
Impact of Ants on the Order Composition of Canopy Arthropod Communities in Temperate and Tropical Forests


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.1.1. Temperate Forests in Central Europe
2.1.2. Tropical Primary Lowland Rain Forests in Southeast Asia
2.1.3. Comparison of Tropical Primary and Disturbed Rain Forests in Southeast Asia
2.2. Canopy Fogging and Order Sorting
2.3. Statistics
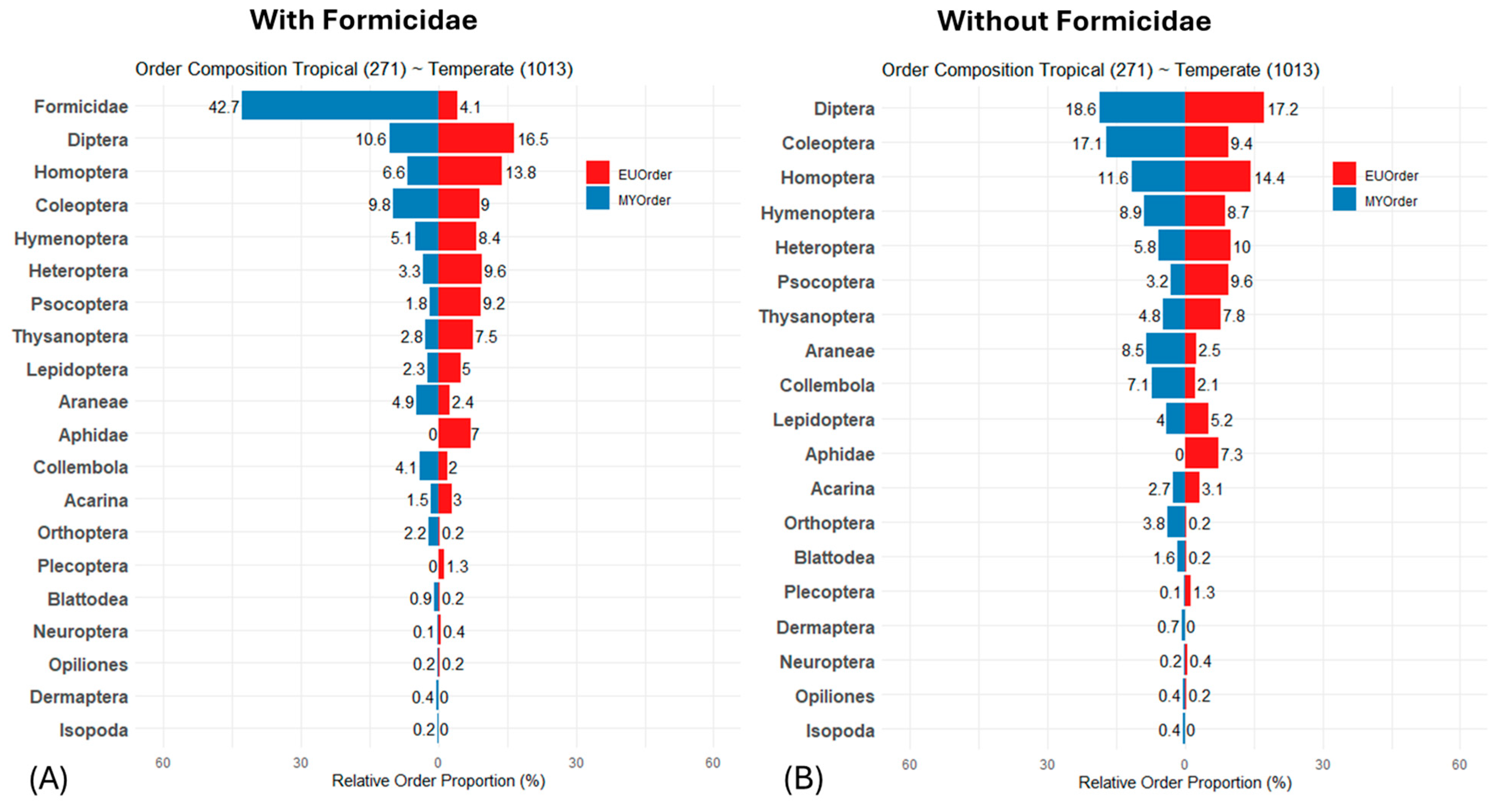
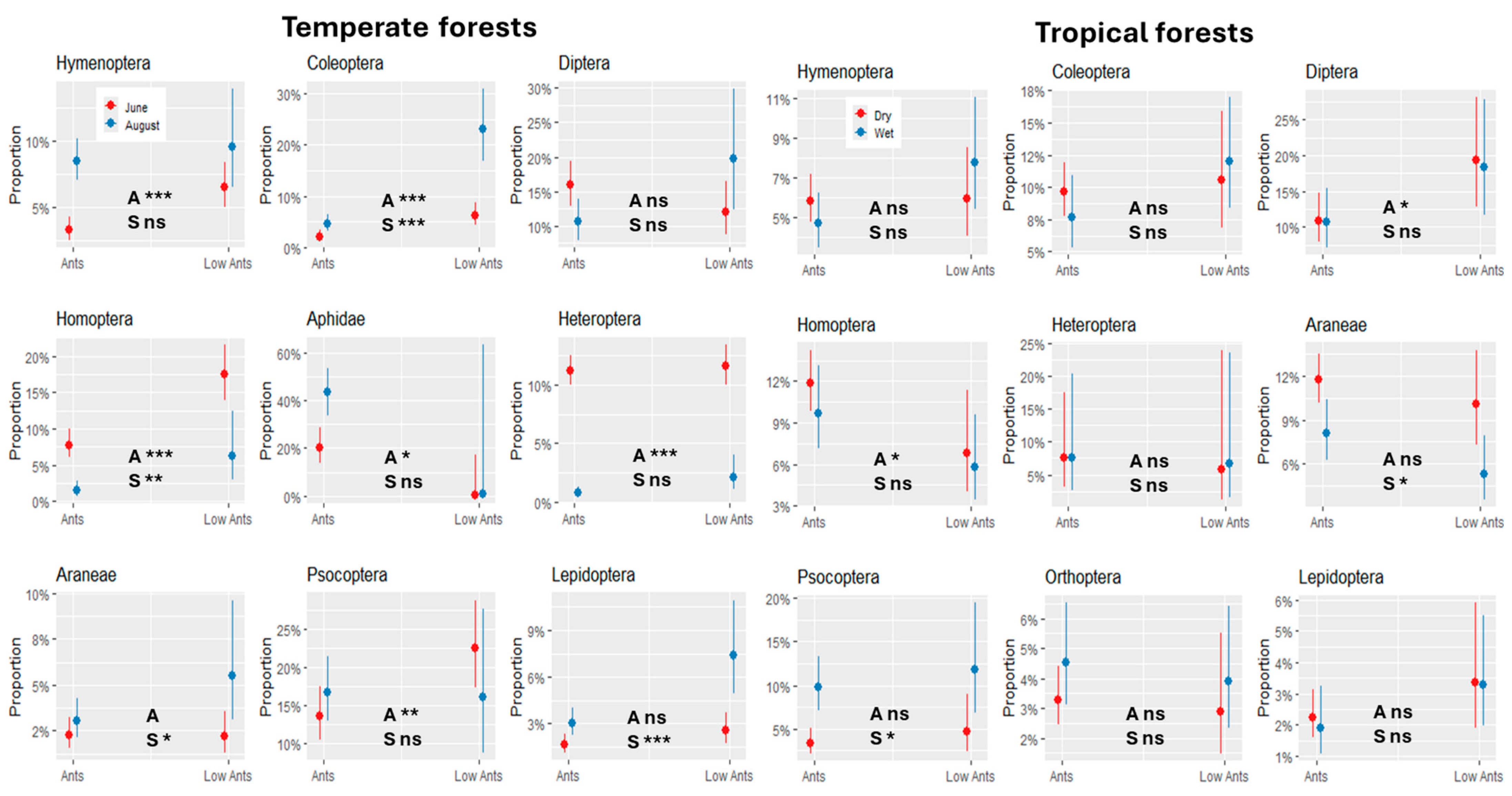
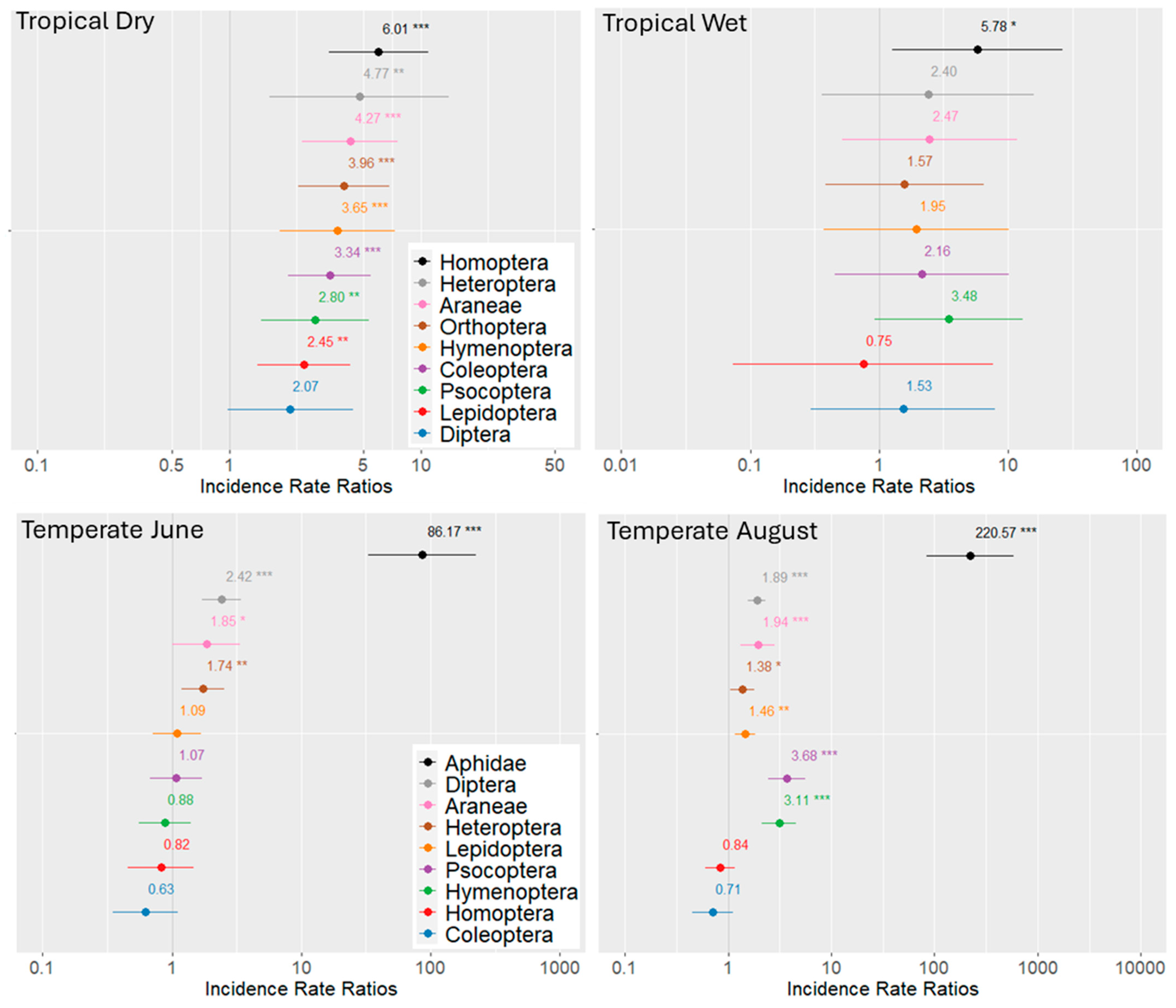
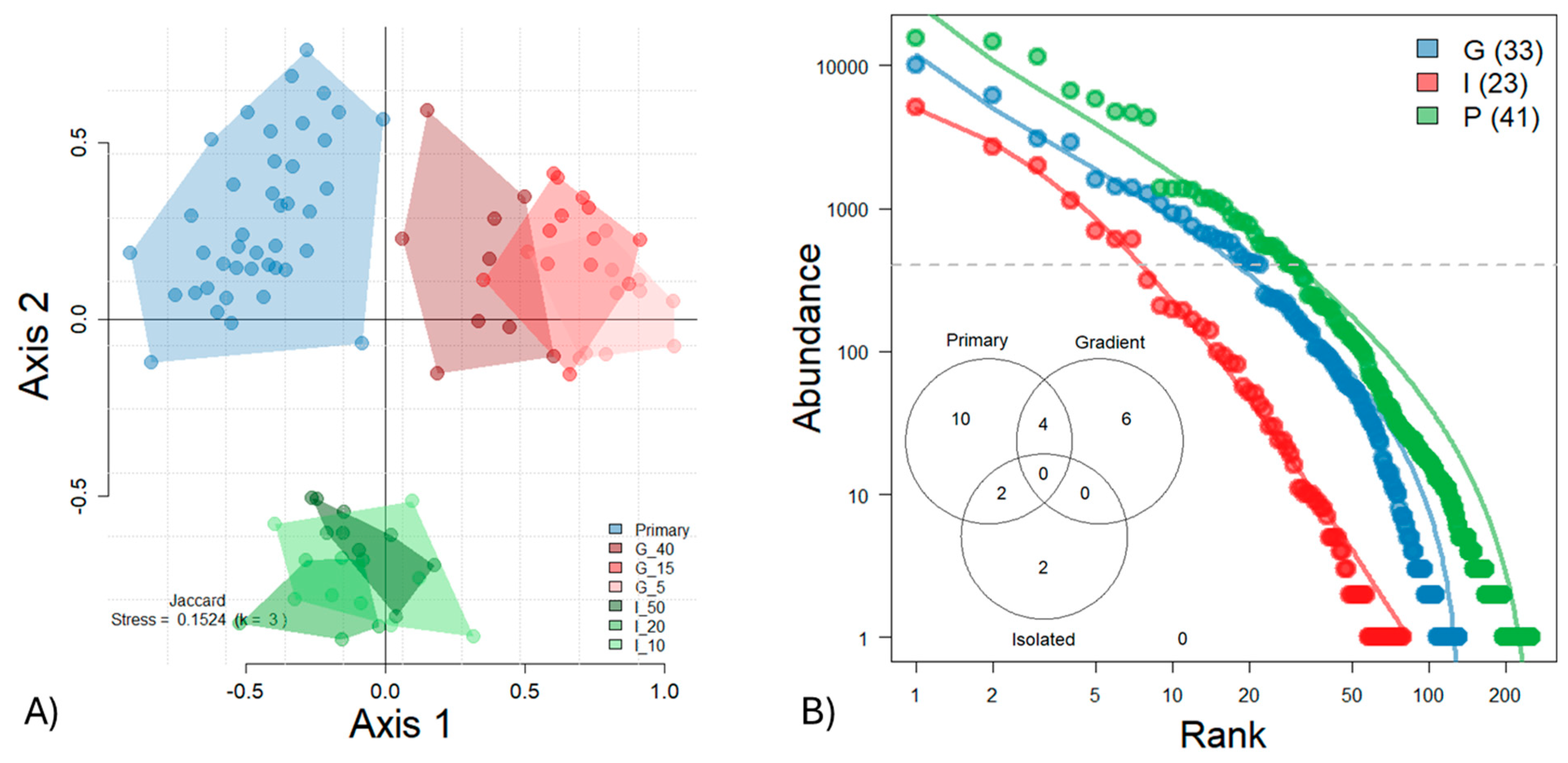
3. Results
4. Discussion
Disturbance Alters Canopy Ant Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parker, J.; Kronauer, D.J.C. How ants shape biodiversity. Curr. Biol. 2021, 31, R1208–R1214. [Google Scholar] [CrossRef] [PubMed]
- Kass, J.M.; Guénard, B.; Dudley, K.L.; Jenkins, C.N.; Azuma, F.; Fisher, B.L.; Parr, C.L.; Gibb, H.; Longino, J.T.; Ward, P.S.; et al. The global distribution of known and undiscovered ant biodiversity. Sci. Adv. 2022, 8, eabp9908. [Google Scholar] [CrossRef] [PubMed]
- Floren, A.; Wetzel, W.; Staab, M. The contribution of canopy species to overall ant diversity (Hymenoptera: Formicidae) in temperate and tropical ecosystems. Myrmecol. News 2014, 19, 65–74. [Google Scholar]
- Schultheiss, P.; Nooten, S.S.; Wang, R.; Wong, M.K.L.; Brassard, F.; Guénard, B. The abundance, biomass, and distribution of ants on Earth. Proc. Natl. Acad. Sci. USA 2022, 119, e2201550119. [Google Scholar] [CrossRef]
- Floren, A.; Biun, A.; Linsenmair, K.E. Arboreal ants as key predators in tropical lowland rainforest trees. Oecologia 2002, 131, 137–144. [Google Scholar] [CrossRef]
- Ali, J.; Abbas, A.; Abbas, S.; Ji, Y.; Khan, K.A.; Ghramh, H.A.; Mahamood, M.; Chen, R. Honeydew: A keystone in insect–plant interactions, current insights and future perspectives. J. Appl. Entomol. 2024, 148, 727–733. [Google Scholar] [CrossRef]
- Nelson, A.S.; Mooney, K.A. The Evolution and Ecology of Interactions Between Ants and Honeydew-Producing Hemipteran Insects. Annu. Rev. Ecol. Evol. Syst. 2022, 53, 379–402. [Google Scholar] [CrossRef]
- Floren, A.; Horchler, P.; Sprick, P.; Müller, T. The ecological role of Fraxinus for species diversity in floodplain forests. Oecologia 2025, 207, 42. [Google Scholar] [CrossRef]
- Basset, Y.; Cizek, L.; Cuénoud, P.; Didham, R.K.; Novotny, V.; Ødegaard, F.; Roslin, T.; Tishechkin, A.K.; Schmidl, J.; Winchester, N.N.; et al. Arthropod Distribution in a Tropical Rainforest: Tackling a Four Dimensional Puzzle. PLoS ONE 2015, 10, e0144110. [Google Scholar] [CrossRef]
- Ismaeel, A.; Tai, A.P.K.; Santos, E.G.; Maraia, H.; Aalto, I.; Altman, J.; Doležal, J.; Lembrechts, J.J.; Camargo, J.L.; Aalto, J.; et al. Patterns of tropical forest understory temperatures. Nat. Commun. 2024, 15, 549. [Google Scholar] [CrossRef]
- Klimes, P. Diversity and Specificity of Ant-Plant Interactions in Canopy Communities: Insights from Primary and Secondary Tropical Forests in New Guinea. In Ant Plant Interactions: Impacts of Humans on Terrestrial Ecosystems; Oliveira, P.S., Koptur, S., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 26–51. [Google Scholar]
- Floren, A.; Linsenmair, K.E. The influence of anthropogenic disturbances on the structure of arboreal arthropod communities. Plant Ecol. 2001, 153, 153–167. [Google Scholar] [CrossRef]
- Wilker, I.; Queiroz, A.C.M.; Ribas, C.R.; Morini, M.S.C.; Lasmar, C.J.; Schmidt, F.A.; Feitosa, R.M.; Nogueira, A.; Baccaro, F.B.; Ulysséa, M.A.; et al. A systematic review of the land use change effects on ant diversity in Neotropics. Biol. Conserv. 2024, 299, 110778. [Google Scholar] [CrossRef]
- Sivault, E.; Kollross, J.; Jorge, L.R.; Finnie, S.; Diez-Méndez, D.; Fernandez-Garzon, S.; Maraia, H.; Lenc, J.; Libra, M.; Murakami, M.; et al. Insectivorous birds and bats outperform ants in the top-down regulation of arthropods across strata of a Japanese temperate forest. J. Anim. Ecol. 2024, 93, 1622–1638. [Google Scholar] [CrossRef]
- Gösswald, K. Die Waldameise: Biologie, Ökologie und forstliche Nutzung; Aula Verlag: Wiebelsheim, Germany, 2012; Volume Teil1, Teil2. [Google Scholar]
- Horstmann, J. Der Einfluß der kleinen Waldameisen (Formica polyctena FOERSTER) auf die Zusammensetzung und Diversität arborikoler Arthropodengemeinschaften in Eichen; Bayerische Julius-Maximilians-Universität Würzburg: Würzburg, Germany, 1999. [Google Scholar]
- Kitayama, K. An altitudinal transect study of the vegetation on Mount Kinabalu, Borneo. Vegetatio 1992, 102, 149–171. [Google Scholar] [CrossRef]
- Barthlott, W.; Mutke, J.; Rafiqpoor, M.D.; Kier, G.; Kreft, H. Global centers of vascular plant diversity. Nova Acta Leopold. 2005, 92, 61–83. [Google Scholar]
- Beaman, J.H.; Beaman, R.S. Diversity and distribution patterns in the flora of Mount Kinabalu. In The Plant Diversity of Malesia; Baas, P., Kalkman, K., Geesink, R., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 147–160. [Google Scholar]
- Horstmann, K.; Floren, A.; Linsenmair, K.E. Ichneumonidae (Hymenoptera) from the canopy of tropical forests in Sabah, Malaysia: A comparison between primary and secondary forests. Ecotropica 2005, 11, 41–52. [Google Scholar]
- Marini, F.; Linke, J.; Binder, H. ideal: An R/Bioconductor package for Interactive Differential Expression Analysis. BMC Bioinform. 2020, 21, 565. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Oksanen, J.; Simpson, G.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology. R package version 2.6-4. The Comprehensive R Archive Network, 2022. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 15 May 2025).
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: New York, NY, USA, 2017. [Google Scholar]
- Lüdecke, D. sjPlot: Data Visualization for Statistics in Social Science, R package version 2.8.10; GitHub: San Francisco, CA, USA, 2021.
- Martinez, A.P. pairwiseAdonis: Pairwise Multilevel Comparison Using Adonis, R package version 0.4.1; GitHub: San Francisco, CA, USA, 2017.
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Nelsen, M.P.; Moreau, C.S.; Kevin Boyce, C.; Ree, R.H. Macroecological diversification of ants is linked to angiosperm evolution. Evol. Lett. 2023, 7, 79–87. [Google Scholar] [CrossRef]
- Reyes-González, R.; Hugo, T.-H.V.; Alejandro, F.-P.; Matthias, R.; Julián, B.-V.; and Corona-López, A.M. Exploring the proportion of rarity in tropical insects: Evaluating hypotheses and variables. Écoscience 2024, 31, 229–238. [Google Scholar] [CrossRef]
- Floren, A.; von Rintelen, T.; Hebert, P.D.N.; de Araujo, B.C.; Schmidt, S.; Balke, M.; Narakusumo, R.P.; Peggie, D.; Ubaidillah, R.; von Rintelen, K.; et al. Integrative ecological and molecular analysis indicate high diversity and strict elevational separation of canopy beetles in tropical mountain forests. Sci. Rep. 2020, 10, 16677. [Google Scholar] [CrossRef] [PubMed]
- Fernando, N.; Schultz, N.; Palmer, G.; Barton, P. Disturbance impacts on ant interactions in different climatic regions. Insectes Sociaux 2025. [Google Scholar] [CrossRef]
- Roberts, N.; Fyfe, R.M.; Woodbridge, J.; Gaillard, M.J.; Davis, B.A.S.; Kaplan, J.O.; Marquer, L.; Mazier, F.; Nielsen, A.B.; Sugita, S.; et al. Europe’s lost forests: A pollen-based synthesis for the last 11,000 years. Sci. Rep. 2018, 8, 716. [Google Scholar] [CrossRef]
- Seifert, B. The ants of Central European tree canopies (Hymenoptera: Formicidae)—An underestimated population? In Canopy Arthropod Research in Europe; Floren, A., Schmidl, J., Eds.; Bioform Entomology: Nuremberg, Germany, 2008; pp. 157–173. [Google Scholar]
- Kirchner, M.; Sorenson, C.; Blaimer, B.B.; Youngsteadt, E. Reaching new heights: Arboreal ant diversity in a North American temperate forest ecosystem. Insect Conserv. Divers. 2025, 18, 95–106. [Google Scholar] [CrossRef]
- Böhm, S.M.; Wells, K.; Kalko, E.K.V. Top-Down Control of Herbivory by Birds and Bats in the Canopy of Temperate Broad-Leaved Oaks (Quercus robur). PLoS ONE 2011, 6, e17857. [Google Scholar] [CrossRef]
- Mestre, L.; Piñol, J.; Barrientos, J.A.; Espadaler, X. Differential ant exclusion from canopies shows contrasting top-down effects on community structure. Oecologia 2016, 180, 193–203. [Google Scholar] [CrossRef]
- Styrsky, J.D.; Eubanks, M.D. Ecological consequences of interactions between ants and honeydew-producing insects. Proc. R. Soc. B Biol. Sci. 2007, 274, 151–164. [Google Scholar] [CrossRef]
- Davidson, D.W.; Cook, S.C.; Snelling, R.R.; Chua, T.H. Explaining the abundance of ants in lowland tropical rainforest canopies. Science 2003, 300, 969–972. [Google Scholar] [CrossRef]
- Adams, R.M.M.; Wells, R.L.; Yanoviak, S.P.; Frost, C.J.; Fox, E.G.P. Interspecific Eavesdropping on Ant Chemical Communication. Front. Ecol. Evol. 2020, 8, 24. [Google Scholar] [CrossRef]
- Schwab, D.; Wurz, A.; Grass, I.; Rakotomalala, A.A.N.A.; Osen, K.; Soazafy, M.R.; Martin, D.A.; Tscharntke, T. Decreasing predation rates and shifting predator compositions along a land-use gradient in Madagascar’s vanilla landscapes. J. Appl. Ecol. 2021, 58, 360–371. [Google Scholar] [CrossRef]
- Klimes, P.; Idigel, C.; Rimandai, M.; Fayle, T.M.; Janda, M.; Weiblen, G.D.; Novotny, V. Why are there more arboreal ant species in primary than in secondary tropical forests? J. Anim. Ecol. 2012, 81, 1103–1112. [Google Scholar] [CrossRef] [PubMed]
- Boyle, M.J.W.; Bonebrake, T.C.; Dias da Silva, K.; Dongmo, M.A.K.; Machado França, F.; Gregory, N.; Kitching, R.L.; Ledger, M.J.; Lewis, O.T.; Sharp, A.C.; et al. Causes and consequences of insect decline in tropical forests. Nat. Rev. Biodivers. 2025, 1, 315–331. [Google Scholar] [CrossRef]
- Trigos-Peral, G.; Juhász, O.; Kiss, P.J.; Módra, G.; Tenyér, A.; Maák, I. Wood ants as biological control of the forest pest beetles Ips spp. Sci. Rep. 2021, 11, 17931. [Google Scholar] [CrossRef] [PubMed]
- Floren, A.; Linsenmair, K.E. The Importance of Primary Tropical Rain Forest For Species Diversity: An Investigation Using Arboreal Ants as an example. Ecosystems 2005, 8, 559–567. [Google Scholar] [CrossRef]
- Hoenle, P.O.; Plowman, N.S.; Matos-Maraví, P.; de Bello, F.; Bishop, T.R.; Libra, M.; Idigel, C.; Rimandai, M.; Klimes, P. Forest disturbance increases functional diversity but decreases phylogenetic diversity of an arboreal tropical ant community. J. Anim. Ecol. 2024, 93, 501–516. [Google Scholar] [CrossRef]
- Grevé, M.E.; Hager, J.; Weisser, W.W.; Schall, P.; Gossner, M.M.; Feldhaar, H. Effect of forest management on temperate ant communities. Ecosphere 2018, 9, e02303. [Google Scholar] [CrossRef]
- Dolek, M.; Freese-Hager, A.; Bussler, H.; Floren, A.; Liegl Alois; Schmidl, J. Ants on oaks: Effects of forest structure on species composition. J. Insect Conserv. 2009, 13, 367–375. [Google Scholar] [CrossRef]
- Ulyshen, M.; Urban-Mead, K.R.; Dorey, J.B.; Rivers, J.W. Forests are critically important to global pollinator diversity and enhance pollination in adjacent crops. Biol. Rev. 2023, 98, 1118–1141. [Google Scholar] [CrossRef]
- Pillay, R.; Watson, J.E.M.; Hansen, A.J.; Burns, P.; Virnig, A.L.S.; Supples, C.; Armenteras, D.; González-del-Pliego, P.; Aragon-Osejo, J.; Jantz, P.A.; et al. Global rarity of high-integrity tropical rainforests for threatened and declining terrestrial vertebrates. Proc. Natl. Acad. Sci. USA 2024, 121, e2413325121. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formicidae | 25% | 46.30% | 30.10% | 57.80% | Formicidae | 1.10% | 37.10% | 0.40% | 66.50% | ||
| 2429 | 26,937 | 3915 | 20,080 | 53,361 | 256 | 23,800 | 28 | 63,833 | 87,917 | ||
| Aphidae | 0% | 0% | 0% | 0% | Aphidae | 0.40% | 12.60% | 0.70% | 14.50% | ||
| 0 | 0 | 0 | 0 | 0 | 88 | 8107 | 45 | 13,896 | 22,136 | ||
| Diptera | 17.50% | 7.40% | 18.40% | 5.20% | Diptera | 12% | 10% | 19.70% | 3.50% | ||
| 1705 | 4298 | 2391 | 1817 | 10,211 | 2673 | 6431 | 1285 | 3403 | 13,792 | ||
| Coleoptera | 7.90% | 5.20% | 8.40% | 3.20% | Coleoptera | 6.20% | 1.40% | 23% | 1.60% | ||
| 767 | 3016 | 1093 | 1122 | 5998 | 1379 | 887 | 1504 | 1494 | 5264 | ||
| Homoptera | 5.10% | 6.40% | 4% | 4.10% | Homoptera | 17.20% | 4.90% | 6.20% | 0.50% | ||
| 498 | 3705 | 524 | 1420 | 6147 | 3845 | 3140 | 402 | 470 | 7857 | ||
| Heteroptera | 7.60% | 7.10% | 11.40% | 4.80% | Heteroptera | 11.50% | 7% | 2.10% | 0.30% | ||
| 734 | 4152 | 1488 | 1657 | 8031 | 2565 | 4519 | 134 | 259 | 7477 | ||
| Hymenoptera | 4.20% | 2.90% | 4.70% | 1.90% | Hymenoptera | 6.40% | 2.10% | 9.60% | 2.80% | ||
| 404 | 1694 | 612 | 654 | 3364 | 1434 | 1323 | 624 | 2720 | 6101 | ||
| Psocoptera | 3.10% | 1.60% | 6.20% | 3.80% | Psocoptera | 22.30% | 8.60% | 16% | 5.60% | ||
| 298 | 902 | 810 | 1311 | 3321 | 4980 | 5515 | 1047 | 5389 | 16,931 | ||
| Araneae | 7.60% | 6.30% | 3.70% | 3.40% | Araneae | 2.20% | 1.40% | 5.50% | 1% | ||
| 736 | 3668 | 483 | 1185 | 6072 | 482 | 905 | 359 | 975 | 2721 | ||
| Lepidoptera | 2.50% | 1.20% | 2.30% | 0.80% | Lepidoptera | 2.50% | 1% | 7.40% | 1% | ||
| 245 | 703 | 300 | 276 | 1524 | 561 | 664 | 482 | 984 | 2691 | ||
| Orthoptera | 2.20% | 1.80% | 2.70% | 1.90% | Orthoptera | 0.39% | 0.16% | 0.44% | 0.03% | ||
| 211 | 1028 | 354 | 665 | 2258 | 87 | 101 | 29 | 24 | 241 | ||
| Others | 17.40% | 13.90% | 7.90% | 13% | Others | 17.81% | 13.75% | 9.11% | 2.65% | ||
| 1694 | 8065 | 1029 | 4529 | 15,317 | 3978 | 8834 | 595 | 2543 | 15,950 | ||
| Sum | 9721 | 58,168 | 12,999 | 34,716 | 115,604 | Sum | 22,328 | 64,226 | 6534 | 95,990 | 189,078 |
| (A) | Dry | Wet | Sum | (B) | June | August | Sum | ||||
| LowAnts (7) | Ants (9) | LowAnts (11) | Ants (5) | LowAnts (7) | Ants (7) | LowAnts (5) | Ants (7) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Floren, A.; Müller, T. Impact of Ants on the Order Composition of Canopy Arthropod Communities in Temperate and Tropical Forests. Animals 2025, 15, 1914. https://doi.org/10.3390/ani15131914
Floren A, Müller T. Impact of Ants on the Order Composition of Canopy Arthropod Communities in Temperate and Tropical Forests. Animals. 2025; 15(13):1914. https://doi.org/10.3390/ani15131914
Chicago/Turabian StyleFloren, Andreas, and Tobias Müller. 2025. "Impact of Ants on the Order Composition of Canopy Arthropod Communities in Temperate and Tropical Forests" Animals 15, no. 13: 1914. https://doi.org/10.3390/ani15131914
APA StyleFloren, A., & Müller, T. (2025). Impact of Ants on the Order Composition of Canopy Arthropod Communities in Temperate and Tropical Forests. Animals, 15(13), 1914. https://doi.org/10.3390/ani15131914