Effect of Curcumin Supplement in Summer Diet on Blood Metabolites, Antioxidant Status, Immune Response, and Testicular Gene Expression in Hu Sheep

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materal and Methods
2.1. Animals and Chemicals
2.2. Experimental Design
2.2.1. Body and Testis Weight
2.2.2. Serum Biochemical Parameters
2.2.3. Serum Activity of SOD and GPX
2.2.4. The Plasma Concentration of IgA, IgM, and IgG
2.2.5. Quantitative Real-Time PCR
2.2.6. Plasma Testosterone Concentration
2.2.7. Statistical Analysis
3. Result
3.1. Serum Biochemical Parameters
3.2. Serum SOD and GPX activity
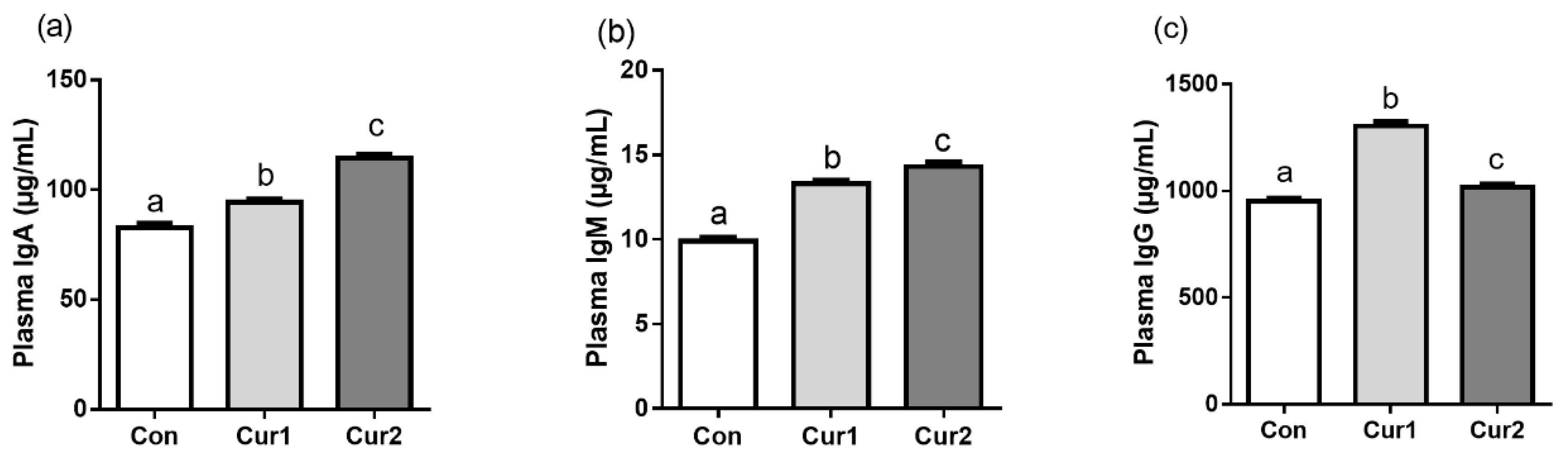
3.3. Plasma Concentration of IgA, IgM, and IgG
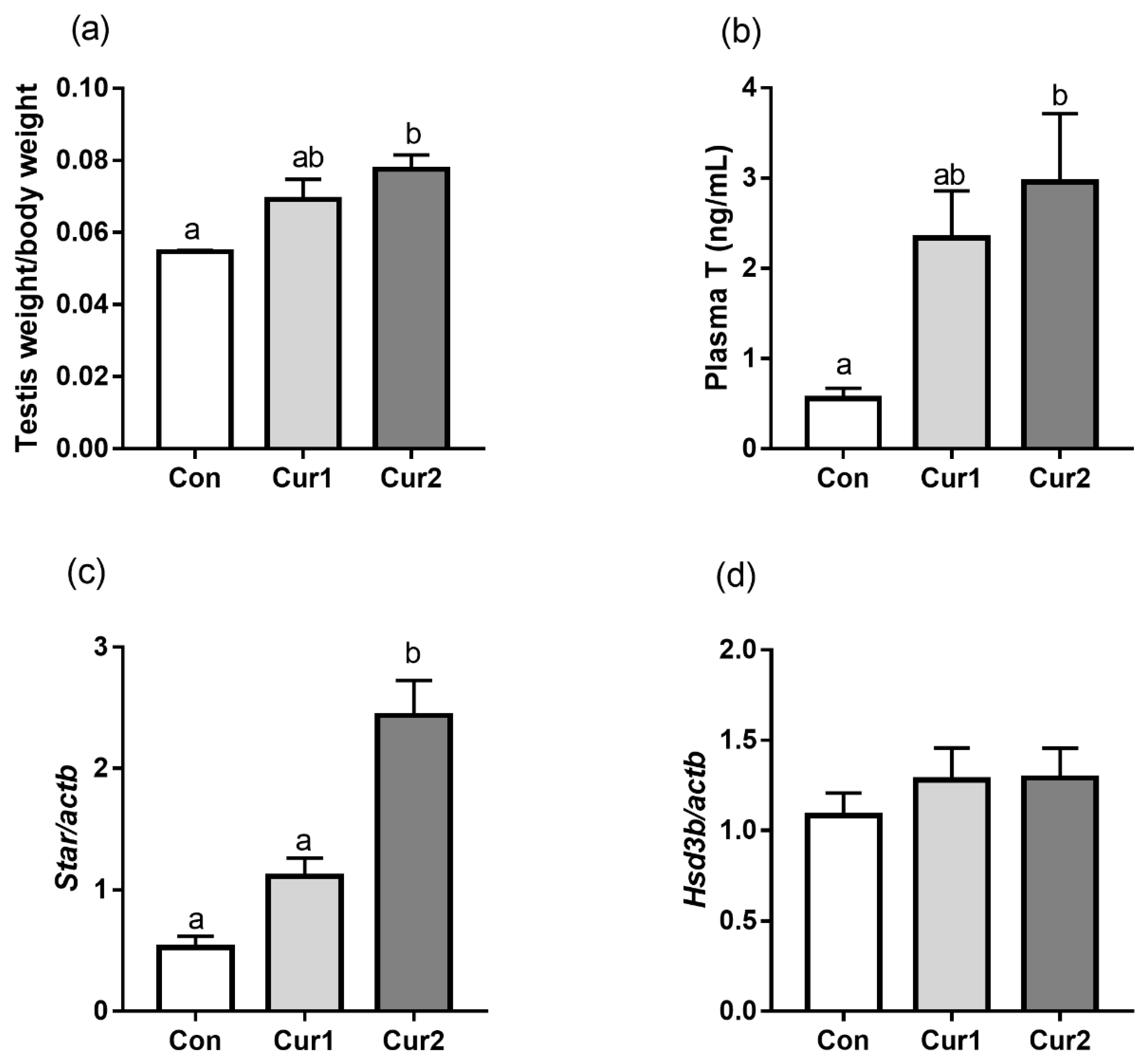
3.4. Testis Weight/Body Weight, Testosterone Concentration and Androgen-Related Gene Expressions
3.5. Expression Patterns of Apoptosis-Related Genes in Testis
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Date Availability Statement
References
- Belhadj Slimen, I.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.W.; Hale, B.J.; Seibert, J.T.; Romoser, M.R.; Adur, M.K.; Keating, A.F.; Baumgard, L.H. Physiological mechanisms through which heat stress compromises reproduction in pigs. Mol. Reprod. Dev. 2017, 84, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.; Xu, L.; Drake, V.; Oberley, L.; Oberley, T.; Moseley, P.; Kregel, K. Aging reduces adaptive capacity and stress protein expression in the liver after heat stress. J. Appl. Physiol. 2000, 89, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Victoria Sanz Fernandez, M.; Johnson, J.S.; Abuajamieh, M.; Stoakes, S.K.; Seibert, J.T.; Cox, L.; Kahl, S.; Elsasser, T.H.; Ross, J.W.; Clay Isom, S. Effects of heat stress on carbohydrate and lipid metabolism in growing pigs. Physiol. Rep. 2015, 3, e12315. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, J.; Peng, D.; Li, G.; Chen, J.; Gu, X. Exposure to heat-stress environment affects the physiology, circulation levels of cytokines, and microbiome in dairy cows. Sci. Rep. 2018, 8, 14606. [Google Scholar] [CrossRef] [PubMed]
- Hicks, T.A.; McGlone, J.J.; Whisnant, C.S.; Kattesh, H.G.; Norman, R.L. Behavioral, endocrine, immune, and performance measures for pigs exposed to acute stress. Sci. J. Anim. Sci. 1998, 76, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Mashaly, M.; Hendricks 3rd, G.; Kalama, M.; Gehad, A.; Abbas, A.; Patterson, P. Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult. Sci. J. 2004, 83, 889–894. [Google Scholar] [CrossRef]
- Tao, S.; Monteiro, A.; Thompson, I.; Hayen, M.; Dahl, G. Effect of late-gestation maternal heat stress on growth and immune function of dairy calves. J. Dairy Sci. 2012, 95, 7128–7136. [Google Scholar] [CrossRef]
- Zulkifli, I.; Che Norma, M.; Israf, D.; Omar, A. The effect of early age feed restriction on subsequent response to high environmental temperatures in female broiler chickens. Poult. Sci. J. 2000, 79, 1401–1407. [Google Scholar] [CrossRef]
- Nisar, A.; Sultana, M.; Ashraf, H. Oxidative stress-threat to animal health and production. Int. J. Livest. Res. 2013, 3, 76–83. [Google Scholar]
- Cong, X.; Zhang, Q.; Li, H.; Jiang, Z.; Cao, R.; Gao, S.; Tian, W. Puerarin ameliorates heat stress–induced oxidative damage and apoptosis in bovine Sertoli cells by suppressing ROS production and upregulating Hsp72 expression. Theriogenology 2017, 88, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Kantidze, O.; Velichko, A.; Luzhin, A.; Razin, S. Heat stress-induced DNA damage. Acta Nat. 2016, 8. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.M.; Gomes, A.; Pinheiro, M.L.; Ribeiro, A.; Ferraz-de-Paula, V.; Astolfi-Ferreira, C.S.; Ferreira, A.J.P.; Palermo-Neto, J. Heat stress impairs performance and induces intestinal inflammation in broiler chickens infected with Salmonella Enteritidis. Avian Pathol. 2012, 41, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Caroprese, M.; Ciliberti, M.G.; Annicchiarico, G.; Albenzio, M.; Muscio, A.; Sevi, A. Hypothalamic-pituitary-adrenal axis activation and immune regulation in heat-stressed sheep after supplementation with polyunsaturated fatty acids. J. Dairy Sci. 2014, 97, 4247–4258. [Google Scholar] [CrossRef] [PubMed]
- Maurya, V.; Sejian, V.; Kumar, D.; Naqvi, S. Impact of heat stress, nutritional restriction and combined stresses (heat and nutritional) on growth and reproductive performance of Malpura rams under semi-arid tropical environment. J. Anim. Physiol. Anim. Nutr. 2016, 100, 938–946. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.M.; Rodrigues, M.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.; Sá, L.R.M.d.; Ferreira, A.J.P.; Palermo-Neto, J. Acute heat stress impairs performance parameters and induces mild intestinal enteritis in broiler chickens: Role of acute hypothalamic-pituitary-adrenal axis activation. J. Anim. Sci. 2012, 90, 1986–1994. [Google Scholar] [CrossRef] [PubMed]
- Akhavan-Salamat, H.; Ghasemi, H.A. Alleviation of chronic heat stress in broilers by dietary supplementation of betaine and turmeric rhizome powder: Dynamics of performance, leukocyte profile, humoral immunity, and antioxidant status. Trop. Anim. Health Prod. 2016, 48, 181–188. [Google Scholar] [CrossRef]
- He, S.; Zhao, S.; Dai, S.; Liu, D.; Bokhari, S.G. Effects of dietary betaine on growth performance, fat deposition and serum lipids in broilers subjected to chronic heat stress. J. Anim. Sci. 2015, 86, 897–903. [Google Scholar] [CrossRef]
- Hosseini-Vashan, S.; Golian, A.; Yaghobfar, A. Growth, immune, antioxidant, and bone responses of heat stress-exposed broilers fed diets supplemented with tomato pomace. Int. J. Biometeorol. 2016, 60, 1183–1192. [Google Scholar] [CrossRef]
- Wan, X.; Zhang, J.; He, J.; Bai, K.; Zhang, L.; Wang, T. Dietary enzymatically treated Artemisia annua L. supplementation alleviates liver oxidative injury of broilers reared under high ambient temperature. Int. J. Biometeorol. 2017, 61, 1629–1636. [Google Scholar] [CrossRef]
- Agrawal, D.K.; Mishra, P.K. Curcumin and its analogues: Potential anticancer agents. Med. Res. Rev. 2010, 30, 818–860. [Google Scholar] [PubMed]
- Jin, W.; Wang, J.; Zhu, T.; Yuan, B.; Ni, H.; Jiang, J.; Wang, H.; Liang, W. Anti-inflammatory effects of curcumin in experimental spinal cord injury in rats. Inflamm. Res. 2014, 63, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Mosley, C.A.; Liotta, D.C.; Snyder, J.P. Highly active anticancer curcumin analogues. In The Molecular Targets and Therapeutic Uses of Curcumin in Health and Disease; Springer: Berlin, Germany, 2007; pp. 77–103. [Google Scholar]
- Wongcharoen, W.; Phrommintikul, A. The protective role of curcumin in cardiovascular diseases. Int. J. Cardiol. 2009, 133, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Jaguezeski, A.M.; Perin, G.; Crecencio, R.B.; Baldissera, M.D.; Stefanil, L.M.; da Silva, A.S. Addition of Curcumin in Dairy Sheep Diet in the Control of Subclinical Mastitis. Acta Sci. Vet. 2018, 46, 7. [Google Scholar] [CrossRef]
- Bulku, E.; Stohs, S.J.; Cicero, L.; Brooks, T.; Halley, H.; Ray, S.D. Curcumin exposure modulates multiple pro-apoptotic and anti-apoptotic signaling pathways to antagonize acetaminophen-induced toxicity. Curr. Neurovasc. Res. 2012, 9, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, F. Effect of turmeric (Curcumin longa) powder on performance, oxidative stress state and some of blood parameters in broiler fed on diets containing aflatoxin B1. Glob. Vet. 2010, 5, 312–317. [Google Scholar]
- Emadi, M.; Kermanshahi, H. Effect of turmeric rhizome powder on the activity of some blood enzymes in broiler chickens. Int. J. Poult. Sci. 2007, 6, 48–51. [Google Scholar]
- Riasi, A.; Kermanshahi, H.; Fathi, M. Effect of Turmeric rhizome powder (Curcuma longa) on performance, egg quality and some blood serum parameters of laying hens. In Proceedings of the 1st Mediterranean Summit of World Poultry Science Association, Chalkidiki, Greece, 7–10 May 2008. [Google Scholar]
- Molosse, V.; Souza, C.F.; Baldissera, M.D.; Glombowsky, P.; Campigotto, G.; Cazaratto, C.J.; Stefani, L.M.; da Silva, A.S. Diet supplemented with curcumin for nursing lambs improves animal growth, energetic metabolism, and performance of the antioxidant and immune systems. Small Rumin. Res. 2019, 170, 74–81. [Google Scholar] [CrossRef]
- Geng, R.-Q.; Chang, H.; Yang, Z.-P.; Sun, W.; Wang, L.-P.; Lu, S.-X.; Tsunoda, K.; Ren, Z. Study on origin and phylogeny status of Hu sheep. Asian-Australas. J. Anim. Sci. 2003, 16, 743–747. [Google Scholar] [CrossRef]
- DiGiacomo, K.; Simpson, S.; Leury, B.; Dunshea, F. Dietary betaine impacts the physiological responses to moderate heat conditions in a dose dependent manner in sheep. Animals 2016, 6, 51. [Google Scholar] [CrossRef]
- Liu, H.; Li, K.; Mingbin, L.; Zhao, J.; Xiong, B. Effects of chestnut tannins on the meat quality, welfare, and antioxidant status of heat-stressed lambs. Meat Sci. 2016, 116, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Alhidary, I.; Shini, S.; Al Jassim, R.; Gaughan, J. Physiological responses of Australian Merino wethers exposed to high heat load. J. Anim. Sci. 2012, 90, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Shen, G.; Wang, Y.; Wu, C. Curcumin supplementation regulates lipid metabolism in broiler chickens. Poult. Sci. 2018, 98, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.K.; Ha, T.Y.; McGregor, R.A.; Choi, M.S. Long-term curcumin administration protects against atherosclerosis via hepatic regulation of lipoprotein cholesterol metabolism. Mol. Nutr. Food Res. 2011, 55, 1829–1840. [Google Scholar] [CrossRef] [PubMed]
- Zingg, J.M.; Hasan, S.T.; Meydani, M. Molecular mechanisms of hypolipidemic effects of curcumin. Biofactors 2013, 39, 101–121. [Google Scholar] [CrossRef] [PubMed]
- Hajati, H.; Hassanabadi, A.; Golian, A.; Nassiri-Moghaddam, H.; Nassiri, M.R. The effect of grape seed extract and vitamin C feed supplementation on some blood parameters and HSP70 gene expression of broiler chickens suffering from chronic heat stress. Ital. J. Anim. Sci. 2015, 14, 3273. [Google Scholar] [CrossRef]
- Jaguezeski, A.M.; Gündel, S.S.; Favarin, F.R.; Gündel, A.; Souza, C.F.; Baldissera, M.D.; Cazarotto, C.C.; Volpato, A.; Fortuoso, B.F.; Ourique, A.F. Low-dose curcumin-loaded Eudragit L-100-nanocapsules in the diet of dairy sheep increases antioxidant levels and reduces lipid peroxidation in milk. J. Food Eng. 2019, e12942. [Google Scholar] [CrossRef]
- Lin, H.; Decuypere, E.; Buyse, J. Acute heat stress induces oxidative stress in broiler chickens. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2006, 144, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Bauché, F.; Fouchard, M.-H.; Jégou, B. Antioxidant system in rat testicular cells. FEBS Lett. 1994, 349, 392–396. [Google Scholar] [CrossRef]
- Li, Y.; Cao, Y.; Wang, F.; Li, C. Scrotal heat induced the Nrf2-driven antioxidant response during oxidative stress and apoptosis in the mouse testis. Acta. Histochem. 2014, 116, 883–890. [Google Scholar] [CrossRef]
- Zhang, J.; Bai, K.; Su, W.; Wang, A.; Zhang, L.; Huang, K.; Wang, T. Curcumin attenuates heat-stress-induced oxidant damage by simultaneous activation of GSH-related antioxidant enzymes and Nrf2-mediated phase II detoxifying enzyme systems in broiler chickens. Poult. Sci. 2018, 97, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Jaguezeski, A.M.; Perin, G.; Bottari, N.B.; Wagner, R.; Fagundes, M.B.; Schetinger, M.R.C.; Morsch, V.M.; Stein, C.S.; Moresco, R.N.; Barreta, D.A. Addition of curcumin to the diet of dairy sheep improves health, performance and milk quality. Anim. Feed Sci. Tech. 2018, 246, 144–157. [Google Scholar] [CrossRef]
- Cao, D.; Li, H.; Yi, J.; Zhang, J.; Che, H.; Cao, J.; Yang, L.; Zhu, C.; Jiang, W. Antioxidant properties of the mung bean flavonoids on alleviating heat stress. PLoS ONE 2011, 6, e21071. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; He, J.; Xie, H.; Yang, Y.; Li, J.; Zou, Y. Resveratrol induces antioxidant and heat shock protein mRNA expression in response to heat stress in black-boned chickens. Poult. Sci 2013, 93, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, A.; Upadhyay, R. Heat stress and immune function. In Heat Stress and Animal Productivity; Springer: Berlin, Germany, 2013; pp. 113–136. [Google Scholar]
- Min, L.; Zheng, N.; Zhao, S.; Cheng, J.; Yang, Y.; Zhang, Y.; Yang, H.; Wang, J. Long-term heat stress induces the inflammatory response in dairy cows revealed by plasma proteome analysis. Biochem. Biophys. Res. Commun. 2016, 471, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Dong, X.; Tong, J.; Zhang, Q. The probiotic Bacillus licheniformis ameliorates heat stress-induced impairment of egg production, gut morphology, and intestinal mucosal immunity in laying hens. Poult. Sci. 2012, 91, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Smith, M. Effects of different levels of zinc on the performance and immunocompetence of broilers under heat stress. Poult. Sci. 2003, 82, 1580–1588. [Google Scholar]
- Liu, D.Y.; He, S.J.; Liu, S.Q.; Tang, Y.G.; Jin, E.H.; Chen, H.L.; Li, S.H.; Zhong, L.T. Daidzein enhances immune function in late lactation cows under heat stress. Anim. Sci. J. 2014, 85, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Çiftçi, O. Curcumin prevents toxic effects of 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin (TCDD) on humoral immunity in rats. Food Agr. Immunol. 2011, 22, 31–38. [Google Scholar] [CrossRef]
- Krishnaraju, A.; Sundararaju, D.; Sengupta, K.; Venkateswarlu, S.; Trimurtulu, G. Safety and toxicological evaluation of demethylatedcurcuminoids; a novel standardized curcumin product. Toxicol. Mech. Methods 2009, 19, 447–460. [Google Scholar] [CrossRef]
- Dandekar, P.; Dhumal, R.; Jain, R.; Tiwari, D.; Vanage, G.; Patravale, V. Toxicological evaluation of pH-sensitive nanoparticles of curcumin: Acute, sub-acute and genotoxicity studies. Food Chem. Toxicol. 2010, 48, 2073–2089. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Shin, D.-G.; Park, S.G.; Chu, S.B.; Gwon, L.W.; Lee, J.-G.; Yon, J.-M.; Baek, I.-J.; Nam, S.-Y. Curcumin dose-dependently improves spermatogenic disorders induced by scrotal heat stress in mice. Food Funct. 2015, 6, 3770–3777. [Google Scholar]
- Juan, M.E.; Gonzalez-Pons, E.; Munuera, T.; Ballester, J.; Rodriguez-Gil, J.E.; Planas, J.M. trans-Resveratrol, a natural antioxidant from grapes, increases sperm output in healthy rats. J. Nutr. 2005, 135, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Hailing, L.; Hui, M.; Guijie, Z. Effect of vitamin E supplementation on development of reproductive organs in Boer goat. Anim. Reprod. Sci. 2009, 113, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Aktas, C.; Kanter, M. A morphological study on Leydig cells of scrotal hyperthermia applied rats in short-term. J. Mol. Histol. 2009, 40, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Stocco, D.M.; Wang, X.; Jo, Y.; Manna, P.R. Multiple signaling pathways regulating steroidogenesis and steroidogenic acute regulatory protein expression: more complicated than we thought. Mol. Endocrinol. 2005, 19, 2647–2659. [Google Scholar] [CrossRef] [PubMed]
- Rizk, S.M.; Zaki, H.F.; Mina, M.A. Propolis attenuates doxorubicin-induced testicular toxicity in rats. Food. Chem. Toxicol. 2014, 67, 176–186. [Google Scholar] [CrossRef]
- Bozkaya, F.; Atli, M.; Guzeloglu, A.; Kayis, S.; Yildirim, M.; Kurar, E.; Yilmaz, R.; Aydilek, N. Effects of long-term heat stress and dietary restriction on the expression of genes of steroidogenic pathway and small heat-shock proteins in rat testicular tissue. Andrologia 2017, 49, e12668. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Choi, Y.S.; Park, S.G.; Gwon, L.W.; Lee, J.G.; Yon, J.-M.; Baek, I.-J.; Lee, B.J.; Yun, Y.W.; Nam, S.-Y. Enhanced protective effects of combined treatment with β-carotene and curcumin against hyperthermic spermatogenic disorders in mice. Biomed. Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Cormier, M.; Ghouili, F.; Roumaud, P.; Bauer, W.; Touaibia, M.; Martin, L.J. Influences of flavones on cell viability and cAMP-dependent steroidogenic gene regulation in MA-10 Leydig cells. Cell Biol. Toxicol. 2018, 34, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Cormier, M.; Ghouili, F.; Roumaud, P.; Martin, L.J.; Touaibia, M. Influence of flavonols and quercetin derivative compounds on MA-10 Leydig cells steroidogenic genes expressions. Toxicol. in Vitro 2017, 44, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Kanter, M.; Aktas, C.; Erboga, M. Heat stress decreases testicular germ cell proliferation and increases apoptosis in short term: an immunohistochemical and ultrastructural study. Toxicol. Ind. Health. 2013, 29, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Paul, C.; Teng, S.; Saunders, P.T. A single, mild, transient scrotal heat stress causes hypoxia and oxidative stress in mouse testes, which induces germ cell death. Biol. Reprod. 2009, 80, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Xi, H.; Zhang, Z.; Liang, Y.; Li, Q.; He, J. Germ cell apoptosis and expression of Bcl-2 and Bax in porcine testis under normal and heat stress conditions. Acta Histochem. 2017, 119, 198–204. [Google Scholar] [CrossRef]
- Tan, Q.; An, C.; Xiao, Y.; Liao, Z. Protective effect of lycium barbarum polysaccharides against heat stress-induced germ cell apoptosis in rats and its mechanism. Zhonghua Nan Ke Xue Natl. J. Androl. 2012, 18, 88–92. [Google Scholar] [PubMed]
- Kanter, M.; Aktas, C.; Erboga, M. Curcumin attenuates testicular damage, apoptotic germ cell death, and oxidative stress in streptozotocin-induced diabetic rats. Mol. Nutr. Food Res. 2013, 57, 1578–1585. [Google Scholar] [CrossRef]
- Verma, R.J.; Mathuria, N. Effect of curcumin on aflatoxin-induced biochemical changes in testis of mice. Fertil. Steril. 2009, 91, 597–601. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Content/% | Item | Content |
|---|---|---|---|
| Concentrate | |||
| Corn | 51.22 | DE (MJ/Kg) | 12.81 |
| Wheat bran | 17.41 | CP (g/Kg) | 171.73 |
| Barley bran | 15.91 | Ca/% | 0.85 |
| Sesame | 4.98 | P/% | 0.66 |
| Premix | 6.46 | ||
| NaHCO3 | 2.48 | ||
| Salt | 1.49 | ||
| NaSeO3 | 0.05 | ||
| Total | 100 | ||
| Roughage | |||
| Silage | 52.24 | DE (MJ/Kg) | 3.79 |
| Hay | 29.85 | CP (g/Kg) | 49 |
| Soybean milk residue | 17.91 | Ca/% | 0.4 |
| Total | 100 | P/% | 0.07 |
| Gene | Primer Sequence (5′-3′) | Product Size (bp) | GeneBank Accession Number |
|---|---|---|---|
| star | F: GGGCATCCTCAAAGACCAG | 120 | NM_001009243.1 |
| R: TCCACCACCACCTCCAAC | |||
| hsd3b | F: ATCCACACCAGCACCATAG | 144 | NM_001135932.1 |
| R: TTCCAGCACAGCCTTCTC | |||
| bcl-2 | F: CGCATCGTGGCCTTCTTT | 113 | XM_027960877.1 |
| R: CGGTTCAGGTACTCGGTCATC | |||
| caspase-3 | F: TCAGGGAAACCTTCACGAGC | 274 | XM_027962551.1 |
| R: CCTCGGCAGGCCTGAATAAT | |||
| actb | F: CCAAGGCCAACCGTGAGAAG | 349 | NM_001009784.3 |
| R: CCATCTCCTGCTTCGAAGTCC |
| Item | Con | Cur1 | Cur2 |
|---|---|---|---|
| Glu (mmol/L) | 3.37 + 0.81 | 3.30 + 0.89 | 3.81 + 1.17 |
| TC (mmol/L) | 3.00 + 0.43 | 3.26 + 0.36 | 3.30 + 0.52 |
| TG (mmol/L) | 0.31 + 0.08 | 0.33 + 0.06 | 0.34 + 0.08 |
| NEFA (mmol/L) | 0.26 + 0.07 a | 0.30 + 0.08 ab | 0.36 + 0.09 b |
| HDL (mmol/L) | 0.85 + 0.14 | 0.88+0.11 | 0.87 + 0.10 |
| LDL (mmol/L) | 0.71 + 0.11 | 0.73 + 0.15 | 0.71 + 0.16 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Z.; Wan, Y.; Li, P.; Xue, Y.; Cui, W.; Chen, Q.; Chen, J.; Wang, F.; Mao, D. Effect of Curcumin Supplement in Summer Diet on Blood Metabolites, Antioxidant Status, Immune Response, and Testicular Gene Expression in Hu Sheep. Animals 2019, 9, 720. https://doi.org/10.3390/ani9100720
Jiang Z, Wan Y, Li P, Xue Y, Cui W, Chen Q, Chen J, Wang F, Mao D. Effect of Curcumin Supplement in Summer Diet on Blood Metabolites, Antioxidant Status, Immune Response, and Testicular Gene Expression in Hu Sheep. Animals. 2019; 9(10):720. https://doi.org/10.3390/ani9100720
Chicago/Turabian StyleJiang, Zhiyang, Yongjie Wan, Peng Li, Yang Xue, Wenwen Cui, Qi Chen, Jianqin Chen, Feng Wang, and Dagan Mao. 2019. "Effect of Curcumin Supplement in Summer Diet on Blood Metabolites, Antioxidant Status, Immune Response, and Testicular Gene Expression in Hu Sheep" Animals 9, no. 10: 720. https://doi.org/10.3390/ani9100720
APA StyleJiang, Z., Wan, Y., Li, P., Xue, Y., Cui, W., Chen, Q., Chen, J., Wang, F., & Mao, D. (2019). Effect of Curcumin Supplement in Summer Diet on Blood Metabolites, Antioxidant Status, Immune Response, and Testicular Gene Expression in Hu Sheep. Animals, 9(10), 720. https://doi.org/10.3390/ani9100720