

Effects of Cyclic Adenosine Monophosphate Nanoliposomes on Growth Performance, Gut Development and Microbiota of Broilers

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experiment Design
2.3. Sample Collection
2.4. Detection Indicators and Methods
2.4.1. Growth Performance
2.4.2. Gastrointestinal Index
2.4.3. Histological Morphology of Jejunum
2.4.4. Expression of Duodenal Hormone mRNA
2.4.5. Microbiota of Jejunum and Cecum
2.4.6. Statistical Analysis of Data
3. Results
3.1. Effects on Growth Performance
3.2. Effects on Indices of Major Digestive Tract
3.3. Effects on Jejunal Intestinal Tissue Morphology
3.4. Effects on Duodenal Intestinal Hormone mRNA Expression
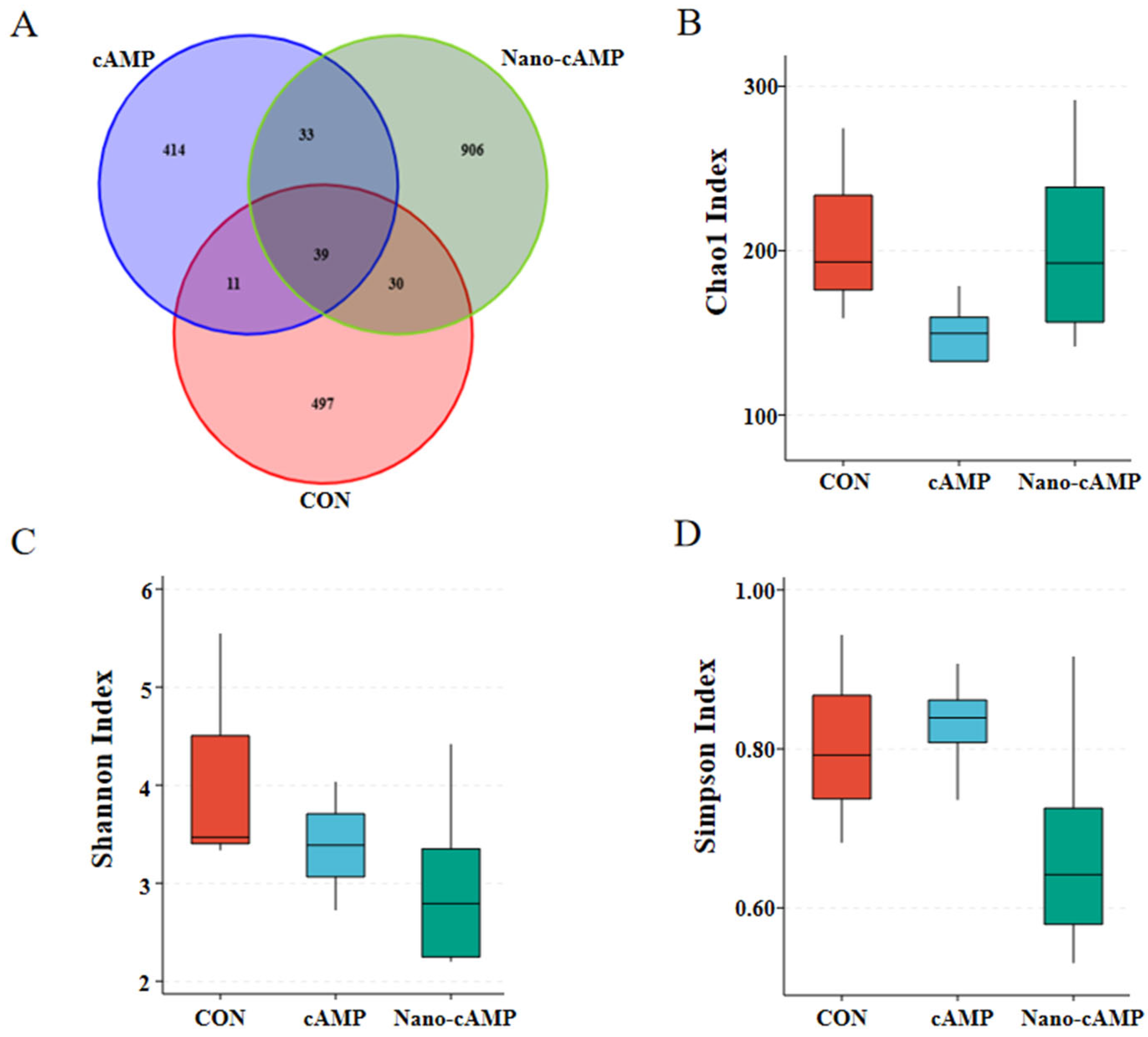
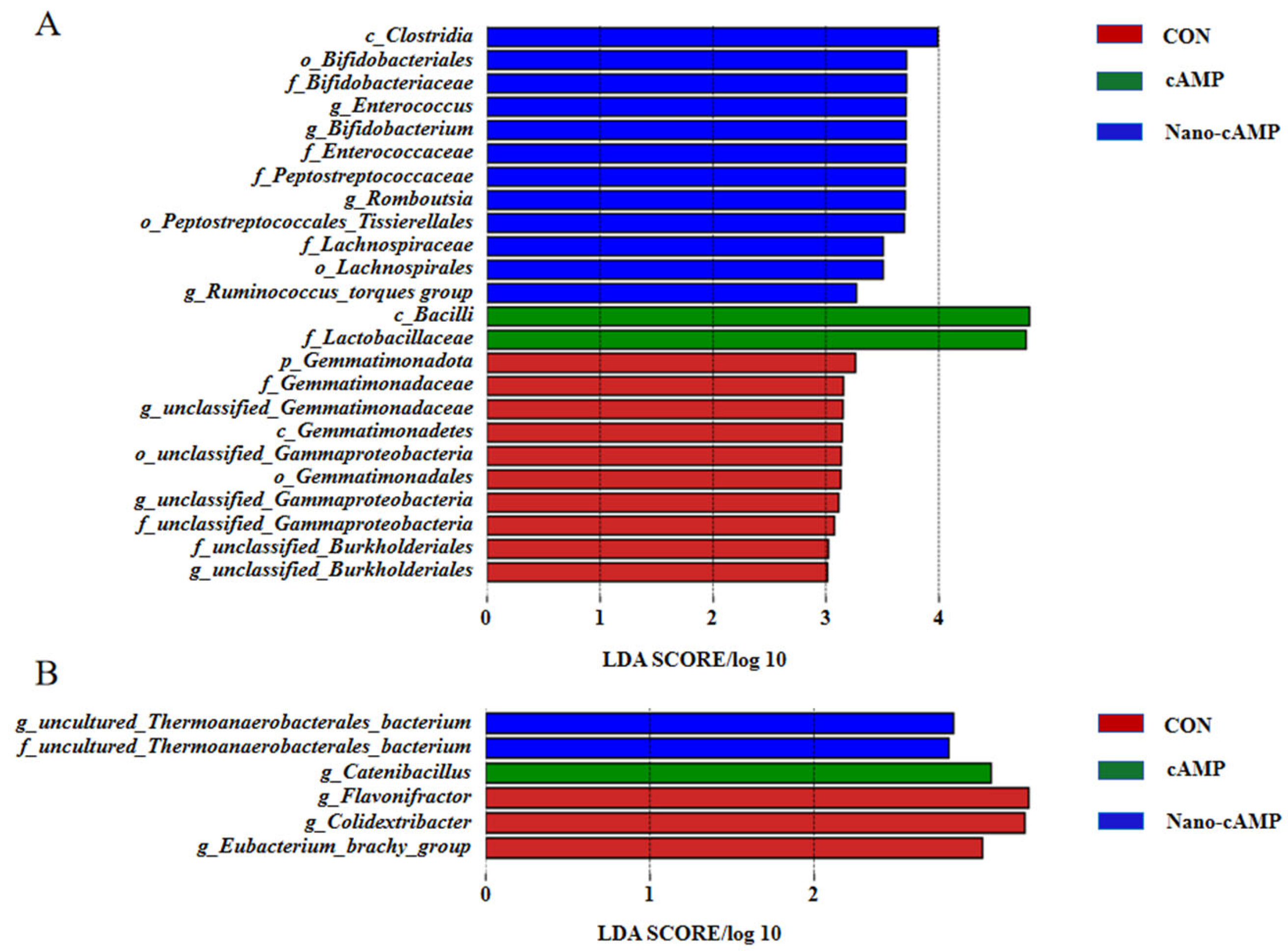
3.5. Effects on the Jejunal and Cecal Microbiota
4. Discussion
4.1. Effects of Nano-cAMP Liposomes on Growth Performance of Broilers
4.2. Effects of Nano-cAMP Liposomes on Major Digestive Organ Indices and Jejunal Tissue Morphology
4.3. Effects of Nano-cAMP Liposomes on Duodenal Enteric Hormone mRNA Expression
4.4. Effects of Nano-cAMP Liposomes on Intestinal Microbiota of Broilers
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qi, L.; Friml, J. Tale of cAMP as a second messenger in auxin signaling and beyond. New Phytol. 2023, 240, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sun, D.; Zhu, J.; Liu, J.; Zhu, W. The Regulation of Bacterial Biofilm Formation by cAMP-CRP: A Mini-Review. Front. Microbiol. 2020, 11, 802. [Google Scholar] [CrossRef]
- Li, Z.; Tan, H.; Lu, N.; Zhang, Z.; Chang, J. Enhanced ATP and antioxidant levels for cAMP biosynthesis by Arthrobacter sp. CCTCC 2013431 with polyphosphate addition. Biotechnol. Lett. 2021, 43, 2223–2231. [Google Scholar] [CrossRef]
- Allan, Z. Control of food intake through regulation of cAMP. Curr. Top. Dev. Biol. 2005, 67, 207–224. [Google Scholar] [CrossRef]
- Wang, S.; Chen, A. The physiological functions of cyclic adenosine monophosphate and its application in animal production. China Feed 2003, 10, 3–5. [Google Scholar] [CrossRef]
- Chen, W.; Xia, W.G.; Ruan, D.; Wang, S.; Abouelezz, K.F.M.; Wang, S.L.; Zhang, Y.N.; Zheng, C.T. Dietary calcium deficiency suppresses follicle selection in laying ducks through mechanism involving cyclic adenosine monophosphate-mediated signaling pathway. Animal 2020, 14, 2100–2108. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Li, W.; Xiao, Y.N.; Yang, X.; Gao, K.; Jiang, Z. Dietary dibutyryl cAMP supplementation regulates the fat deposition in adipose tissues of finishing pigs via cAMP/PKA pathway. Anim. Biotechnol. 2023, 34, 921–934. [Google Scholar] [CrossRef]
- Zahednezhad, F.; Saadat, M.; Valizadeh, H.; Parvin, Z.; Behzad, B. Liposome and immune system interplay: Challenges and potentials. J. Control. Release 2019, 305, 194–209. [Google Scholar] [CrossRef]
- Shahida, A.S.; Nur, A.B.; Ahmed, T.; Abdel-Moneim, E.A.M.; Abdelrazeq, M.S.; Chen, T.; Mohammad, S.K.; Yuan, L.; Elham, A.; Roberto, C.M.; et al. Bioactive-loaded nanodelivery systems for the feed and drugs of livestock; purposes, techniques and applications. Adv. Colloid Interface Sci. 2022, 308, 102772. [Google Scholar] [CrossRef]
- Gomes, I.; Sharma, S.K. Uptake of Liposomally Entrapped Adenosine-3′-5′-Cyclic Monophosphate in Mouse Brain. Neurochem. Res. 2004, 29, 441–446. [Google Scholar] [CrossRef]
- Fu, M.Q.; Wang, Y.S. Cyclic Adenosine Monophosphate Liposome Injection and Preparation Method. CN102327224A, 25 January 2012. [Google Scholar]
- NY/T 33-2004; Ministry of Agriculture of the People’s Republic of China. Feeding Standard of Chicken. China Agriculture Press: Beijing, China, 2004.
- Lin, J.; Comi, M.; Vera, P.; Alessandro, A.; Qiu, K.; Wang, J.; Wu, S.; Qi, G.; Zhang, H. Effects of Saccharomyces cerevisiae hydrolysate on growth performance, immunity function, and intestinal health in broilers. Poult. Sci. 2023, 102, 102237. [Google Scholar] [CrossRef]
- Sreerag, G.; Preetha, B. Evaluation and Clinical Comparison Studies on Liposomal and Non-Liposomal Ascorbic Acid (Vitamin C) and their Enhanced Bioavailability. J. Liposome Res. 2020, 31, 356–364. [Google Scholar] [CrossRef]
- Ahmed, K.; Hussein, S.; Ali, A.; Sameh, A.; Qiu, L.; Chen, J. Liposome: Composition, characterisation, preparation, and recent innovation in clinical applications. J. Drug Target. 2019, 27, 742–761. [Google Scholar] [CrossRef]
- Gancedo, J.M. Biological roles of cAMP: Variations on a theme in the different kingdoms of life. Biol. Rev. 2013, 88, 645–668. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Chang, L.; Zhang, G.; Song, Z.H.; Wan, D.; Xie, C.Y.; Wang, H.; Fan, Z.Y. Oral administration of dibutyryl adenosine cyclophosphate improved growth performance in weaning piglets by enhancing lipid fatty acids metabolism. Anim. Nutr. 2018, 4, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, G.E.; Metzler-Zebeli, B.U.; Lawlor, P.G. Impact of Intestinal Microbiota on Growth and Feed Efficiency in Pigs: A Review. Microorganisms 2020, 8, 1886. [Google Scholar] [CrossRef]
- Banerjee, P.; Carmelo, V.A.O.; Kadarmideen, H.N. Genome-wide epistatic interaction networks affecting feed efficiency in duroc and landrace pigs. Front. Genet. 2020, 11, 121. [Google Scholar] [CrossRef]
- Bedford, A.; Yu, H.; Hernandez, M.; Squires, E.J.; Leeson, S.; Gong, J. Effects of fatty acid glyceride product SILOhealth 104 on the growth performance and carcass composition of broiler chickens. Poult. Sci. 2018, 97, 1315–1323. [Google Scholar] [CrossRef]
- Elsharkawy, M.; Chen, Y.; Liu, R.; Xiaodong, T.; Wei, L.; Ibrahim, E.; Dongqin, Z.; Maiqing, Z.; Jie, W.; Guiping, Z. Paternal dietary methionine supplementation improves carcass traits and meat quality of chicken progeny. Animals 2021, 11, 325. [Google Scholar] [CrossRef]
- Yue, Y.X.; Xiao, L.Q.; Ming, Y.H.; Ruiyi, L.; Ye, H.; Zhangxu, W.; Huanhuan, Z.; Yunxia, Z.; Yu, L.; Shuhong, Z.; et al. Transcriptome Analysis of Adipose Tissue Indicates That the cAMP Signaling Pathway Affects the Feed Efficiency of Pigs. Genes 2018, 9, 336. [Google Scholar] [CrossRef]
- Lu, J.; Ye, H.; Hui, W.; Yuanxin, M.; Xinyun, L.; Jianhua, C.; John, M.B.; Tim, P.; Shuhong, Z. Transcriptome analysis of mRNA and miRNA in skeletal muscle indicates an important network for differential Residual Feed Intake in pigs. Sci. Rep. 2015, 5, 11953. [Google Scholar] [CrossRef]
- Chuang, W.Y.; Hsieh, Y.C.; Lee, T.T. The Effects of Fungal Feed Additives in Animals: A Review. Animals 2020, 10, 805. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Wu, S.G.; Zhang, H.J.; Guanghai, Q.; Jing, W. Dynamic alterations in early intestinal development, microbiota and metabolome induced by in ovo feeding of L-arginine in a layer chick model. J. Anim. Sci. Biotechnol. 2020, 011, 896–911. [Google Scholar] [CrossRef]
- Wang, S.; Chen, Y.; Guo, J.; Huang, Q. Liposomes for Tumor Targeted Therapy: A Review. Int. J. Mol. Sci. 2023, 24, 2643. [Google Scholar] [CrossRef]
- Adewole, D.I.; Oladokun, S.; Santin, E. Effect of organic acids-essential oils blend and oat fiber combination on broiler chicken growth performance, blood parameters, and intestinal health. Anim. Nutr. 2021, 4, 1039–1051. [Google Scholar] [CrossRef]
- Monika, B.T.; Ewa, Z.; Anna, S.; Maria, S.; Joanna, B. Modulation of Intestinal Histology by Probiotics, Prebiotics and Synbiotics Delivered In Ovo in Distinct Chicken Genotypes. Animals 2021, 11, 3293. [Google Scholar] [CrossRef] [PubMed]
- Guerra, D.G.; Bok, R.; Ramon, A.L.; Joseph, H.K. Protein kinase A facilitates relaxation of mouse ileum via phosphorylation of neuronal nitric oxide synthase. Br. J. Pharmacol. 2020, 177, 2765–2778. [Google Scholar] [CrossRef]
- Ahmed, K.; Takaoki, S.; Hiroshi, K.; Kazuhisa, H. Effects of fasting and re-feeding on the expression of CCK, PYY, hypothalamic neuropeptides, and IGF-related genes in layer and broiler chicks. Comp. Biochem. Physiol. 2021, 257, 110940. [Google Scholar] [CrossRef]
- Anastasiia, R.V.; Ilya, R.A.; Mikhail, A.K.; Ivan, S.Y.; Michael, N.R.; Elena, I.S.; Oleg, A.G.; Fedor, A.K. A bird’s-eye overview of molecular mechanisms regulating feed intake in chickens-with mammalian comparisons. Anim. Nutr. 2024, 17, 61–74. [Google Scholar] [CrossRef]
- Bany, B.R.; Reimann, F.; Gribble, F.M. The intestine as an endocrine organ and the role of gut hormones in metabolic regulation. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 784–796. [Google Scholar] [CrossRef]
- Zhu, S.G.; Xu, C.F. Biochemistry, 4th ed.; Higher Education Press: Beijing, China, 2017; pp. 1–596. [Google Scholar]
- Wu, S.; Shen, Y.; Zhang, S.; Yunqi, X.; Shourong, S. Salmonella Interacts with Autophagy to Offense or Defense. Front. Microbiol. 2020, 11, 721. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Lee, J.; Tan, K. Infection with pathogenic Blastocystis ST7 is associated with decreased bacterial diversity and altered gut microbiome profiles in diarrheal patients. Parasites Vectors 2022, 15, 312. [Google Scholar] [CrossRef]
- Cao, H.; Chen, D.; Guo, L.; Rong, J.; Yunteng, X.; Wei, M.; Chen, W.; Pengfei, L.; Hui, W. Effects of Bacillus subtilis on growth performance and intestinal flora of Penaeus vannamei. Aquac. Rep. 2022, 23, 101070. [Google Scholar] [CrossRef]
- Zhang, X.; Akhtar, M.; Chen, Y.; Ziyu, M.; Yuyun, L.; Deshi, S.; Ranran, C.; Lei, C.; Yafang, H.; Abdallah, A.N.; et al. Chicken jejunal microbiota improves growth performance by mitigating intestinal inflammation. Microbiome 2022, 10, 107. [Google Scholar] [CrossRef]
- Qiu, K.; Li, C.L.; Wang, J. Effects of Dietary Supplementation with Bacillus subtilis, as an Alternative to Antibiotics, on Growth Performance, Serum Immunity, and Intestinal Health in Broiler Chickens. Front. Nutr. 2021, 8, 786878. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Z.; Li, F.; Hu, L.; Xiao, T.; Zhao, Y.; Yang, M. The Effects of Dietary Fermented Soybean Residue on the Growth, Antioxidant Capacity, Digestive Enzyme Activities, and Microbial Compositions of the Intestine in Furong Crucian Carp (Furong Carp♀×Red Crucian Carp♂). Fishes 2024, 9, 138. [Google Scholar] [CrossRef]
- Ivan, R. Composition and Function of Chicken Gut Microbiota. Animals 2020, 10, 103. [Google Scholar] [CrossRef]
- Yu, X.; Niu, S.; Tie, K.; Zhang, Q.; Deng, H.; Gao, C.; Yu, T.; Lei, L.; Feng, X. Characteristics of the intestinal flora of specific pathogen free chickens with age. Microb. Pathog. 2019, 132, 325–334. [Google Scholar] [CrossRef]
- Paneru, D.; Tellez, I.G.; Bottje, W.G.; Emmanuel, A.; Ahmed, A.A.A.W.; Md, S.; Jayant, L. Modulation of Immune Response and Cecal Microbiota by Dietary Fenugreek Seeds in Broilers. Vet. Sci. 2024, 11, 57. [Google Scholar] [CrossRef]
- Francesca, D.F.; Edoardo, P.; Danilo, E. Newly Explored Faecalibacterium Diversity Is Connected to Age, Lifestyle, Geography and Disease. Curr. Biol. 2020, 30, 4932–4943. [Google Scholar] [CrossRef]
- Shinuo, L.; Qingfeng, W.; Jinqiu, M.; Haotian, C.; Tianhao, Y.; Yue, W.; Lihong, Z.; Qiugang, M.; Shimeng, H. Lactobacillus crispatus-Mediated Gut-Reproductive Tract Axis-Alleviated Microbial Dysbiosis and Oviductal Inflammation in a Laying Hen Model. Microorganisms 2024, 12, 1559. [Google Scholar] [CrossRef]
- Przemysław, L.; Sebastian, B. Effect of hyperthermophilic pretreatment on methane and hydrogen production from garden waste under mesophilic and thermophilic CONditions. Bioresour. Technol. 2021, 335, 125264. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, Y.; Xu, Z.; Yuming, Z.; Lei, W.; Chunguang, L.; Ailing, C.; Shanyu, J.; Weimin, S.; Guohui, Y.; et al. Dysregulation of gut health in zebrafish by differentially charged nanoplastic exposure: An integrated analysis of histopathology, immunology, and microbial informatics. Environ. Sci. Nano 2023, 10, 933–947. [Google Scholar] [CrossRef]
- Chaudhary, D.K.; Kim, S.E.; Park, H.J.; Kyoung, H.K. Unveiling the Bacterial Community across the Stomach, Hepatopancreas, Anterior Intestine, and Posterior Intestine of Pacific Whiteleg Shrimp. J. Microbiol. Biotechnol. 2024, 34, 1260–1269. [Google Scholar] [CrossRef]
- Wang, S.; Yin, F.; Guo, Z.; Rui, L.; Wei, S.; Yuchao, W.; Yichen, G.; Chao, S.; Daqing, S. Association between gut microbiota and glioblastoma: A Mendelian randomization study. Front. Genet. 2024, 14, 1308263. [Google Scholar] [CrossRef]
- Ren, X.; Xu, J.; Xu, Y.; Qin, W.; Kunlun, H.; Xiaoyun, H. Artemether Attenuates Gut Barrier Dysfunction and Intestinal Flora Imbalance in High-Fat and High-Fructose Diet-Fed Mice. Nutrients 2023, 15, 4860. [Google Scholar] [CrossRef]
- Hu, X.; Ouyang, S.; Xie, Y.; Gong, Z.; Du, J. Characterizing the gut microbiota in patients with chronic kidney disease. Postgrad. Med. 2020, 132, 495–505. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Content |
|---|---|
| cAMP | 5.41 |
| Lecithin | 22.70 |
| Cholesterol | 4.32 |
| Citric acid monohydrate(C6H8O7·H2O) | 22.16 |
| Sodium citrate dihydrate (Na3C6H5O7·2H2O) | 7.57 |
| Chitosan(MW 200,000) | 10.81 |
| Lactose | 27.03 |
| Total | 100.00 |
| Items | Content/% |
|---|---|
| Ingredients | |
| Corn | 37.50 |
| Wheat | 25.00 |
| Soybean meal | 2.50 |
| Wheat bran | 15.00 |
| Corn protein meal | 12.50 |
| Soybean oil | 2.00 |
| Limestone | 1.25 |
| CaHPO4 | 2.15 |
| L-Lysine·H2SO4 | 0.84 |
| DL-Methionine | 0.11 |
| NaCl | 0.25 |
| Choline chloride | 0.10 |
| NaHCO3 | 0.10 |
| Phytase | 0.02 |
| Cellulase | 0.05 |
| Premix 1 | 0.63 |
| Total | 100.00 |
| Nutrient levels | |
| Metabolizable energy/(MJ/kg) | 12.12 |
| Crude protein | 18.70 |
| Lysine | 0.91 |
| Methionine | 0.45 |
| Methionine + Cysteine | 0.70 |
| Calcium | 1.10 |
| Total phosphorus | 0.72 |
| Gene Name 1 | Primer Sequence (5′-3′) | Product Size (bp) |
|---|---|---|
| CCK | F: ACTGGGAGGTTCTCTGTCCT | 155 |
| R: CTGTTGCTATCGCCTGCTGT | ||
| Ghrelin | F: AACAGCAAGATTACATCGCCG | 169 |
| R: CATCTTCTCCAGTGCTTGTCC | ||
| Secretin | F: AGCAATCCAAGCCTGGTCA | 132 |
| R: AGCTTCCTTGGCATCGTTGT | ||
| GIP | F: ATGCACAGACGCTACTCGG | 153 |
| R: AGGCTCGGCTTCTCTCTTGT | ||
| GAPDH | F: ATGGGCACGCCATCACTATC | 138 |
| R: CACCACCCTTCAGATGAGCC |
| Items 1 | CON | cAMP | Nano-cAMP | SEM | p-Value |
|---|---|---|---|---|---|
| Initial body weight/g | 354.00 | 353.75 | 352.92 | 3.042 | 0.989 |
| Final body weight/g | 1096.00 | 1083.67 | 1128.75 | 10.390 | 0.138 |
| Feed/gain ratio | 2.39 a | 2.33 ab | 2.27 b | 0.019 | 0.025 |
| Average daily feed intake/g | 84.43 | 80.08 | 83.93 | 1.129 | 0.253 |
| Average daily gain/g | 35.33 | 34.42 | 36.94 | 0.494 | 0.122 |
| Items 1 | CON | cAMP | Nano-cAMP | SEM | p-Value |
|---|---|---|---|---|---|
| Glandular gastric index | 0.40 | 0.41 | 0.43 | 0.022 | 0.907 |
| Muscle stomach index | 1.33 | 1.44 | 1.34 | 0.032 | 0.301 |
| Duodenal index | 0.46 b | 0.61 a | 0.49 b | 0.026 | 0.011 |
| Jejunal index | 0.91 | 1.10 | 0.99 | 0.044 | 0.264 |
| Ileal index | 0.76 | 0.70 | 0.70 | 0.026 | 0.631 |
| Cecal index | 0.34 | 0.33 | 0.31 | 0.013 | 0.741 |
| Items 1 | CON | cAMP | Nano-cAMP | SEM | p-Value |
|---|---|---|---|---|---|
| Villous height/μm | 1084.57 b | 1330.44 ab | 1423.81 a | 56.947 | 0.033 |
| Crypt depth/μm | 155.59 | 181.06 | 183.75 | 10.752 | 0.570 |
| Villus height/Crypt depth | 7.11 | 7.52 | 7.91 | 0.331 | 0.660 |
| Items 1 | CON | cAMP | Nano-cAMP | SEM | p-Value |
|---|---|---|---|---|---|
| CCK | 1.00 b | 5.24 a | 4.81 a | 0.695 | 0.003 |
| Ghrelin | 1.01 | 1.03 | 1.31 | 0.078 | 0.209 |
| Secretin | 1.02 b | 2.65 a | 2.31 a | 0.320 | 0.006 |
| GIP | 1.01 | 0.90 | 1.10 | 0.053 | 0.400 |
| Items 1 | CON | cAMP | Nano-cAMP | SEM | p-Value |
|---|---|---|---|---|---|
| Firmicutes | 83.640 b | 97.526 a | 95.152 a | 2.110 | 0.022 |
| Proteobacteria | 5.350 a | 1.013 b | 1.434 b | 0.629 | 0.007 |
| Bacteroidota | 2.729 | 0.310 | 0.468 | 0.435 | 0.063 |
| Actinobacteriota | 1.320 | 0.535 | 2.138 | 0.321 | 0.085 |
| Cyanobacteria | 0.810 | 0.022 | 0.052 | 0.154 | 0.097 |
| Acidobacteriota | 1.133 | 0.097 | 0.126 | 0.188 | 0.058 |
| Chloroflexi | 0.483 a | 0.068 b | 0.084 b | 0.069 | 0.029 |
| Myxococcota | 0.779 | 0.037 | 0.040 | 0.160 | 0.150 |
| Gemmatimonadota | 0.354 a | 0.010 b | 0.055 b | 0.050 | 0.010 |
| Others | 3.403 a | 0.382 b | 0.450 b | 0.474 | 0.012 |
| Items 1 | CON | cAMP | Nano-cAMP | SEM | p-Value |
|---|---|---|---|---|---|
| Firmicutes | 70.692 | 68.167 | 65.144 | 2.133 | 0.597 |
| Bacteroidota | 18.877 | 18.953 | 20.350 | 1.806 | 0.940 |
| Actinobacteriota | 9.362 | 11.655 | 13.544 | 1.406 | 0.506 |
| Desulfobacterota | 0.535 | 0.540 | 0.666 | 0.104 | 0.861 |
| Proteobacteria | 0.296 | 0.154 | 0.161 | 0.054 | 0.511 |
| Campylobacterota | 0.078 | 0.414 | 0.045 | 0.087 | 0.163 |
| Cyanobacteria | 0.058 | 0.064 | 0.071 | 0.029 | 0.985 |
| Verrucomicrobiota | 0.009 | 0.011 | 0.009 | 0.003 | 0.941 |
| Acidobacteriota | 0.016 | 0.005 | 0.002 | 0.003 | 0.078 |
| Others | 0.078 | 0.036 | 0.008 | 0.014 | 0.131 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Wang, L.; Huang, S.; Su, G.; Jiang, S.; Yang, F.; Huang, J. Effects of Cyclic Adenosine Monophosphate Nanoliposomes on Growth Performance, Gut Development and Microbiota of Broilers. Animals 2025, 15, 1852. https://doi.org/10.3390/ani15131852
Chen L, Wang L, Huang S, Su G, Jiang S, Yang F, Huang J. Effects of Cyclic Adenosine Monophosphate Nanoliposomes on Growth Performance, Gut Development and Microbiota of Broilers. Animals. 2025; 15(13):1852. https://doi.org/10.3390/ani15131852
Chicago/Turabian StyleChen, Li, Li Wang, Sheng Huang, Guoqi Su, Shan Jiang, Feiyun Yang, and Jingxiu Huang. 2025. "Effects of Cyclic Adenosine Monophosphate Nanoliposomes on Growth Performance, Gut Development and Microbiota of Broilers" Animals 15, no. 13: 1852. https://doi.org/10.3390/ani15131852
APA StyleChen, L., Wang, L., Huang, S., Su, G., Jiang, S., Yang, F., & Huang, J. (2025). Effects of Cyclic Adenosine Monophosphate Nanoliposomes on Growth Performance, Gut Development and Microbiota of Broilers. Animals, 15(13), 1852. https://doi.org/10.3390/ani15131852