 2012, 159, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Liu, C.; Deng, Y.; Cheng, C.; Ma, H.; Guo, Z.; Feng, J. Molecular typing of Streptococcus agalactiae isolates of serotype Ia from tilapia in southern China. FEMS Microbiol. Lett. 2019, 366, 366. [Google Scholar] [CrossRef] [PubMed]
- Eto, S.F.; Fernandes, D.C.; Moraes, A.C.; Prado, E.J.R.; Baldassi, A.C.; Manrique, W.G.; Silva, I.C.; Medeiros, A.S.; Belo, M.A.; Balbuena, T.S.; et al. Validation of IgY for the diagnosis of Streptococcus agalactiae-caused endocarditis and bacterial meningitis in Nile tilapia (Oreochromis niloticus). Fish Shellfish. Immunol. 2018, 76, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Genomic Diversity of Streptoccocus agalactiae Isolates from Multiple Hosts and Their Infectivity in Nile Tilapia. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=4&ved=2ahUKEwi-4ZmH9p_lAhV8yYsBHUHHByMQFjADegQIBhAC&url=https%3A%2F%2Fag.arizona.edu%2Fazaqua%2Fista%2FISTA8%2FJoyce.doc&usg=AOvVaw28CiplwywSDB0W_Nw3rp08 (accessed on 16 October 2019).
- Barnes, A.C.; Silayeva, O. Vaccination against streptococcal infections in farmed fish. Microbiol. Aust. 2016. [Google Scholar] [CrossRef]
- Kim, K.S. Mechanisms of microbial traversal of the blood-brain barrier. Nat. Rev. Genet. 2008, 6, 625–634. [Google Scholar] [CrossRef]
- Tenenbaum, T.; Spellerberg, B.; Adam, R.; Vogel, M.; Kim, K.; Schroten, H. Streptococcus agalactiae invasion of human brain microvascular endothelial cells is promoted by the laminin-binding protein Lmb. Microbes Infect. 2007, 9, 714–720. [Google Scholar] [CrossRef]
- Guo, C.-M.; Chen, R.-R.; Kalhoro, D.H.; Wang, Z.-F.; Liu, G.-J.; Lu, C.-P.; Liu, Y.-J. Identification of Genes Preferentially Expressed by Highly Virulent Piscine Streptococcus agalactiae upon Interaction with Macrophages. PLoS ONE 2014, 9, e87980. [Google Scholar] [CrossRef]
- Drevets, D.A. Dissemination of Listeria monocytogenes by Infected Phagocytes. Infect. Immun. 1999, 67, 3512–3517. [Google Scholar]
- Drevets, D.A.; Jelinek, T.A.; Freitag, N.E. Listeria monocytogenes-Infected Phagocytes Can Initiate Central Nervous System Infection in Mice. Infect. Immun. 2001, 69, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Drevets, D.A.; Sawyer, R.T.; Potter, T.A.; Campbell, P.A. Listeria monocytogenes infects human endothelial cells by two distinct mechanisms. Infect. Immun. 1995, 63, 4268–4276. [Google Scholar] [PubMed]
- Gutekunst, H.; Eikmanns, B.J.; Reinscheid, D.J. The Novel Fibrinogen-Binding Protein FbsB Promotes Streptococcus agalactiae Invasion into Epithelial Cells. Infect. Immun. 2004, 72, 3495–3504. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, C.; Waggoner, J.D.; Harris, T.O.; Tamura, G.S.; Rubens, C.E. Identification of Novel Adhesins from Group B Streptococci by Use of Phage Display Reveals that C5a Peptidase Mediates Fibronectin Binding. Infect. Immun. 2002, 70, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Doran, K.S.; Engelson, E.J.; Khosravi, A.; Maisey, H.C.; Fedtke, I.; Equils, O.; Michelsen, K.S.; Arditi, M.; Peschel, A.; Nizet, V. Blood-brain barrier invasion by group B Streptococcus depends upon proper cell-surface anchoring of lipoteichoic acid. J. Clin. Investig. 2005, 115, 2499–2507. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhang, W.; Lu, C. Comparative genomics analysis of Streptococcus agalactiae reveals that isolates from cultured tilapia in China are closely related to the human strain A909. BMC Genomics 2013, 14, 775. [Google Scholar] [CrossRef]
- Wang, R.; Li, L.; Huang, Y.; Luo, F.; Liang, W.; Gan, X.; Huang, T.; Lei, A.; Chen, M.; Chen, L. Comparative genome analysis identifies two large deletions in the genome of highly-passaged attenuated Streptococcus agalactiae strain YM001 compared to the parental pathogenic strain HN016. BMC Genomics 2015, 16, 897. [Google Scholar] [CrossRef]
- Kayansamruaj, P.; Pirarat, N.; Katagiri, T.; Hirono, I.; Rodkhum, C. Molecular characterization and virulence gene profiling of pathogenic Streptococcus agalactiae populations from tilapia (Oreochromis sp.) farms in Thailand. J. Veter Diagn. Investig. 2014, 26, 488–495. [Google Scholar] [CrossRef]
- Li, L.; Wang, R.; Liang, W.; Huang, T.; Huang, Y.; Luo, F.; Lei, A.; Chen, M.; Gan, X. Development of live attenuated Streptococcus agalactiae vaccine for tilapia via continuous passage in vitro. Fish Shellfish. Immunol. 2015, 45, 955–963. [Google Scholar] [CrossRef]
- Westermann, A.J.; Gorski, S.A.; Vogel, J. Dual RNA-seq of pathogen and host. Nat. Rev. Genet. 2012, 10, 618–630. [Google Scholar] [CrossRef]
- Keel, B.N.; Snelling, W.M. Comparison of burrows-wheeler transform-based mapping algorithms used in high-throughput whole-genome sequencing: application to illumina data for livestock genomes1. Front. Genet. 2018, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Mao, X.; Cai, T.; Luo, J.; Wei, L. KOBAS server: A web-based platform for automated annotation and pathway identification. Nucleic Acids Res. 2006, 34, W720–W724. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed]
- Florindo, C.; Ferreira, R.; Borges, V.; Spellerberg, B.; Gomes, J.P.; Borrego, M.J. Selection of reference genes for real-time expression studies in Streptococcus agalactiae. J. Microbiol. Methods 2012, 90, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Song, Y.-L.; Wang, B.; Zhang, X.-Y.; Zhang, X.-J.; Wang, Y.-L.; Cheng, Y.-Y.; Chen, D.-D.; Xia, X.-Q.; Lu, Y.-S.; et al. Fish gut-liver immunity during homeostasis or inflammation revealed by integrative transcriptome and proteome studies. Sci. Rep. 2016, 6, 36048. [Google Scholar] [CrossRef]
- Liu, Y.; Li, L.; Huang, T.; Wang, R.; Liang, W.; Yang, Q.; Lei, A.; Chen, M. Comparative multi-omics systems analysis reveal the glycolysis/gluconeogenesis signal pathway play an important role in virulence attenuation in fish-derived GBS YM001. PLoS ONE 2019, 14, e0221634. [Google Scholar] [CrossRef]
- Dabrowska-Szponar, M.; Galiński, J. Drug resistance of group B streptococci. Polski Merkur. Lek. Organ Polskiego Towar. Lek. 2001, 10, 442–444. [Google Scholar]
- Poole, K. Resistance to β-lactam antibiotics. Cell. Mol. Life Sci. 2004, 61, 2200–2223. [Google Scholar] [CrossRef]
- May, S.M.; Hartz, M.F.; Joshi, A.Y.; Park, M.A. Intrapartum antibiotic exposure for group B Streptococcus treatment did not increase penicillin allergy in children. Ann. Allergy Asthma Immunol. 2016, 116, 134–138. [Google Scholar] [CrossRef]
- James, D.C. Maternal screening and treatment for group B streptococcus. J. Obstet. Gynecol. Neonatal Nurs. 2001, 30, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Scasso, S.; Laufer, J.; Rodriguez, G.; Alonso, J.G.; Sosa, C.G. Vaginal group b streptococcus status during intrapartum antibiotic prophylaxis. Int. J. Gynaecol. Obstet. 2015, 129, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Turrentine, M.A.; Greisinger, A.J.; Brown, K.S.; Wehmanen, O.A.; Mouzoon, M.E. Duration of Intrapartum Antibiotics for Group B Streptococcus on the Diagnosis of Clinical Neonatal Sepsis. Infect. Dis. Obstet. Gynecol. 2013, 2013, 525878. [Google Scholar] [CrossRef] [PubMed]
- Muller-Eberhard, J.H. Molecular organization and function of the complement system. Annu. Rev. Biochem. 1988, 57, 321–347. [Google Scholar] [CrossRef] [PubMed]
- Inal, J.M.; Schifferli, J.A. Complement C2 Receptor Inhibitor Trispanning and the β-Chain of C4 Share a Binding Site for Complement C2. J. Immunol. 2002, 168, 5213–5221. [Google Scholar] [CrossRef] [PubMed]
- Henney, C.S.; Mayer, M.M. Specific cytolytic activity of lymphocytes: Effect of antibodies against complement components c2, c3, and c5. Cell. Immunol. 1971, 2, 705. [Google Scholar] [CrossRef]
- Rawal, N.; Pangburn, M.K. Structure/function of C5 convertases of complement. Int. Immunopharmacol. 2001, 1, 415–422. [Google Scholar] [CrossRef]
- Taylor, F.; Chang, A.; Ferrell, G.; Mather, T.; Catlett, R.; Blick, K.; Esmon, C.T. C4b-binding protein exacerbates the host response to Escherichia coli. Blood 1991, 78, 357–363. [Google Scholar] [CrossRef]
- Koopman, W.J.; Sandberg, A.L.; Wahl, S.M.; Mergenhagen, S.E. Interaction of soluble C3 fragments with guinea pig lymphocytes. Comparison of effects of C3a, C3b, C3c, and C3d on lymphokine production and lymphocyte proliferation. J. Immunol. 1976, 117, 331–336. [Google Scholar]
- Barber, D.F.; Faure, M.; Long, E.O. LFA-1 contributes an early signal for NK cell cytotoxicity. J. Immunol. 2004, 173, 3653–3659. [Google Scholar] [CrossRef]
- Ostermann, G.; Weber, K.S.C.; Zernecke, A.; Schröder, A.; Weber, C. JAM-1 is a ligand of the β2 integrin LFA-1 involved in transendothelial migration of leukocytes. Nat. Immunol. 2002, 3, 151–158. [Google Scholar] [CrossRef]
- Bai, M.; Grieshaber-Bouyer, R.; Wang, J.; Schmider, A.B.; Wilson, Z.S.; Zeng, L.; Halyabar, O.; Godin, M.D.; Nguyen, H.N.; Levescot, A.; et al. CD177 modulates human neutrophil migration through activation-mediated integrin and chemoreceptor regulation. Blood 2017, 130, 2092–2100. [Google Scholar] [CrossRef]
- Kristóf, E.; Zahuczky, G.; Katona, K.; Doró, Z.; Nagy, É.; Fésüs, L. Novel role of icam3 and lfa-1 in the clearance of apoptotic neutrophils by human macrophages. Apoptosis 2013, 18, 1235–1251. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pan, X.-J.; Xie, L.-X. Expression of integrin during the course of adhesion between fungi and corneal epithelial cells. [Zhonghua yan ke za zhi] Chin. J. Ophthalmol. 2010, 46, 834. [Google Scholar]
- Cornacchione, P.; Scaringi, L.; Fettucciari, K.; Rosati, E.; Sabatini, R.; Orefici, G.; van Hunolstein, C.; Modesti, A.; Modica, A.; Minelli, F.; et al. Group B streptococci persist inside macrophages. Immunology 1998, 93, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Doran, K.S.; Lawrence, T.; Turkson, N.; Puliti, M.; Tissi, L.; Nizet, V. Sword and shield: Linked group B streptococcal β-hemolysin/cytolysin and carotenoid pigment function to subvert host phagocyte defense. Proc. Natl. Acad. Sci. USA 2004, 101, 14491–14496. [Google Scholar] [CrossRef] [PubMed]
- Valenti-Weigand, P.; Benkel, P.; Rohde, M.; Chhatwal, G.S. Entry and intracellular survival of group B streptococci in J774 macrophages. Infect. Immun. 1996, 64, 2467–2473. [Google Scholar]
- Barbano, G.; Cappa, F.; Prigione, I.; Tedesco, F.; Pausa, M.; Gugliemino, R.; Pistoia, V.; Gusmano, R.; Perfumo, F. Peritoneal mesothelial cells produce complement factors and express CD59 that inhibits C5b-9-mediated cell lysis. Adv. Perit. Dial. Conf. Perit. Dial. 1999, 15, 253–257. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Description | YM001-Brain Relative Expression Level | HN016-Brain Relative Expression Level |
|---|---|---|---|
| XP_003451887.1 | collagen alpha-1(II) chain isoform X1 | 1.524 | 2.9755 |
| XP_003439010.1 | collagen alpha-1(II) chain isoform X1 | 1.332 | 2.451 |
| XP_003438858.1 | collagen alpha-3(IX) chain isoform X1 | 0.8685 | 1.4765 |
| XP_005461453.1 | collagen alpha-1(XI) chain | 0.8435 | 1.2605 |
| XP_005459969.3 | vitellogenin | 0.572 | 1.0565 |
| XP_003452622.2 | vitellogenin-2 | 0.545 | 1.0055 |
| XP_003447299.1 | apolipoprotein A-I | 0.5 | 0.9025 |
| XP_003453341.1 | leukocyte cell-derived chemotaxin 1 | 1.387 | 2.254 |
| XP_003459588.2 | complement C1q tumor necrosis factor-related protein 3 | 0.389 | 0.643 |
| XP_003444152.1 | type-4 ice-structuring protein | 0.5765 | 0.948 |
| XP_019213314.1 | hemoglobin subunit beta-A | 0.3215 | 0.4815 |
| XP_025755569.1 | uncharacterized protein LOC100702986 | 2.3545 | 3.593 |
| XP_003450540.1 | apolipoprotein A-I | 0.668 | 1.04 |
| XP_025755734.1 | serotransferrin-like isoform X1 | 0.7645 | 1.1855 |
| NP_001298250.1 | serotransferrin-like precursor | 0.662 | 1.0275 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Li, L.; Huang, T.; Wu, W.; Liang, W.; Chen, M. The Interaction between Phagocytes and Streptococcus agalactiae (GBS) Mediated by the Activated Complement System is the Key to GBS Inducing Acute Bacterial Meningitis of Tilapia. Animals 2019, 9, 818. https://doi.org/10.3390/ani9100818
Liu Y, Li L, Huang T, Wu W, Liang W, Chen M. The Interaction between Phagocytes and Streptococcus agalactiae (GBS) Mediated by the Activated Complement System is the Key to GBS Inducing Acute Bacterial Meningitis of Tilapia. Animals. 2019; 9(10):818. https://doi.org/10.3390/ani9100818
Chicago/Turabian StyleLiu, Yu, Liping Li, Ting Huang, Wende Wu, Wanwen Liang, and Ming Chen. 2019. "The Interaction between Phagocytes and Streptococcus agalactiae (GBS) Mediated by the Activated Complement System is the Key to GBS Inducing Acute Bacterial Meningitis of Tilapia" Animals 9, no. 10: 818. https://doi.org/10.3390/ani9100818
APA StyleLiu, Y., Li, L., Huang, T., Wu, W., Liang, W., & Chen, M. (2019). The Interaction between Phagocytes and Streptococcus agalactiae (GBS) Mediated by the Activated Complement System is the Key to GBS Inducing Acute Bacterial Meningitis of Tilapia. Animals, 9(10), 818. https://doi.org/10.3390/ani9100818