Observation of the Breeding Behavior of the Chinese Giant Salamander (Andrias davidianus) Using a Digital Monitoring System


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Digital Monitoring System
2.3. Observation Index
2.4. Data Processing
3. Results
3.1. Sand-Pushing
3.2. Showering Behavior
3.3. Courtship Behavior
3.4. Oviposition
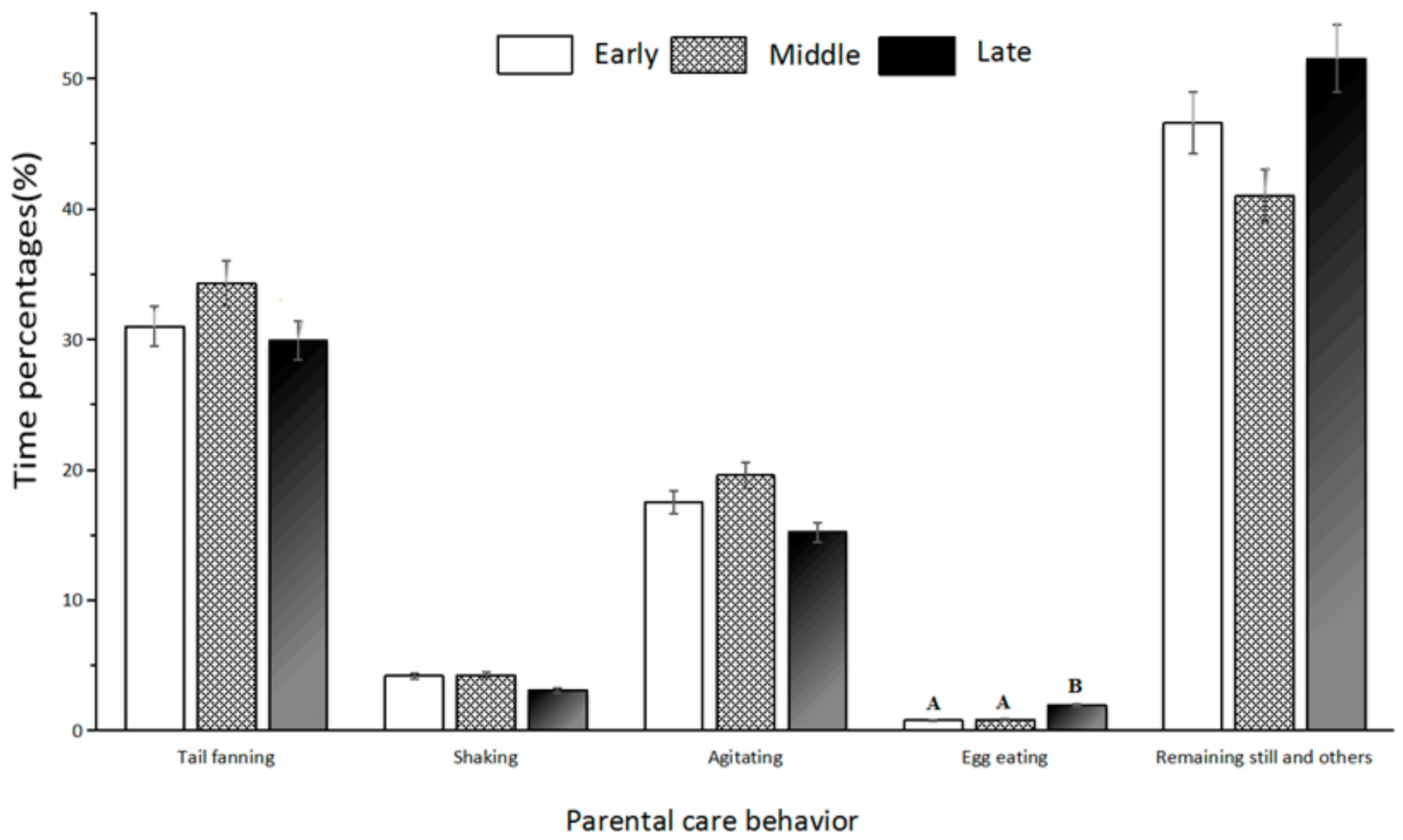
3.5. Parental Care
4. Discussion
4.1. Sand-Pushing
4.2. Showering Behavior
4.3. Courtship
4.4. Oviposition
4.5. Parental care
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Number | Total Length (m) | SVL | Weight (kg) | Age | Mating Success | |
|---|---|---|---|---|---|---|
| Partner A | Male A | 1.08 | 0.68 | 8.41 | 9 | Yes |
| Female A | 1.02 | 0.64 | 8.23 | 9 | Yes | |
| Partner B | Male B | 1.01 | 0.63 | 7.81 | 8 | Yes |
| Female B | 1.06 | 0.66 | 8.34 | 9 | Yes | |
| Partner C | Male C | 0.96 | 0.60 | 7.78 | 8 | No |
| Female C | 1.05 | 0.66 | 8.25 | 9 | No | |
| Partner D | Male D | 0.91 | 0.57 | 5.86 | 8 | No |
| Female D | 0.95 | 0.60 | 6.37 | 8 | No | |
References
- Murphy, R.W.; Fu, J.Z.; Upton, D.E.; De, L.T.; Zhao, E.M. Genetic variability among endangered Chinese giant salamanders, Andrias davidianus. Mol. Ecol. 2000, 9, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Roelants, K.; Gower, D.J.; Wilkinson, M.; Loader, S.P.; Biju, S.D.; Guillaume, K.; Moriau, L.; Bossuyt, F. Global patterns of diversification in the history of modern amphibians. Proc. Natl. Acad. Sci. USA 2007, 104, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Tominaga, A.; Liu, W.Z.; T-U, T. Reduced genetic variation in the Japanese giant salamander, Andrias japonicus (Amphibia: Caudata). Mol. Phylogenet. Evol. 2008, 49, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Ijspeert, A.J.; Crespi, A.; Ryczko, D.; Cabelguen, J.-M. From swimming to walking with a salamander robot driven by a spinal cord model. Science 2007, 315, 1416–1420. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Zhang, K.J.; Wang, Z.H.; Ding, Y.Z.; Wu, W.; Huang, S. The decline of the Chinese giant salamander Andrias davidianus and implications for its conservation. Oryx 2004, 38, 197–202. [Google Scholar] [CrossRef]
- Cunningham, A.A.; Turvey, S.T.; Zhou, F.; Meredith, H.M.R. Development of the Chinese giant salamander Andrias davidianus farming industry in Shaanxi Province, China: Conservation threats and opportunities. Oryx 2016, 50, 265–273. [Google Scholar] [CrossRef]
- Petranka, J.W. Salamanders of the United States and Canada, 4th ed.; Smithsonian Institution Press: Washington, DC, USA, 2010. [Google Scholar]
- Browne, R.K.; Wang, Z.; Okada, S.; Hime, P.; McMillan, A.; Wu, M.Y.; Diaz, R.; McGinnity, D.; Briggler, J.T. The giant salamanders (Cryptobranchidae): Part B. Biogeography, ecology and reproduction. Amphib. Reptil. Conserv. 2014, 5, 35–50. [Google Scholar]
- Wang, J. Current status of Japanese giant salamander and the enlightenment on the conservation of Chinese giant salamander. Chin. J. Appl. Environ. Biol. 2015, 21, 683–688. [Google Scholar]
- Kawamichi, T.; Ueda, H. Spawning at nests of extra-large males in the giant salamander Andrias japonicus. J. Herpetol. 1998, 32, 133–136. [Google Scholar] [CrossRef]
- Okada, S.; Utsunomiya, T.; Okada, T.; Fumihiko, I. Characteristics of Japanese giant salamander (Andrias japonicus) populations in two small tributary streams in Hiroshima Prefecture, western Honshu, Japan. Herpetol. Conserv. Biol. 2008, 3, 192–202. [Google Scholar]
- Liu, B.H. Ecological survey and observation of Chinese giant salamander in Jiulongshan Moutain, Zhejiang Province. Chin. J. Wildl. 1990, 6, 12–14. [Google Scholar]
- Ge, Y.R.; Zheng, H.X. Natural breeding cycle of the giant salamander (Andrias davidianus). J. Henan Edu. Inst. 1994, 22, 67–70. [Google Scholar]
- Wang, W.L.; Jiang, F.J.; Wang, B.L. Survey the reproductive habits of Chinese giant salamander. Res. Fish 2000, 20, 12–13. [Google Scholar]
- Zhang, H.X.; Wang, K.F.; Quan, Q.Z.; Fan, W.D.; Fang, S.R. Productive ecology and behavior of Chinese giant salamander (Andrias davidianus). J. Shanxi Norm. Univ. 2006, 34, 70–75. [Google Scholar]
- Liang, G.; Wu, F. The Activity Rhythm and Reproductive Behaviors of Andrias davidianus. Chin. J. Zool. 2010, 45, 77–82. [Google Scholar]
- Xu, W.G.; Wang, Z.Q.; Liang, G. The washing behavior and its significance for male adult Andrias davidianus in the pre-reproductive period. Chin. J. Zool. 2013, 48, 529–533. [Google Scholar]
- Wang, J.; Zhang, H.X.; Xie, F.; Wei, G.; Jian, J.P. Genetic bottlenecks of the wild Chinese giant salamander in karst caves. Asian Herpetol. Res. 2017, 8, 174–183. [Google Scholar]
- Song, Y.J.; Luo, Q.H.; Zhang, L.Y.; Chen, Q.Y.; Peng, H.; Wang, H.; Ou, D.S. Ecological breeding engineering for Chinese giant salamander in Zhangjiajie city. J. Hunan Agric. Univ. 2016, 42, 635–640. [Google Scholar]
- Li, D.P.; Zhuang, P.; Yan, A.S.; Wang, X.M.; Zhang, L.Z. The influences of illumination, water current and stocking density on feeding, behavior and growth in juveniles Acipenser schrenckii. J. Fish. China 2004, 28, 54–61. [Google Scholar]
- Arnold, S.J. The Evolution of Courtship Behavior in New World Salamanders with Some Comments on Old World Salamanders. In The Reproductive Biology of Amphibians; Taylor, D.H., Guttman, S.I., Eds.; Springer: Boston, MA, USA, 1977. [Google Scholar]
- Yu, H.H. Relation between Environmental Factors and Nocturnal Active Rhythm during the Pre-Reproductive Period of the Chinese Giant Salamander; Shanxi Normal Universiy: Xi’an, China, 2013. [Google Scholar]
- Verrell, P.A. Male mate choice for fecund females in a plethodontid salamander. Anim. Behav. 1989, 38, 1086–1088. [Google Scholar] [CrossRef]
- Verrell, P.A. The sexual strategies of natural populations of newts and salamanders. Herpetologica 1989, 45, 265–282. [Google Scholar]
- Okada, S.; Fukuda, Y.; Takahashi, M.K. Paternal care behaviors of Japanese giant salamander Andrias japonicus in natural populations. J. Ethol. 2015, 33, 1–7. [Google Scholar] [CrossRef]
- Yao, Y.B.; Zhou, W.; Xiao, T.Y.; Liu, Q.L.; Liu, M. A developmental morphology characteristic comparative of Andrias davidianus embryos development. J. Hunan Univ. Arts Sci. 2013, 25, 33–39. [Google Scholar]
- Rohwer, S. Parent cannibalism of offspring and egg raiding as a courtship strategy. Am. Nat. 1978, 112, 429–440. [Google Scholar] [CrossRef]







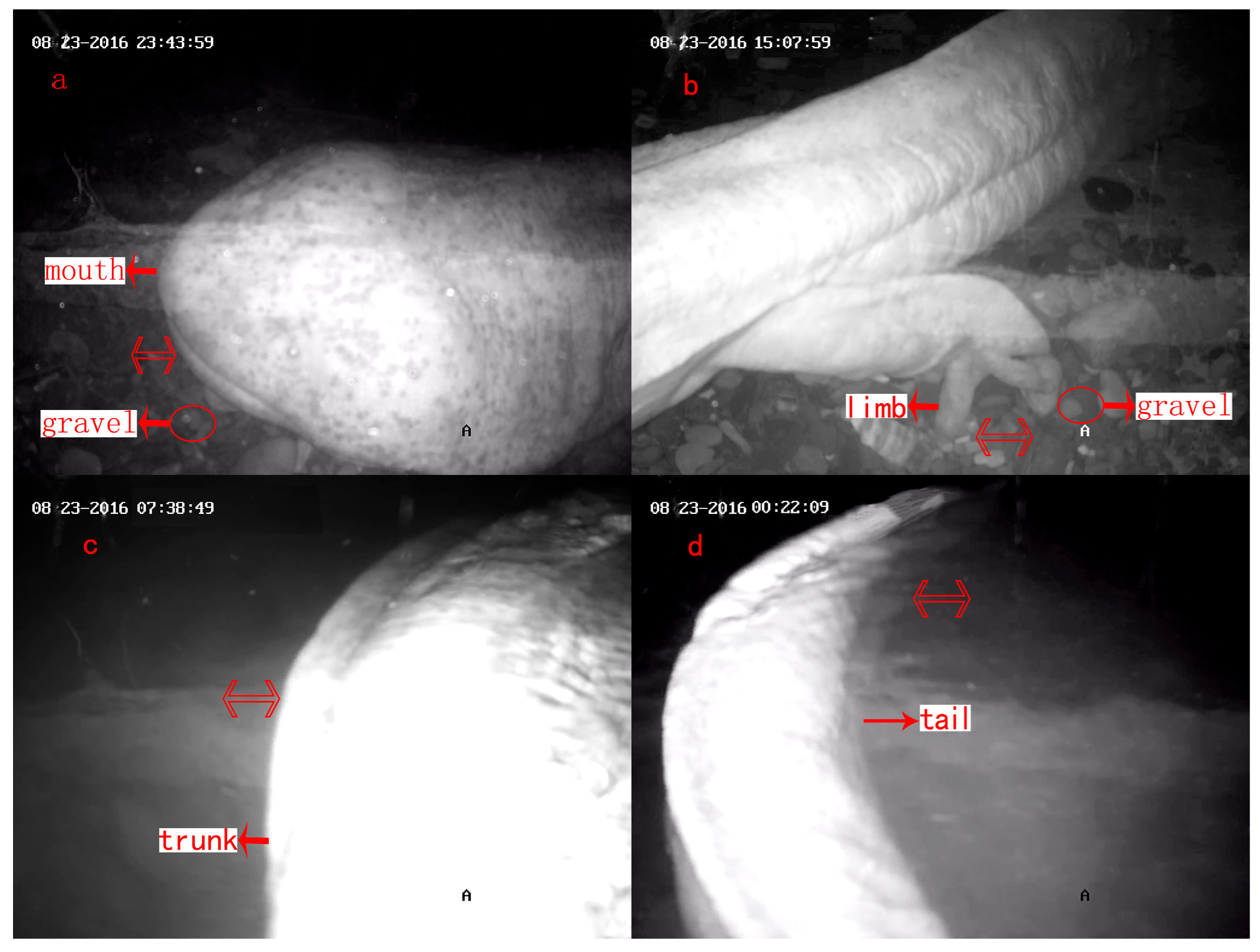
| Part of Body | Description of Posture | Duration of Each Continuous Gravel-Pushing (s) | Total Duration of Actions (h) and Coverage (%) |
|---|---|---|---|
| Mouth | Move gravel away from den with mouth (Figure 2a) | 36.8 ± 31.8 | 4.2 (6.2%) |
| Limbs | Push gravel backward with fore or hind limbs, generally with head inside the den (Figure 2b) | 58.79 ± 42.83 | 36.3 (53.8%) |
| Trunk | Scrape gravel away from body flanks with trunk (Figure 2c) | 93.0 ± 71.2 | 2.1 (3.1%) |
| Tail | Shuffle silt or sand from the bottom of the den with tail (Figure 2d) | 23.5 ± 12.4 | 24.9 (36.9%) |
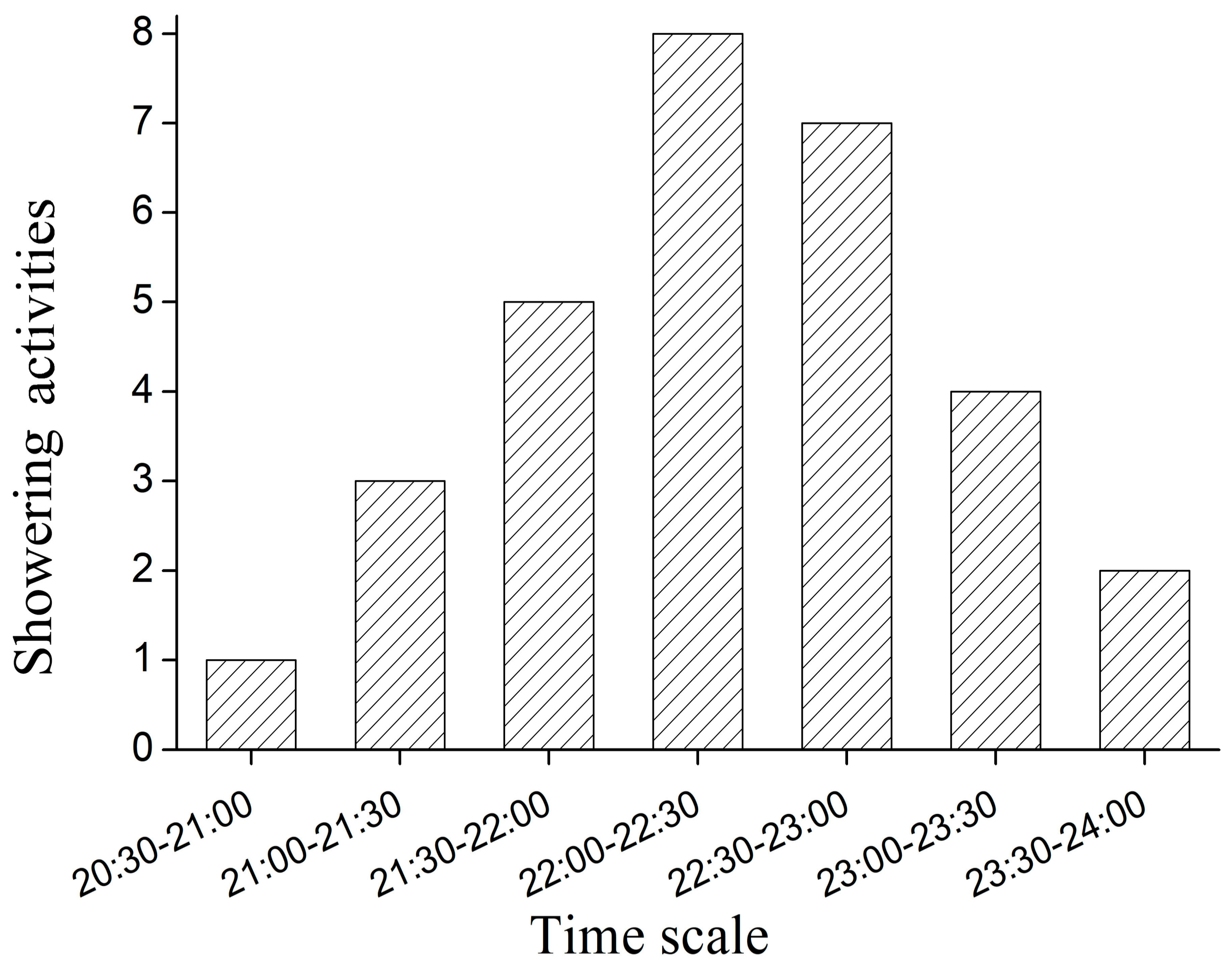
| Showering Behavior | Description | Frequency | Duration (min)/Percentage (%) |
|---|---|---|---|
| Head | Head rinsed with a swinging motion (Figure 3a) | 8 | 3.52 ± 0.44/27.5 |
| Trunk | All parts of the trunk rinsed with a swinging motion (Figure 3b) | 7 | 4.23 ± 0.52/28.9 |
| Tail | Tail rinsed with a swinging motion (Figure 3c) | 6 | 4.02 ± 0.35/23.6 |
| Curve | Body washed by bending around the water from the outlet (Figure 3d) | 5 | 2.86 ± 0.25/14.0 |
| Snout | Snout washed by moving the head out of the water (Figure 3e) | 4 | 1.55 ± 0.16/6.1 |
| Courtship Behavior | Description | Frequency Percentage (%) | Duration Percentage (%) | Duration of Each Continuous Behavior (min) |
|---|---|---|---|---|
| Side-by-side | A male and female stand side-by-side, with heads touching. | 21.58 ± 0.46 | 2.83 ± 0.46 | 2.49 ± 0.41 |
| Knocking bellies | A male knocks the belly of his female partner with his snout and his head. | 19.49 ± 0.30 | 2.61 ± 0.58 | 2.53 ± 0.48 |
| Riding | A male climbs onto a female’s trunk. or head and places his head on the back of the female. | 15.26 ± 0.37 | 2.41 ± 0.71 | 2.92 ± 0.51 |
| Mouth-to-mouth posturing | A male and female touch each other with their snouts. | 12.93 ± 1.03 | 2.06 ± 0.65 | 1.60 ± 0.24 |
| Chasing | A male chases a female in the den. | 10.78 ± 2.11 | 1.56 ± 0.37 | 3.04 ± 0.57 |
| Cohabiting | A male and female live in the same den. | 10.76 ± 0.58 | 88.32 ± 3.23 | 181.25 ± 38.67 |
| Inviting | A male swims in and out of the den repeatedly until a female enters. | 7.76 ± 0.68 | 1.77 ± 0.38 | 2.80 ± 0.43 |
| Rolling over | A male turns his body over to show himself to a female. | 1.41 ± 0.15 | 0.40 ± 0.14 | 4.71 ± 0.81 |
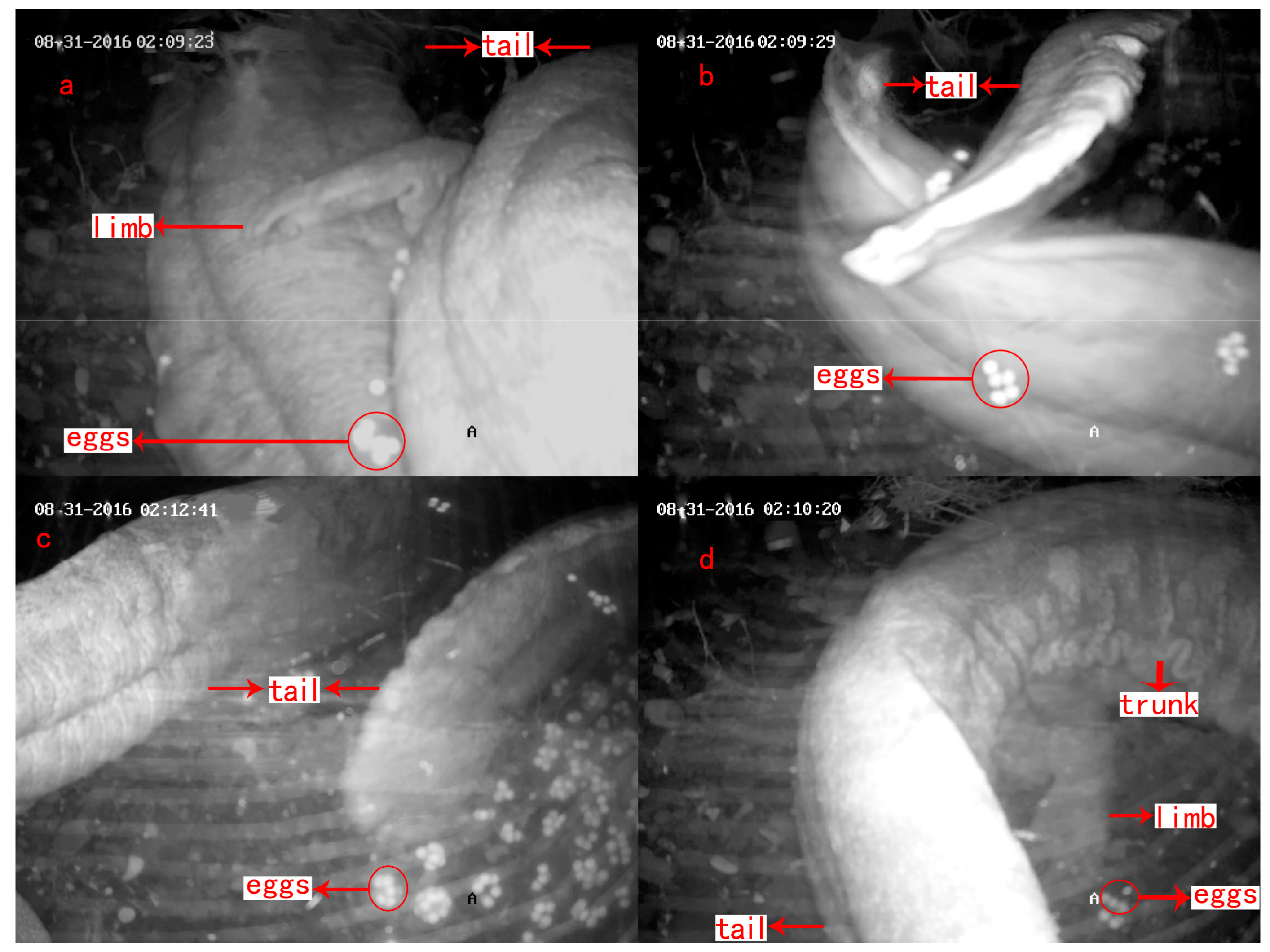
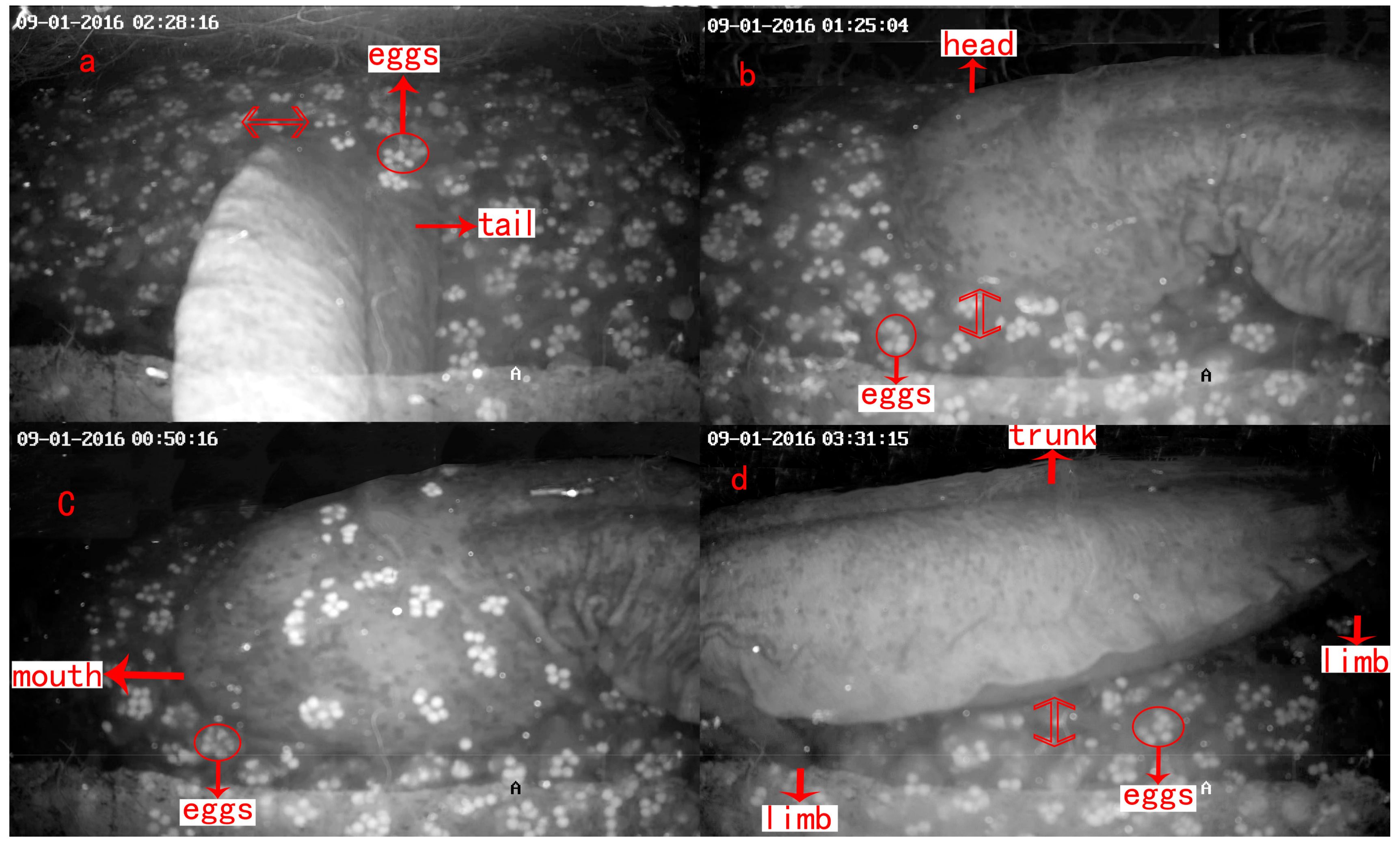
| Parental Care | Description | Duration (min/time) |
|---|---|---|
| Tail fanning | Tail swayed within or beside a pile of eggs like a fan (Figure 6a) | 3.5 ± 0.6 |
| Agitating | Head or body moved within a cluster of eggs (Figure 6b) | 3.1 ± 0.6 |
| Egg eating | The male ate the eggs that were unfertilized, had yolk stuck to shell or had water mold infection. This often followed agitating behavior (Figure 6c) | 2.6 ± 0.8 |
| Shaking | The trunk was placed above a pile of eggs, and shaken up and down or right and left with four limbs grounded on the same position (Figure 6d) | 2.1 ± 0.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Q.; Tong, F.; Song, Y.; Wang, H.; Du, M.; Ji, H. Observation of the Breeding Behavior of the Chinese Giant Salamander (Andrias davidianus) Using a Digital Monitoring System. Animals 2018, 8, 161. https://doi.org/10.3390/ani8100161
Luo Q, Tong F, Song Y, Wang H, Du M, Ji H. Observation of the Breeding Behavior of the Chinese Giant Salamander (Andrias davidianus) Using a Digital Monitoring System. Animals. 2018; 8(10):161. https://doi.org/10.3390/ani8100161
Chicago/Turabian StyleLuo, Qinghua, Fang Tong, Yingjie Song, Han Wang, Maolin Du, and Hongbing Ji. 2018. "Observation of the Breeding Behavior of the Chinese Giant Salamander (Andrias davidianus) Using a Digital Monitoring System" Animals 8, no. 10: 161. https://doi.org/10.3390/ani8100161
APA StyleLuo, Q., Tong, F., Song, Y., Wang, H., Du, M., & Ji, H. (2018). Observation of the Breeding Behavior of the Chinese Giant Salamander (Andrias davidianus) Using a Digital Monitoring System. Animals, 8(10), 161. https://doi.org/10.3390/ani8100161