
Exploring the Use of Helminthophagous Fungi in the Control of Helminthoses in Horses: A Review

 ,
, 
 ,
, 
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Methodology
3. Literature Review
3.1. Gastrointestinal Helminths in Horses
3.2. Anthelmintic Resistance
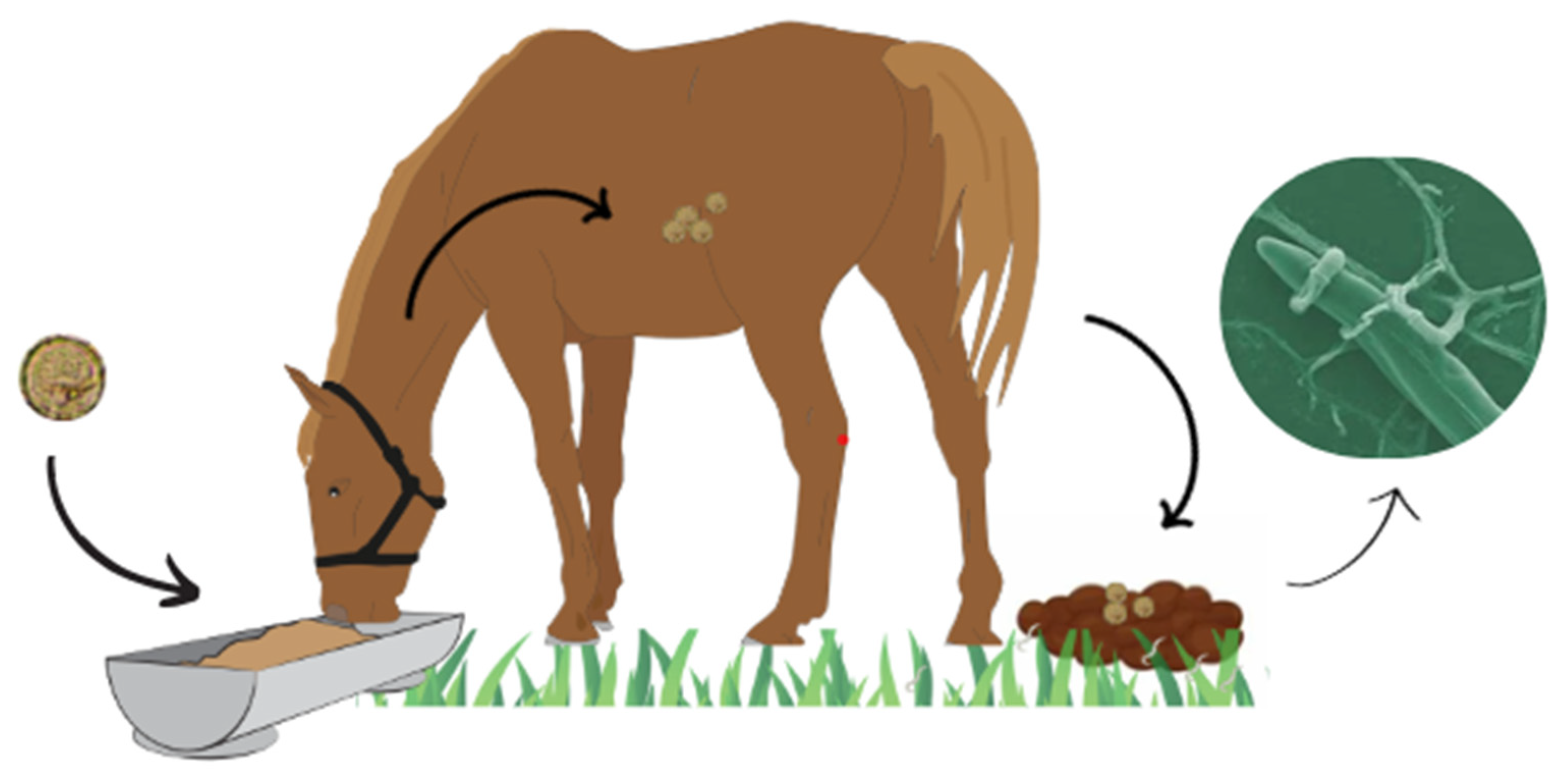
3.3. Helminthophagous Fungi
3.3.1. Virulence Mechanisms
3.3.2. Survival Mechanisms
3.3.3. Overview of Helminthophagous Fungi Species
3.3.4. Methods of Administering Helminthophagous Fungi
3.3.5. Commercial Products—Biotechnological Innovation
3.4. Biological and Sustainable Solution
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lima, R.A.S. The importance and challenges of understanding the equine population in Brazil. Braz. J. Equine Med. 2023, 18, 32–34. [Google Scholar]
- Braga, F.R.; Araújo, J.V.; Silva, A.R.; Araújo, J.M.; Carvalho, R.O.; Tavel, A.O.; Carvalho, G.R.; Campos, A.K. Biological control of horse cyathostomin (Nematoda: Cyathostominae) using the nematophagous fungus Duddingtonia flagrans in tropical southeastern Brazil. Vet. Parasitol. 2009, 163, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Barret, E.J.; Farlam, J.; Proudman, C.J. Field trial of the efficacy of a combination of ivermectin and praziquantel in horses infected with roundworms and tapeworms. Vet. Rec. 2004, 154, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Cupello, F.; Vilarim, D.S.S.; Nogueira, G.R.; Campos, L.D.F.; Costa, M.P.B.; Marques, B.L.O.; Souza, G.O.; Monteiro, J.L.A.B.C.; Salema, M.H.M.; Pacheco, A. Challenge in the management and control of parasites in horses: Literature review. Ciências Agrárias 2024, 28, 59–60. [Google Scholar] [CrossRef]
- Lagaggio, V.R.A.; Jorge, L.L. Achados de formas parasitárias em camas de equinos Santa Maria-RS/Brasil. 2007. Available online: https://www.redevet.com.br/index.php/profissionais/na-rede/ibrajournal/113-geral/390-achados-de-formas-parasitarias-em-camas-de-equinos-1?showall=1 (accessed on 21 January 2025).
- Molento, M.B. Resistência parasitária em helmintos de equídeos e propostas de manejo. Ciência Rural 2005, 35, 1469–1477. [Google Scholar] [CrossRef]
- Araújo, J.V.; Braga, F.R.; Mendoza-de-Gives, P.; Paz-Silva, A.; Vilela, V.L.R. Recent advances in the control of helminths of domestic animals by helminthophagous fungi. Parasitologia 2021, 1, 168–176. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.; Li, H.; Wang, R.; Zhang, K.Q.; Xu, J. Fungi-Nematode Interactions: Diversity, Ecology, and Biocontrol Prospects in Agriculture. J. Fungi 2020, 6, 206. [Google Scholar] [CrossRef]
- Salmo, R.; Viña, C.; Zubiria, I.; Malagón, J.Á.H.; Sanchís, J.M.; Cazapal, C.; Arias, M.S.; Sánchez-Andrade, R.; Paz-Silva, A. Formulating Parasiticidal Fungi in Dried Edible Gelatins to Reduce the Risk of Infection by Trichuris sp. among Continuous Grazing Bison. Pathogens 2024, 13, 82. [Google Scholar] [CrossRef]
- Carvalho, R.O.; Araújo, J.V.; Braga, F.R.; Araujo, J.M.; Silva, A.R.; Tavela, A.O. Predatory activity of nematophagous fungi on infective larvae of Ancylostoma sp.: Evaluation in vitro and after passing through the gastrointestinal tract of dogs. J. Helminthol. 2009, 83, 231–236. [Google Scholar] [CrossRef]
- Ferreira, S.R.; Machado, A.R.T.; Furtado, L.F.; Gomes, J.H.D.S.; de Almeida, R.M.; de Oliveira Mendes, T.; Maciel, V.N.; Barbosa, F.S.; Carvalho, L.M.; Bueno, L.L.; et al. Ketamine can be produced by Pochonia chlamydosporia: An old molecule and a new anthelmintic. Parasites Vectors 2020, 13, 527. [Google Scholar] [CrossRef] [PubMed]
- Lichtenfels, J.R.; Kharchenko, V.A.; Dvojnos, G.M. Illustrated identification keys to strongylid parasites (Strongylidae: Nematoda) of horses, zebras and asses (Equidae). Vet. Parasitol. 2008, 156, 4–161. [Google Scholar] [CrossRef]
- Nielsen, M. Sustainable equine parasite control: Perspectives and research needs. Vet. Parasitol. 2012, 185, 32–44. [Google Scholar] [CrossRef]
- Rehbein, S.; Visser, M.; Winter, R. Koproskopische Untersuchun gen bei Pferden in Deutschland und Österreich. Pferdeheilkunde, 2002, 18:439–449. Pferdeheilkunde 2002, 18, 439–449. [Google Scholar] [CrossRef]
- Madeira de Carvalho, L.; Gomes, L.; Cernea, M.; Cernea, C.; Santos, C.A.; Bernardes, N.; Rosário, M.A.; Soares, M.J.; Fazendeiro, I. Parasitismo gastrointestinal e seu controlo em asininos e híbridos estabulados. Rev. Port. De Ciências Veterinárias 2007, 102, 225–231. [Google Scholar]
- Lester, H.E.; Spanton, J.; Stratford, C.H.; Bartley, D.J.; Morgan, E.R.; Hodgkinson, J.E.; Coumbe, K.; Mair, T.; Swan, B.; Lemon, G.; et al. Anthelmintic efficacy against cyathostomins in horses in Southern England. Vet. Parasitol. 2014, 197, 189–196. [Google Scholar] [CrossRef]
- Kaplan, R.M.; Nielsen, M.K. An evidence-based approach to equine parasite control: It ain’t the 60s anymore. Equine Vet. Educ. 2010, 22, 306–316. [Google Scholar] [CrossRef]
- Luksovsky, J.; Craig, T.M.; Bingham, G.M.; Cir, T.; Forrets, D. Determining treatment to control two multidrug-resistant parasites on a Texas horse farm. J. Equine Vet. Sci. 2013, 33, 115–119. [Google Scholar] [CrossRef]
- Rehbein, S.; Martin, V.; Renate, W. Prevalence, intensity and seasonality of gastrointestinal parasites in abattoir horses in Germany. Parasitol. Res. 2013, 112, 407–413. [Google Scholar] [CrossRef]
- Ibrahim, N. Equine lung worm: A systematic review. Glob. J. Med. Res. G Vet. Sci. Vet. Med. 2017, 17, 19–25. [Google Scholar]
- Mas-Coma, S.; Esteban, J.G.; Bargues, M.D. Fasciola, lymnaeids and human fascioliasis, with a global overview on disease transmission and control. Infect. Dis. Poverty 2009, 2, 41–146. [Google Scholar] [CrossRef]
- Guide to the Treatment and Control of 8 Equine Gastrointestinal Parasite Infections. ESCCAP Guideline08 Second Edition—March. European Scientific Counsel Companion Animal Parasites. 2019. Available online: https://www.esccap.org/guidelines/gl8/ (accessed on 13 January 2025).
- El-Gameel, S.M.; Al-Mokaddem, A.K.; Salaeh, N.M.K.; Attia, M.M. Morphomolecular characterization of Strongylus vulgaris isolated from donkeys with special reference to histopathological study on the affected organs. J. Parasit. Dis. 2022, 46, 795–803. [Google Scholar] [CrossRef]
- Teixeira, W.F.P.; Felippelli, G.; Cruz, B.C.; Maciel, W.G.; Fávero, F.C.; Gomes, L.V.C.; Buzzulini, C.; Prando, L.; Bichuette, M.A.; Lopes, W.D.Z.; et al. Endoparasites of horses from the Formiga city, located in center-west region of the state of Minas Gerais, Brazil. Rev. Bras. Parasitol. Veterinária 2014, 23, 534–538. [Google Scholar] [CrossRef]
- Saes, I.L.; Gonçalves, J.A.; Do Carmo, T.A.; Mena, M.O.; De Almeida Cipriano, I.; De Favare, G.M.; Bello, H.J.S.; Guelpa, G.J.; de Soutello, R.V.G. Sazonalidade e dinâmica de helmintos gastrointestinais em equinos em pastejo. Acta Vet.-Beogr. 2022, 72, 131–144. [Google Scholar] [CrossRef]
- Soutello RVG de Romão, D.S.; Carmo, T.A.; do Favare, G.M.; de Pagnozzi, F.C.; Cipriano, I.d.A. Seasonal dynamics of gastrointestinal helminths in naturally infected horses in Brazil. Vet. Parasitol. 2025, 324, 110358. [Google Scholar] [CrossRef]
- Canever, R.J.; Braga, P.R.; Boeckh, A.; Grycajuck, M.; Bier, D.; Molento, M.B. Lack of Cyathostomin sp. reduction after anthelmintic treatment in horses in Brazil. Vet. Parasitol. 2013, 194, 9–39. [Google Scholar] [CrossRef]
- Allison, K.; Taylor, N.M.; Wilsmore, A.J.; Garforth, C. Equine anthelmintics: Survey of the patterns of use, beliefs and attitudes among horse owners in the UK. Vet. Rec. 2011, 168, 483–487. [Google Scholar] [CrossRef]
- Peregrine, A.S.; Molento, M.B.; Kaplan, R.M.; Nielsen, M.K. Anthelmintic resistance in important parasites of horses: Does it really matter? Vet. Parasitol. 2014, 201, 1–8. [Google Scholar] [CrossRef]
- Saes, I.L.; Vera, J.H.S.; Fachiolli, D.F.; Yamada, P.H.; Dellaqua, J.V.T.; Saes, R.L.; Amaranto, A.F.; Soutello, R.V.G. Tempo necessário para diferentes anti-helmínticos atingirem os níveis de eficácia esperados em equinos infectados por estrongilídeos. Parasitol. Veterinária 2016, 229, 90–92. [Google Scholar] [CrossRef]
- Vera, J.H.S.; Fachioli, D.F.; Ramires, L.M.; Saes, I.L.; Yamada, P.H.; Gonçalves, J.A.; Oliveira, K.; Amarante, A.F.T.; Soutello, R.V.G. Eficacy of ivermectin, moxidectin and febendazole in equine in Brazil. Vet. Parasitol. 2020, 20, 100374. [Google Scholar] [CrossRef]
- Nielsen, N.K.; Banahan, M.; Kaplan, R.M. Importation of macrocyclic lactone resistant cyathostomins on a US thoroughbred farm. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 99–104. [Google Scholar] [CrossRef]
- de Favare, G.M.; Cipriano, I.A.; do Carmo, T.A.; Mena, M.O.; Guelpa, G.J.; do Amarante, A.F.T.; de Soutello, R.V.G. Anthelmintic resistance of horse strongyle nematodes to ivermectin in São Paulo state, Brazil. Vet. Parasitol. Reg. Stud. Rep. 2023, 41, 100864. [Google Scholar] [CrossRef]
- Nielsen, M.K.; Reinemeyer, C.R.; Donecker, J.M.; Leathwick, D.M. Anthelmintic resistance in equine parasites—Current evidence and knowledge gaps. Vet. Parasitol. 2014, 204, 55–63. [Google Scholar] [CrossRef]
- Oliveira, J.H.; Carvalho, N.; Rinaldi, L.; Oliveira, G.; Amarante, A.F.T. Diagnóstico da resistência anti-helmíntica em bovinos no Brasil: Uma comparação de diferentes metodologias. Vet. Parasitol. 2014, 206, 216–226. [Google Scholar] [CrossRef]
- Oliveira, F.; Oliveira, C.B.; Oliveira, C.Z.; Oliveira, P.; Osmari, V.; Fernandes, F.; Sangioni, L.A.; Vogel, F.S.F. Avaliação molecular e de campo da resistência anti-helmíntica de populações de nematóides de bovinos e ovinos naturalmente infectados em pastagens mistas no Rio Grande do Sul, Brasil. Acta Parasitol. 2019, 65, 118–127. [Google Scholar] [CrossRef]
- Taylor, M.A.; Coop, R.L.; Wall, R.L. Veterinary Parasitology, 4th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Araújo, J.V.; Guimarães, M.P.; Campos, A.K.; Sá, N.C.; Sarti, P.; Assis, R.C. Control of bovine gastrointestinal nematode parasites using pellets of the nematode-trapping fungus Monacrosporium thaumasium. Ciência Rural 2004, 34, 457–463. [Google Scholar] [CrossRef]
- Mota, M.A.; Campos, A.K.; Araújo, J.V. Biological control of animal parasitic helminths by nematophagous fungi. Braz. J. Parasitol. Vete 2003, 13, 165–170. [Google Scholar] [CrossRef]
- Mendoza-de-Gives, P.; López-Arellano, M.E.; Aguilar-Marcelino, L.; Olazarán-Jenkins, S.; Reyes-Guerrero, D.; Oliveira, G.; Veja-Murillo, V.E. The nematophagous fungus Duddingtonia flagrans reduces the gastrointestinal parasitic nematode larvae population in faeces of orally treated calves maintained under tropical conditions—Dose/response assessment. Vet. Parasitol. 2018, 263, 66–72. [Google Scholar] [CrossRef]
- Braga, F.R.; Oliveira, C.M.; Silva, E.M.; Araújo, J.V. Efficiency of the fungal formulation Bioverm (Duddingtonia flagrans) in the in vivo and in vitro control of Haemonchus contortus and Strongyloides papillosus in sheep. Biotecnologia 2020, 10, 62. [Google Scholar] [CrossRef]
- Fausto, G.C.; Fausto, M.C.; Vieira, Í.S.; Freitas, S.G.; Carvalho, L.M.; Oliveira, I.C.; Silva, E.N.; Campos, A.K.; Araújo, J.V. Formulation of the nematophagous fungus Duddingtonia flagrans in the control of equine gastrointestinal parasitic nematodes. Vet. Parasitol. 2021, 296, 109458–109464. [Google Scholar] [CrossRef]
- Carmo, T.A.; Mena, M.O.; Cipriano, I.A.; Favare, G.M.; Guelpa, G.J.; Pinto, S.C.; Amarante, A.F.T.; Araújo, J.V.; Soutello, R.V.G. Biological control of gastrointestinal nematodes in horses fed with grass in association with nematophagous fungi Duddingtonia flagrans and Pochonia chlamydosporia. Biol. Control 2023, 182, 105219. [Google Scholar] [CrossRef]
- Mendoza de Gives, P.; Braga, F.R.; Araújo, J.V. Nematophagous fungi, an extraordinary tool for controlling ruminant parasitic nematodes and other biotechnological applications. Biocontrol Sci. Technol. 2022, 32, 777–793. [Google Scholar] [CrossRef]
- Li, S.; Wang, D.; Gong, J.; Zhang, Y. Individual and combined application of nematophagous fungi as biological control agents against gastrointestinal nematodes in domestic animals. Pathogens 2022, 11, 172. [Google Scholar] [CrossRef] [PubMed]
- Soares, F.E.D.F.; de Queiroz, J.H.; de Araújo, J.V.; Queiroz, P.V.; Gouveia, A.D.S.; Hiura, E.; Braga, F.R. Ação nematicida de quitinases produzidas pelo fungo Monacrosporium thaumasium em condições laboratoriais. Biocontrol Sci. Technol. 2015, 3, 337–344. [Google Scholar] [CrossRef]
- Liu, X.; Xiang, M.; Che, Y. A estratégia viva dos fungos nematófagos. Micociência 2009, 50, 20–25. [Google Scholar] [CrossRef]
- Zhong, W.; Chen, Y.; Gong, S.; Qiao, J.; Meng, Q.; Zhang, X.; Wang, X.; Huang, Y.; Tian, L.; Niu, Y. Enzymological properties and nematode-degrading activity of recombinant chitinase AO-379 of Arthrobotrys oligospora. Kafkas Üni. Vet. Fakültesi Derg. 2019, 25, 435–444. [Google Scholar] [CrossRef]
- Zhang, K.Q.; Hyde, K.Q. Nematode-Trapping Fungi; Fungal Diversity Series 23; Mushroom Research Foundation: Chiang Mai, Thailand, 2014. [Google Scholar] [CrossRef]
- Rahman, M.u.; Chen, P.; Zhang, X.; Fan, B. Predatory strategies of nematophagous fungi as biocontrol agents. Agronomy 2023, 13, 2685. [Google Scholar] [CrossRef]
- Nordbring-Hertz, B.; Jansson, H.B.; Tunlid, A. Nematophagous fungi. In Encyclopedia of Life Sciences; Macmillan Publishers Ltd.: Weinheim, Germany, 2006; pp. 1–11. [Google Scholar] [CrossRef]
- Nordbring-Hertz, B. Ecology and recognition in the nematode-nematophagous fungus system. In Advances in Microbial Ecology; Marshall, K.C., Ed.; Springer: Berlin/Heidelberg, Germany, 1998; Volume 10, pp. 1–16. [Google Scholar] [CrossRef]
- Gronvold, J.; Henriksen, S.A.; Larsen, M.; Nansen, P.; Wolstrup, J. Aspects of biological control with special reference to arthropods, protozoans and helminths of domesticated animals. Vet. Parasitol. 1996, 64, 47–64. [Google Scholar] [CrossRef]
- Herrera, T.; Ulloa, M. El Reino de los Hongos: Micología Básica y Aplicada; Universidad Nacional Autónoma de México, Fondo de Cultura Económica: Mexico City, Mexico, 1990. [Google Scholar]
- Braga, F.R.; Araújo, J.V. Nematophagous fungi for biological control of gastrointestinal nematodes in domestic animals. Appl. Microbiol. Biotechnol. 2014, 98, 71–82. [Google Scholar] [CrossRef]
- Cruz, D.G.; Araújo, F.B.; Molento, M.B. Kinetics of capture and infection of infective larvae of trichostrongylides and free-living nematodes Panagrellus sp. by Duddingtonia flagrans. Parasitol. Res. 2011, 109, 1085–1091. [Google Scholar] [CrossRef]
- Dallemole-Giaretta, R.; Freitas, L.G.; Cavallin, I.C.; Marmentini, G.A.; Faria, C.M.R.; Resende, J.T.V. Evaluation of a Pochonia chlamydosporia based product, for the control of Meloidogyne javanica in culture and in carrot fields. Nematropica 2013, 43, 131–137. [Google Scholar]
- Araujo, J.M.; Araújo, J.V.; Braga, F.R.; Carvalho, R.O.; Silva, A.R.; Campos, A.K. Interaction and ovicidal activity of the nematophagous fungus Pochonia chlamydosporia on Taenia saginata eggs. Exp. Parasitol. 2009, 121, 338–341. [Google Scholar] [CrossRef]
- Vilela, V.L.R.; Feitosa, T.F.; Braga, F.R.; Araújo, J.V.; Souto, D.V.O.; Santos, H.E.S.; Athayde, A.C.R. Biological control of goat gastrointestinal helminthiasis by Duddingtonia flagrans in a semi-arid region of the Northeastern Brazil. Vet. Parasitol. 2012, 188, 127–133. [Google Scholar] [CrossRef]
- Vilela, V.L.R.; Feitosa, T.F.; Braga, F.R.; Araújo, J.V.; Lucena, S.C.; Dantas, E.S.; Athayde, A.C.R.; Silva, A.R. Effect of Monacrosprorium thaumasium in the control of the goat gastrointestinal helminthiasis in the semi-arid of Brazil. Parasitol. Res. 2013, 112, 871–877. [Google Scholar] [CrossRef]
- Saxena, G.; Mittal, N. Trap formation by conidia of nematode-trapping Monacrosporium spp. Mycol. Res 1995, 7, 839–840. [Google Scholar] [CrossRef]
- Larsen, M. Biological control of helminths. Int. J. Parasitol. 1999, 29, 139–146. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, N.; Gong, P.T.; Li, J. Effect of temperature, pH, physical and chemical factors on germination rate of the chlamydospores of the nematophagous fungus Duddingtonia flagrans. FEMS Microbiol. Lett. 2019, 366, fnz212. [Google Scholar] [CrossRef]
- Braga, F.R.; Araújo, J.V.; Ribeiro, V.L.; Soares, F.E.F.; Araújo, J.M.; Silva, G.R. Viability of the nematophagous fungus Pochonia chlamydosporia after passage through the gastrointestinal tract of horses. Vet. Parasitol. 2010, 168, 264–268. [Google Scholar] [CrossRef]
- Vieira, Í.S.; Oliveira, I.C.; Campos, A.K.; Araújo, J.V. In vitro biological control of bovine parasitic nematodes by Arthrobotrys cladodes, Duddingtonia flagrans, and Pochonia chlamydosporia under different temperature conditions. J. Helminthol. 2020, 94, e194. [Google Scholar] [CrossRef]
- Santos, C.P.; Padilha, T.; Rodrigues, M.L.A. Predatory activity of Arthrobotrys oligospora and Duddingtonia flagrans on pre-parasitic larval stages of cyathostominae under different constant temperatures. Ciência Rural 2001, 31, 839–842. [Google Scholar] [CrossRef]
- Hernández, J.Á.; Cazapal-Monteiro, C.F.; Sanchís, J.; Sánchez-Andrade, R.; Paz-Silva, A.; Arias, M.S. Potential Usefulness of Filamentous Fungi to Prevent Zoonotic Soil-Transmitted Helminths. Vector Borne Zoonotic Dis. 2018, 18, 690–696. [Google Scholar] [CrossRef]
- Braga, F.R.; Araújo, J.V.; Soares, F.E.F.; Araújo, J.M.; Geniêr, H.L.A.; Silva, A.R.; Carvalho, R.O.; Queiroz, J.H.; Ferreira, S.R. Optimizing protease production from an isolate of the nematophagous fungus Duddingtonia flagrans using response surface methodology and its larvicidal activity on horse cyathostomin. J. Helminthol. 2011, 85, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, C.M.; Costa Silva, L.P.; de Freitas Soares, F.E.; Souza, R.L.O.; Tobias, F.L.; de Araújo, J.V.; Rodrigues Veloso, F.B.; Laviola, F.P.; Endringer, D.C.; de Gives, P.M.; et al. Effect of silver nanoparticles (AgNPs) from Duddingtonia flagrans on cyathostomins larvae (subfamily: Cyathostominae). J. Invertebr. Pathol. 2020, 175, 107907. [Google Scholar] [CrossRef]
- Ward, E.; Kerry, B.R.; Manzanilla-López, R.H.; Mutua, G.; Devonshire, J.; Kimenju, J.; Hirsch, P.R. The Pochonia chlamydosporia Serine Protease Gene vcp1 is subject to regulation by carbon, nitrogen and pH: Implications for nematode biocontrol. PLOS ONE 2012, 7, e35657. [Google Scholar] [CrossRef]
- Hao, L.; Zhao, F.; Guo, Y.; Ma, Y.; Li, Z.; Wang, W.; Luo, H.; Wang, R. Antagonistic activity of Pochonia chlamydosporia against three helminth eggs and characterization of its serine protease. Vet. Parasitol. 2024, 334, 110374. [Google Scholar] [CrossRef]
- Esteves, I.; Peteira, B.; Atkins, S.D.; Magan, N.; Kerry, B. Production of extracellular enzymes by different isolates of Pochonia chlamydosporia. Mycol. Res. 2009, 8, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Mi, Q.; Yang, J.; Ye, F.; Gan, Z.; Wu, C.; Niu, X.; Zou, C.; Zhang, K.Q. Clonagem e superexpressão do gene pcchi44 da quitinase de Pochonia chlamydosporia, um potencial fator de virulência na infecção contra nematoides. Process Biochem. 2010, 45, 810–814. [Google Scholar] [CrossRef]
- Soares, F.E.F.; Sufiate, B.L.; Queiroz, J.H. Fungos nematófagos: Muito além dos grupos endoparasita, predador e ovicida. Agric. Nat. Resour. 2018, 52, 1–8. [Google Scholar] [CrossRef]
- Silva, A.R.; Araújo, J.V.; Braga, F.R.; Benjamim, L.A.; Souza, D.L.; Carvalho, R.O. Análise comparativa da destruição das formas infectantes de Trichuris trichiura e Haemonchus contortus por fungos nematófagos Pochonia chlamydosporia, Duddingtonia flagrans e Monacrosporium thaumasium por microscopia eletrônica de varredura. Microbiol. Veterinária 2011, 147, 214–219. [Google Scholar] [CrossRef]
- Cazapal-Monteiro, C.F.; Hernández, J.A.; Arroyo, F.L. Analysing the effect of soil saprophytic fungi on Baylisascaris procyonis eggs. Res. Parasitol. 2015, 114, 2443–2450. [Google Scholar] [CrossRef]
- Arias, M.S.; Oliveira, C.F.; Souza, J.; Miguélez, S.; Oliveira, I.; Oliveira, F.L.; Suárez, J.L.; Paz-Silva, A.; Sánchez-Andrade, R.; De Gives, P.M. Mixed production of filamentous fungal spores for preventing soil-transmitted helminth zoonoses: A preliminary analysis. BioMed Res. Int. 2013, 2013, 567876. [Google Scholar] [CrossRef]
- Arroyo, F.; Oliveira, J.A.; Oliveira, C.F.; Pedreira, J.; Oliveira, J.; Romasanta, Á.; Sánchez-Andrade, R.; Paz-Silva, A.; Arias, M.S. Effect of the Filamentous Fungus Mucor circinelloides On The Development of Eggs of the Rumen Fluke Calicophoron daubneyi (Paramphistomidae). J. Parasitol. 2017, 103, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Cortiñas, F.J.; Cazapal-Monteiro, C.F.; Hernández, J.A.; Arroyo, F.L.; Miguélez, S.; Suárez, J.; López de Arellano, M.E.; Sánchez-Andrade, R.; Mendoza de Gives, P.; Paz-Silva, A.; et al. Potential use of Mucor circinelloides for the biological control of certain helminths affecting livestock reared in a care farm. Biocontrol Sci. Technol. 2015, 25, 1443–1452. [Google Scholar] [CrossRef]
- Voinot, M.; Bonilla, R.; Sousa, S.; Sanchís, J.; Canhão-Dias, M.; Romero Delgado, J.; Lozano, J.; Sánchez-Andrade, R.; Sol Arias, M.; Madeira de Carvalho, L. Control of Strongyles in First-Season Grazing Ewe Lambs by Integrating Deworming and Thrice-Weekly Administration of Parasiticidal Fungal Spores. Pathogens 2021, 10, 1338. [Google Scholar] [CrossRef] [PubMed]
- Paz-Silva, A.; Francisco, I.; Valero-Coss, R.O. Ability of the fungus Duddingtonia flagrans to adapt to the cyathostomin egg-output by spreading chlamydospores. Vet. Parasitol. 2011, 179, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Duddington, C.L. Fungi that attack microscopic animals. Bot. Rev. 1955, 21, 377–479. [Google Scholar] [CrossRef]
- Buzatti, A.; Santos, C.P.; Fernandes, M.A.M.; Yoshitani, U.Y.; Sprenger, L.K.; Molento, M.B. Duddingtonia flagrans no controle de nematoides gastrintestinais de equinos em fases de vida livre. Arq. Bras. Med. Veterinária E Zootec. 2015, 69, 364–370. [Google Scholar] [CrossRef]
- Bampidis, V.; Azimonti, G.; Bastos, M.L.; Christensen, H.; Dusemund, B.; Kos Durjava, M.; Kouba, M.; López-Alonso, M.; López Puente, S.; Marcon, F. Scientific opinion on the safety and efficacy of BioWorma (Duddingtonia flagrans NCIMB 30336) as a feed additive for all grazing animals. EFSA J. 2020, 18, 6208. [Google Scholar] [CrossRef]
- Oliveira, L.S.S.C.B.; Dias, F.G.S.; Melo, A.L.T.; Carvalho, L.M.; Silva, E.N.; Araújo, J.V. Bioverm in the control of nematodes in beef cattle raised in the Central-West region of Brazil. Pathogens 2021, 10, 548. [Google Scholar] [CrossRef]
- Rodrigues, J.A.; Roque, F.L.; Alvares, F.B.V.; Silva, A.L.P.; Lima, E.F.; Silva Filho, G.M.; Feitosa, T.F.; Araújo, J.V.; Braga, F.R.; Vilela, V.L.R. Efficacy of a commercial fungal formulation containing Duddingtonia flagrans (Bioverm) for controlling bovine gastrointestinal nematodes. Rev. Bras. De Parasitol. Veterinária 2021, 30, e026620. [Google Scholar] [CrossRef]
- Hernández, J.A.; Arroyo, F.L.; Suárez, J.; Cazapal-Monteiro, C.F.; Romasanta, A.; López-Arellano, M.E.; Pedreira, J.; Madeira de Carvalho, L.M.; Sánchez-Andrade, R.; Arias, M.S.; et al. Feeding horses with industrially manufactured pellets with fungal spores to promote nematode integrated control. Vet. Parasitol. 2016, 229, 37–44. [Google Scholar] [CrossRef]
- Arroyo, F.L.; Arias, M.S.; Cazapal-Monteiro, C.F.; Hernández, J.Á.; Suárez, J.; Miguélez, S.; Romasanta, A.; Sánchez-Andrade, R.; Paz-Silva, A. The capability of the fungus Mucor circinelloides to maintain parasiticidal activity after the industrial feed pelleting enhances the possibilities of biological control of livestock parasites. Biol. Control 2016, 92, 38–44. [Google Scholar] [CrossRef]
- Hernández, J.Á.; Sánchez-Andrade, R.; Cazapal-Monteiro, C.F.; Arroyo, F.L.; Sanchís, J.M.; Paz-Silva, A.; Arias, M.S. A combined effort to avoid strongyle infection in horses in an oceanic climate region: Rotational grazing and parasiticidal fungi. Parasites Vectors 2018, 11, 240. [Google Scholar] [CrossRef] [PubMed]
- Costa-Silva, L.P.; Pinto-Oliveira, J.; Keijok, W.J.; Silva, A.R.; Aguiar, A.R.; Guimarães, M.C.C.; Ferraz, C.M.; Araújo, J.V.; Tobias, F.L.; Braga, F.R. Extracellular biosynthesis of silver nanoparticles using the cell-free filtrate of nematophagous fungus Duddingtonia flagrans. Int. J. Nanomed. 2017, 12, 6373–6381. [Google Scholar] [CrossRef]
- Barbosa, A.C.M.S.; Silva, L.P.C.; Ferraz, C.M.; Tobias, F.L.; Araújo, J.V.; Loureiro, B.; Braga, G.M.A.M.; Veloso, F.B.R.; Soares, F.E.F.; Fronza, M. Nematicidal activity of silver nanoparticles from the fungus Duddingtonia flagrans. Int. J. Nanomed. 2019, 14, 2341–2348. [Google Scholar] [CrossRef] [PubMed]
- Braga, F.R.; Soares, F.E.F.; Giuberti, T.Z.; Lopes, A.D.C.G.; Lacerda, T.; Ayupe, T.D.H.; Queiroz, P.V.; Gouveia, A.d.S.; Pinheiro, L.; Araújo, A.L.; et al. Nematocidal activity of extracellular enzymes produced by the nematophagous fungus Duddingtonia flagrans on cyathostomin infective larvae. Vet. Parasitol. 2015, 212, 214–218. [Google Scholar] [CrossRef]
- Viña, C.; Salmo, R.; Pena, M.V.; Palomero, A.M.; Hernández, J.Á.; Cazapal-Monteiro, C.; Arias, M.S.; Sánchez-Andrade, R.; Paz-Silva, A. A New Comestible Formulation of Parasiticide Fungi to Reduce the Risk of Soil-Transmitted Helminth Infections in a Canine Shelter. Pathogens 2022, 11, 1391. [Google Scholar] [CrossRef]
- Reinemeyer, R. Anthelmintic resistance in non-strongylid parasites of horses. Vet. Parasitol. 2012, 185, 9–15. [Google Scholar] [CrossRef]
- Heredero, A.M.C.; Segato, G.; Menotta, S.; Faggionato, E.; Vismarra, A.; Genchi, M.; Bertini, S.A. new method for ivermectin detection and quantification through HPLC in organic matter (feed, soil, and water). J. Anal. Methods Chem. 2023, 2023, 6924263. [Google Scholar] [CrossRef]
- Silva, E.P.; Souza, J.R.; Caldas, E. Residues of veterinary medicines in milk and eggs. New Quim. 2014, 37, 111–122. [Google Scholar] [CrossRef]
- Cezar, A.S.; Catto, J.B.; Bianchin, I. Controle alternativo de nematódeos gastrintestinais dos ruminantes: Atualidade e perspectivas. Ciência Rural 2008, 38, 2083–2091. [Google Scholar] [CrossRef]
- Almeida, A.C.D.F.; Chagas, J.D.R.; Ávila, L.M.; Marques, T.L.P.; Moraes, R.F.F.; Gomes, L.P.M.; Roier, E.C.R.; Baêta, B.A. Diagnosis and chemical control of helminthiases in cattle: Literature review. Res. Soc. Dev. 2020, 9, 4089119908. [Google Scholar] [CrossRef]
- Madeira de Carvalho, L.M.; Gillespie, A.T.; Serra, P.M. Eficácia do fungo nematófago Duddingtonia flagrans no controlo biológico da estrongilidose equina no Ribatejo. Rev. Port. Ciências Veterinárias 2007, 102, 233–247. [Google Scholar]
- Araújo, J.M.; Braga, F.R.; Longo Ribeiro Vilela, V.; Soares, F.E.D.F.; Ferraz, C.M.; Bindaco, A.L.S.; Aguiar, D.F.; Negrão-Corrêa, D.A.; Fernandes Rodrigues, V.; de Araújo, J.V. Evaluation of the acute oral toxicity of the fungus Duddingtonia flagrans at the gut level. Biocontrol Sci. Technol. 2022, 32, 1275–1284. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
do Carmo, T.A.; Fonseca, J.d.S.; Braga, F.R.; Paz-Silva, A.; de Soutello, R.V.G.; de Araújo, J.V. Exploring the Use of Helminthophagous Fungi in the Control of Helminthoses in Horses: A Review. Animals 2025, 15, 864. https://doi.org/10.3390/ani15060864
do Carmo TA, Fonseca JdS, Braga FR, Paz-Silva A, de Soutello RVG, de Araújo JV. Exploring the Use of Helminthophagous Fungi in the Control of Helminthoses in Horses: A Review. Animals. 2025; 15(6):864. https://doi.org/10.3390/ani15060864
Chicago/Turabian Styledo Carmo, Tábata Alves, Júlia dos Santos Fonseca, Fabio Ribeiro Braga, Adolfo Paz-Silva, Ricardo Velludo Gomes de Soutello, and Jackson Victor de Araújo. 2025. "Exploring the Use of Helminthophagous Fungi in the Control of Helminthoses in Horses: A Review" Animals 15, no. 6: 864. https://doi.org/10.3390/ani15060864
APA Styledo Carmo, T. A., Fonseca, J. d. S., Braga, F. R., Paz-Silva, A., de Soutello, R. V. G., & de Araújo, J. V. (2025). Exploring the Use of Helminthophagous Fungi in the Control of Helminthoses in Horses: A Review. Animals, 15(6), 864. https://doi.org/10.3390/ani15060864