Nature of Pre-Earthquake Phenomena and their Effects on Living Organisms


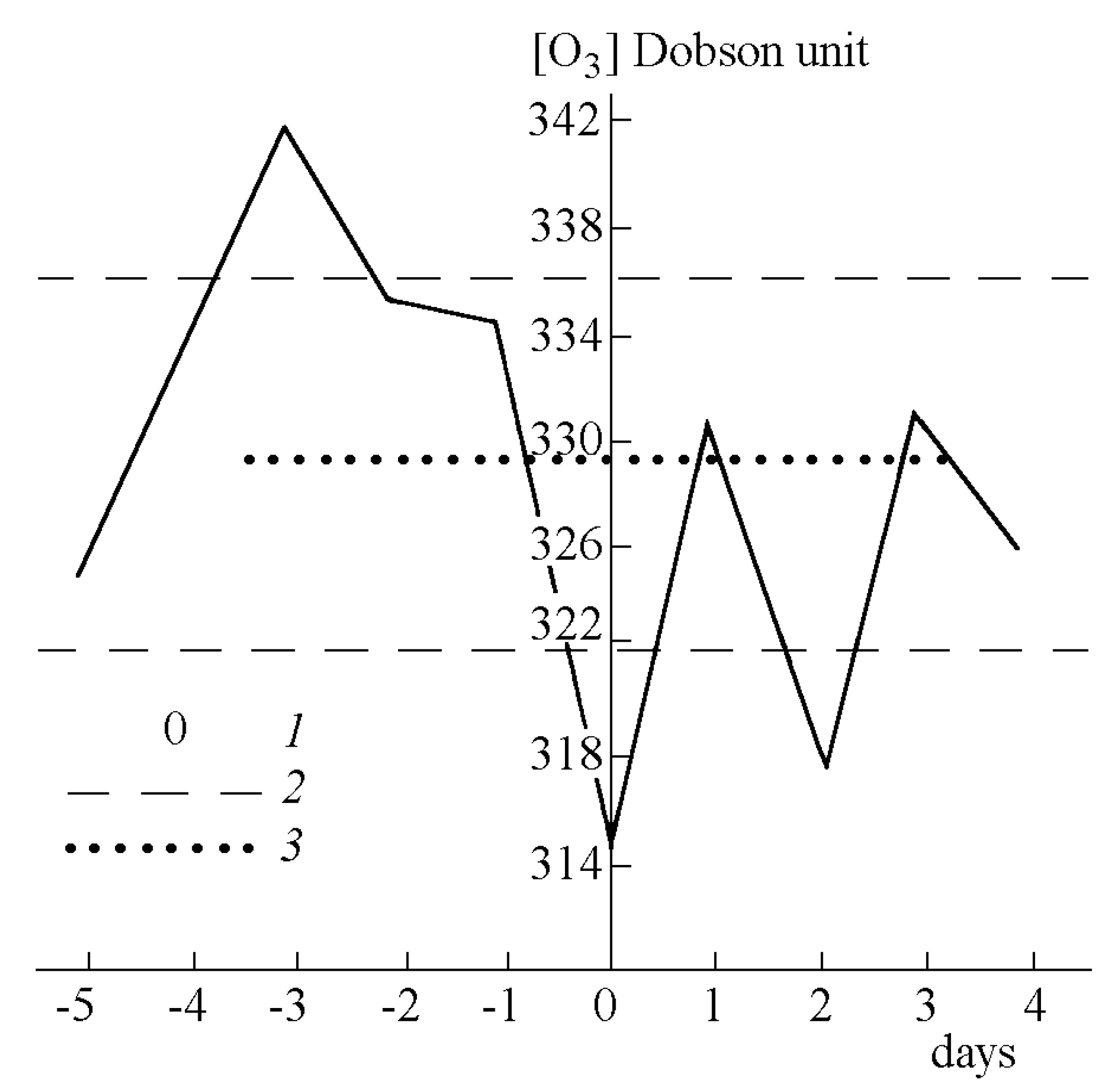{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Setting the Stage
1.2. Shifting Paradigm
2. Pre-Earthquake Processes and the Biosphere
2.1. The Effect of Electromagnetic Waves on Organismal Physiology
2.2. Surface/Subsurface Charges, Air Ionization, and Corona Discharges
2.3. Oxidizing Water to Hydrogen Peroxide at the Rock-Water Interface
2.4. Oxidation Reactions in the Soil
2.4.1. Ozone
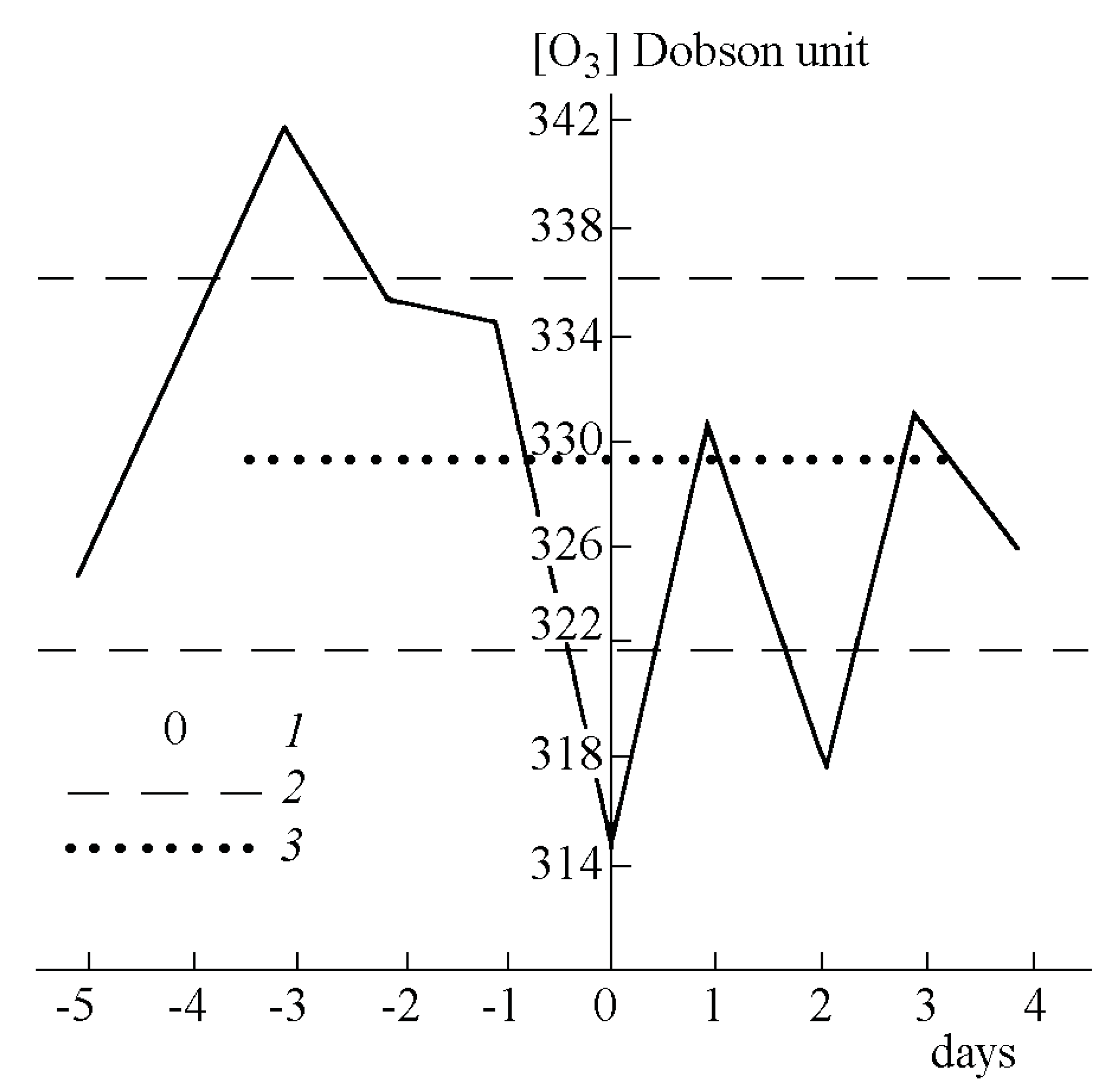
2.4.2. Radon
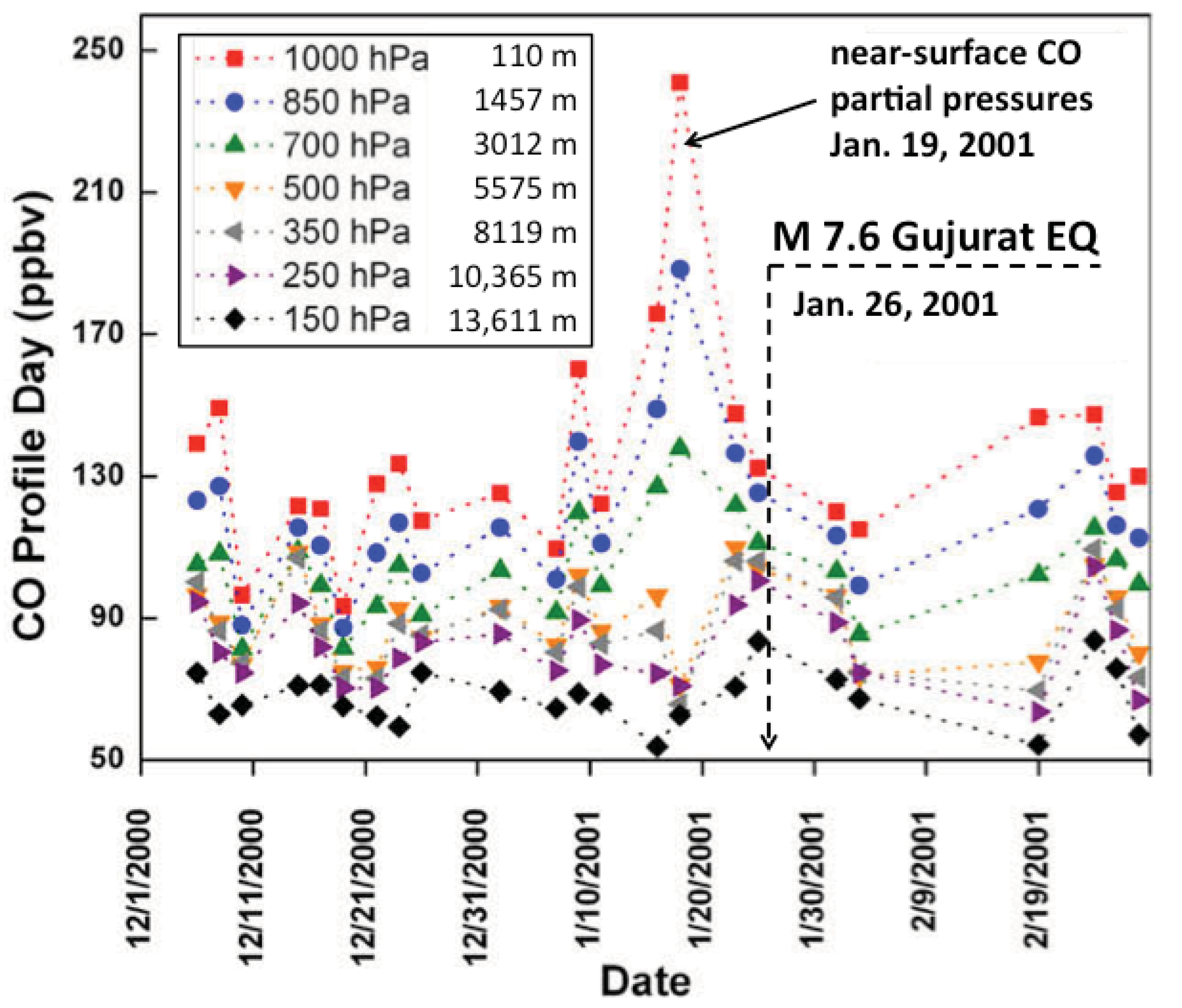
2.4.3. Carbon Monoxide

3. Results and Discussion
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Main, I.G. Statistical physics, seismogenesis, and seismic hazard. Rev. Geophys. 1996, 34, 433–462. [Google Scholar] [CrossRef]
- Hough, S.E. Earthshaking Science: What We Know (and Don’t Know) about Earthquakes; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Rundle, J.B.; Turcotte, D.L.; Shcherbakov, R.; Klein, W.; Sammis, C. Statistical physics approach to understanding the multiscale dynamics of earthquake fault systems. Rev. Geophys. 2003, 41, 1019–1049. [Google Scholar] [CrossRef]
- Keilis-Borok, V. Fundamentals of Earthquake Prediction: Four Paradigms. In Nonlinear Dynamics of the Lithosphere and Earthquake Prediction; Keilis-Borok, V., Soloviev, A.A., Eds.; Springer: Heidelberg, Germany, 2003; pp. 1–36. [Google Scholar]
- Sykes, L.; Shaw, B.E.; Scholz, C.H. Rethinking earthquake prediction. Pure Appl. Geophys. 1999, 155, 207–232. [Google Scholar] [CrossRef]
- Geller, R.J.; Jackson, D.D.; Kagan, Y.Y.; Mulargia, F. Earthquakes cannot be predicted. Science 1997, 275, 1616–1617. [Google Scholar] [CrossRef]
- Scholz, C.H. The Mechanism of Earthquakes and Faulting; Cambrdge University Press: Cambridge, UK, 2002. [Google Scholar]
- Scholz, C.H. Microfractures, aftershocks, and seismicity. Bull. Seismol. Soc. Am. 1968, 58, 1117–1130. [Google Scholar]
- Brace, W.F.; Paulding, B.W., Jr.; Scholz, C. Dilatancy in the fracture of crystalline rocks. J. Geophys. Res. 1966, 71, 3939–3953. [Google Scholar] [CrossRef]
- Cristescu, N. Rock dilatancy in uniaxial tests. Rock Mech. 1982, 15, 133–144. [Google Scholar] [CrossRef]
- Reches, Z.E.; Lockner, D.A. Nucleation and growth of faults in brittle rocks. J. Geophys. Res. 1994, 99, 18159–18173. [Google Scholar] [CrossRef]
- Yamashita, T. Generation of microcracks by dynamic shear rupture and its effects on rupture growth and elastic wave radiation. Geophys. J. Int. 2000, 143, 395–406. [Google Scholar] [CrossRef]
- Dobrovolsky, I.P.; Gershenzon, N.I.; Gokhberg, M.B. Theory of electrokinetic effects occurring at the final stage in the preparation of a tectonic earthquake. Phys. Earth Planet. Int. 1989, 57, 144–156. [Google Scholar] [CrossRef]
- Merzer, M.; Klemperer, S.L. Modeling low-frequency magnetic-field precursors to the Loma Prieta earthquake with a precursory increase in fault-zone conductivity. Pure Appl. Geophys. 1997, 150, 217–248. [Google Scholar] [CrossRef]
- Gershenzon, N.; Bambakidis, G. Modeling of seismo-electromagnetic phenomena. Russian J. Earth Sci. 2001, 3, 247–275. [Google Scholar] [CrossRef]
- Simpson, J.J.; Taflove, A. Electrokinetic effect of the Loma Prieta earthquake calculated by an entire-Earth FDTD solution of Maxwell’s equations. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Hauksson, E. Radon content of groundwater as an earthquake precursor: Evaluation of worldwide data and physical basis. J. Geophys. Res. 1981, 86, 9397–9410. [Google Scholar] [CrossRef]
- Igarashi, G.; Saeki, S.; Takahata, N.; Sumikawa, K.; Tasaka, S.; Sasaki, Y.; Takahashi, M.; Sano, Y. Ground-Water Radon Anomaly Before the Kobe Earthquake in Japan. Science 1995, 269, 60–61. [Google Scholar]
- Virk, H.S.; Walia, V.; Kumar, N. Helium/radon precursory anomalies of Chamoli earthquake, Garhwal Himalaya, India. J. Geodynam. 2001, 31, 201–210. [Google Scholar] [CrossRef]
- King, C.-Y. Episodic radon changes in subsurface soil gas along active faults and possible relation to earthquakes. J. Geophys. Res. 1980, 85, 3065–3078. [Google Scholar] [CrossRef]
- Dobrovolsky, I.P.; Zubkov, S.I.; Miachkin, V.I. Estimation of the size of earthquake preparation zones. Pure Appl. Geophys. 1979, 117, 1025–1044. [Google Scholar] [CrossRef]
- Bowman, D.D.; Ouillon, G.; Sammis, C.G.; Sornette, A.; Sornette, D. An observational test of the critical earthquake concept. J.Geophys. Res. 1998, 103, 24359–24372. [Google Scholar]
- Johnston, M.J.S.; Linde, A.T. Implications of crustal strain during conventional, slow and silent earthquakes. In International Handbook of Earthquake and Engineering Seismology; Academic Press: San Diego, CA, USA, 2002; pp. 589–605. [Google Scholar]
- McGarr, A.; Fletcher, J.B. Maximum slip in earthquake fault zones, apparent stress, and stick-slip friction. Bull. Seism. Soc. Am. 2003, 93, 2355–2362. [Google Scholar] [CrossRef]
- Tzanis, A.; Vallianatos, F.; Gruszow, S. Identification and discrimination of transient electrical earthquake precursors: Fact, fiction and some possibilities. Phys. Earth Planet. Inter. 2000, 121, 223–248. [Google Scholar] [CrossRef]
- Zadro, M.; Braitenberg, C. Measurements and interpretations of tilt-strain gauges in seismically active areas. Earth-Sci. Rev. 1999, 47, 51–187. [Google Scholar]
- Freund, F.T.; Takeuchi, A.; Lau, B.W.S. Electric currents streaming out of stressed igneous rocks—A step towards understanding pre-earthquake low frequency EM emissions. Phys. Chem. Earth 2006, 31, 389–396. [Google Scholar] [CrossRef]
- Freund, F.T. Pre-earthquake signals: Underlying physical processes. J. Asian Earth Sci. 2011, 41, 383–400. [Google Scholar] [CrossRef]
- Bortnik, J.; Bleier, T.E.; Dunson, C.; Freund, F. Estimating the seismotelluric current required for observable electromagnetic ground signals. Ann. Geophys. 2010, 28, 1615–1624. [Google Scholar] [CrossRef]
- Zhadin, M.N. Review of Russian literature on biological action of DC and low-frequency AC magnetic fields. Bioelectromagnetics 2001, 22, 27–45. [Google Scholar] [CrossRef]
- Ritz, T. Quantum effects in biology: Bird navigation. Proc. Chem. 2011, 3, 262–275. [Google Scholar] [CrossRef]
- Ritz, T.; Yoshii, T.; Helfrich-Foerster, C.; Ahmad, M. Cryptochrome: A photoreceptor with the properties of a magnetoreceptor? Commun. Integr. Biol. 2010, 3, 24–27. [Google Scholar] [CrossRef]
- Cherry, N. Schumann Resonances, a plausible biophysical mechanism for the human health effects of Solar/Geomagnetic Activity. Nat. Hazards 2002, 26, 279–331. [Google Scholar] [CrossRef]
- Adair, R.K. Biophysical limits on athermal effects of RF and microwave radiation. Bioelectromagnetics 2003, 24, 39–48. [Google Scholar] [CrossRef]
- International Commission on Non-Ionizing Radiation Protection. Guidelines for limiting exposure to time-varying electric and magnetic fields. Health Phys. 2010, 99, 818–836.
- Wiltschko, W.; Wiltschko, R. Magnetic orientation and magnetoreception in birds and other animals. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2005, 191, 675–693. [Google Scholar] [CrossRef]
- Dyson, P.J. Biology: Electric Cows. Nature 2009, 458. [Google Scholar] [CrossRef]
- Burda, H.; Begall, S.; Červený, J.; Neef, J.; Němec, P. Extremely low-frequency electromagnetic fields disrupt magnetic alignment of ruminants. Proc. Nat. Acad. Sci. USA 2009, 106, 5708–5713. [Google Scholar]
- Nakamoto, R.K.; Baylis Scanlon, J.A.; Al-Shawi, M.K. The Rotary Mechanism of the ATP in Synthase. Arch. Biochem. Biophys. 2008, 476, 43–50. [Google Scholar] [CrossRef]
- Bohren, C.F. How can a particle absorb more than the light incident on it? Am. J. Phys. 1983, 51, 323–327. [Google Scholar] [CrossRef]
- Paul, H.; Fischer, R. Light absorption by a dipole. Sov. Phys. Usp. 1983, 26, 923–926. [Google Scholar] [CrossRef]
- Lloyd, D.; Murray, D.B. The Temporal Architecture of Eukaryotic Growth. FEBS Lett. 2006, 580, 2830–2865. [Google Scholar] [CrossRef]
- Freund, F.T.; Kulahci, I.G.; Cyr, G.; Ling, J.; Winnick, M.; Tregloan-Reed, J.; Freund, M.M. Air ionization at rock surface and pre-earthquake signals. J. Atmos. Sol.-Terr. Phys. 2009, 71, 1824–1834. [Google Scholar] [CrossRef]
- Rycroft, M.J.; Harrison, R.G.; Nicoll, K.A.; Mareev, E.A. An Overview of Earth’s Global Electric Circuit and Atmospheric Conductivity. Space Science Rev. 2008, 137, 83–105. [Google Scholar] [CrossRef]
- Bleier, T.; Dunson, C.; Alvarez, C.; Freund, F.; Dahlgren, R. Correlation of pre-earthquake electromagnetic signals with laboratory and field rock experiments. Nat. Hazards Earth Syst. Sci. 2010, 10, 1965–1975. [Google Scholar] [CrossRef]
- Bryant, N. Ground-based and space-based electromagnetic monitoring for pre-earthquake signals. In The Frontier of Earthquake Prediction Studies; Hayakawa, M., Ed.; Nihon-senmontosho-shuppan: Tokyo, Japan, 2011. [Google Scholar]
- Krueger, A.P.; Reed, E.J. Effect of the air ion environment on influenza in the mouse. Int. J. Biometeor. 1972, 16, 209–232. [Google Scholar] [CrossRef]
- Krueger, A.P.; Reed, E.J. Biological impact of small air ions. Science 1976, 193, 1209–1213. [Google Scholar]
- Ganguly, N.D. Variation in atmospheric ozone concentration following strong earthquakes. Int. J. Remote Sens. 2009, 30, 349–356. [Google Scholar] [CrossRef]
- Balk, M.; Bose, M.; Gözen, E.; Rogoff, D.A.; Rothschild, L.J.; Freund, F.T. Oxidation of water to hydrogen peroxide at the rock-water interface due to stress-activated electric currents in rocks. Earth Planet. Sci. Lett. 2009, 283, 87–92. [Google Scholar] [CrossRef]
- Grant, R.A.; Halliday, T.; Balderer, W.P.; Leuenberger, F.; Newcomer, M.; Cyr, G.; Freund, F.T. Ground Water Chemistry Changes before Major Earthquakes and Possible Effects on Animals. Int. J. Environ. Res. Public Health 2011, 8, 1936–1956. [Google Scholar] [CrossRef]
- Kirschvink, J.L. Earthquake prediction by animals: Evolution and sensory perception. Bull. Seism. Soc. Am. 2000, 90, 312–323. [Google Scholar] [CrossRef]
- LiveScience. Mystery of Mass Squid ‘Suicides’ Possibly Solved. Available online: http://www.livescience.com/25550-mass-squid-suicide.html (accessed on 27 May 2013).
- Lelong, A.; Hégaret, H.; Soudant, P.; Bates, S.S. Pseudo-nitzschia (Bacillariophyceae) Species, Domoic Acid and Amnesic Shellfish Poisoning: Revisiting Previous Paradigms. Phycologia 2012, 51, 168–216. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Robertson, A. Domoic acid and human exposure risks: A review. Toxicon 2010, 56, 218–230. [Google Scholar] [CrossRef]
- Than, K. Dozens of Jumbo Squid Beached after Quake—Coincidence? National Geographic News 2009. [Google Scholar]
- Martens, D.A.; Frankenberger, W.T. Enhanced degradation of polycyclic aromatic hydrocarbons in soil treated with an advanced oxidative process—Fenton’s reagent. J. Soil Contam. 1995, 4, 175–190. [Google Scholar] [CrossRef]
- Salazar, J.M.L.; Pérez, N.M.; Hernández, P.A.; Soriano, T.; Barahona, F.; Olmos, R.; Cartagena, R.; López, D.L.; Lima, R.N.; Melián, G.; Galindo, I.; Padrón, E.; Sumino, H.; Notsu, K. Precursory diffuse carbon dioxide degassing signature related to a 5.1 magnitude earthquake in El Salvador, Central America. Earth Planet. Sci. Lett. 2003, 205, 81–89. [Google Scholar]
- Tyrtyshinikov, A.V. The variations of ozone content in the atmosphere above strong earthquake epicenter. Phys. Solid Earth 1996, 31, 789–794. [Google Scholar]
- Heraud, J.A.; Lira, J.A. Co-seismic luminescence in Lima, 150 km from the epicenter of the Pisco, Peru earthquake of 15 August 2007. Nat. Haz. Earth Syst. Sci. 2011, 11, 1025–1036. [Google Scholar] [CrossRef]
- Health Effects of Ozone in the General Population; EPA: Research Triangle Park, NC, USA, 2013.
- Nagarajaa, K.; Prasad, B.S.N.; Madhava, M.S.; Chandrashekara, M.S.; Paramesh, L.; Sannappa, J.; Pawar, S.D.; Murugavel, P.; Kamra, A.K. Radon and its short-lived progeny: Variations near the ground. Radiat. Meas. 2003, 36, 413–417. [Google Scholar] [CrossRef]
- Tsivion, E.; Zilberg, S.; Gerber, R.B. Predicted stability of the organo-xenon compound HXeCCH above the cryogenic range. Chem. Phys. Lett. 2008, 460, 23–26. [Google Scholar] [CrossRef]
- Freund, F.T. Toward a unified solid state theory for pre-earthquake signals. Acta Geophys. 2010, 58, 719–766. [Google Scholar] [CrossRef]
- Kamsali, N.; Pawar, S.D.; Murugavel, P.; Gopalakrishnan, V. Estimation of Small ion concentration near the Earth’s surface. J. Atmos. Sol.-Terr. Phys. 2011, 73, 2345–2351. [Google Scholar] [CrossRef]
- Singh, R.P.; Kumar, J.S.; Zlotnicki, J.; Kafatos, M. Satellite Detection of Carbon Monoxide Emission Prior to the Gujarat Earthquake of 26 January 2001. Appl. Geochem. 2010, 25, 580–585. [Google Scholar] [CrossRef]
- Stolc, V.; Shmygelska, A.; Griko, Y. Adaptation of organisms by resonance of RNA transcription with the cellular redox cycle. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Lu, D. Impending Earthquake Prediction; Jinangsu Science and Publishing House: Nanjing, China, 1988. (In Chinese) [Google Scholar]
- Varon, J.; Marik, P.E. Carbon Monoxide Poisoning. Int. J. Emerg. Intens. Care Med. 1997, 1. [Google Scholar] [CrossRef] [Green Version]
- Italy – Hundreds of Dead Mice Found in Earthquake Area (in Italian, translated into English). Available online: http://iceagenow.info/2012/06/italy-hundreds-dead-mice-earthquake-area/ (accessed on 27 May 2013). [Green Version]
- Jordan, T.H.; Chen, Y.; Gasparini, P.; Madariaga, R.; Main, I.; Marzocchi, W.; Papadopoulos, G.; Sobolev, G.; Yamaoka, K.; Zschau, J. Operational earthquake forecasting: State of knowledge and guidelines for utilization. Ann. Geophys. 2011, 54, 315–391. [Google Scholar]
- Grant, R.A.; Halliday, T. Predicting the unpredictable: Evidence of pre-seismic anticipatory behaviour in the common toad. J. Zool. 2010, 281, 263–271. [Google Scholar]
- Schall, R.B. An evaluation of the animal-behavior theory for earthquake prediction. Calif. Geol. 1988, 41, 41–45. [Google Scholar]
- Evernden, J.F. Abnormal Animal Behavior Prior to Earthquakes; U.S. Department of Interior Geological Survey: Menlo Park, CA, USA, 1976. [Green Version]
- Tributsch, H. When the Snakes Awake; The MIT Press: Cambridge, MA, USA, 1982. [Google Scholar] [Green Version]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Freund, F.; Stolc, V. Nature of Pre-Earthquake Phenomena and their Effects on Living Organisms. Animals 2013, 3, 513-531. https://doi.org/10.3390/ani3020513
Freund F, Stolc V. Nature of Pre-Earthquake Phenomena and their Effects on Living Organisms. Animals. 2013; 3(2):513-531. https://doi.org/10.3390/ani3020513
Chicago/Turabian StyleFreund, Friedemann, and Viktor Stolc. 2013. "Nature of Pre-Earthquake Phenomena and their Effects on Living Organisms" Animals 3, no. 2: 513-531. https://doi.org/10.3390/ani3020513
APA StyleFreund, F., & Stolc, V. (2013). Nature of Pre-Earthquake Phenomena and their Effects on Living Organisms. Animals, 3(2), 513-531. https://doi.org/10.3390/ani3020513